
Abstract
We investigated the persistence of SARS-CoV-2-specific neutralizing antibodies in serum (CoV-2-SNAb) against the “WH-Human 1” coronavirus in 57 convalescent persons from January 2020 to January 2021. The CoV-2-SNAb response against authentic “WH-Human 1” showed a significant (p < 0.01) neutralizing high effect (≥95%) in the following manner: by 94.7% neutralization for up to 6 months, by 73.1% for up to 8 months, and by 31.7% for up to 10 months in correlation with a significant decrease in the concentration of the virus determined by SARS-CoV-2 spike protein extracellular domain and spike-receptor-binding domain (S-RBD). There was neutralizing effect (<95%) when the S-RBD optical density (OD) value was more than 1.0, showing a suitable threshold of S-RBD = 1.0 (antibody-tittering, OD). However, in some convalescent persons, no neutralizing effect (<95%) was observed although the SARS-CoV-2-specific neutralizing antibodies were bound to the S-RBD (OD >1.0). The neutralization of the virus in these cases may not involve S-RBD, but rather B- and T cell memory responses in overall immunity, using the threshold value (OD = 1.0) of S-RBD as a simple and effective method to determine the neutralization effect of the antibody efficacy and use of vaccination in combination with a standard pseudovirus neutralizing assay. We suggest that convalescent persons should contact their physicians 6-month postinfection to test the function of their serum neutralizing antibodies and determine whether administering a SARS-CoV-2 vaccine is necessary to prevent the development of severe illness in the future.
Introduction
COVID-19
Neutralizing antibodies usually target the spike-receptor-binding domain (S-RBD). The crystal structure of neutralizing antibodies isolated from SARS-CoV-2 convalescent persons demonstrated that the S-RBD bound to the cell receptor angiotensin-converting enzyme 2 (Bošnjak et al, 2021; Yan et al, 2020; Yuan et al, 2020). Today, the gold standard for confirmation of infection is real-time reverse transcription-polymerase chain reaction (RT-PCR) detection of viral RNA in oral swabs (Chen et al, 2020). Although RT-PCR is used to confirm viral infections, viral immunoglobulin M (IgM) and immunoglobulin G (IgG) serological tests are useful methodologies for immunological screening. The combination of RT-PCR (Tang et al, 2020) and IgG/IgM assays improves the robustness of laboratory diagnostics (Kellam et al, 2000; Hiki et al, 2021; Valdez-Cruz et al, 2021; Veyrenche et al, 2021; Wajnberg et al, 2020). Serum levels of SARS-CoV-2-specific IgG and IgM (n = 285, Wanzhou, Chongqing, China, 2020) were 100% positive within 19 days after symptom onset (Long et al, 2020).
IgG antibodies (n = 136, between April and June 2020, Traunstein, Germany) were present for approximately one year, while IgM antibodies were short and detectable only briefly (Glück et al, 2022). IgG levels (n = 50, March–May 2020, Euroimmun, Lübeck, Germany) strongly correlated with symptom severity in ∼90% of mildly and 100% of severely infected persons (Bewley et al, 2021). A robust IgG response against the viral spike protein at ∼5 months postinfection (n = 30,082, April–October 2020, Mount Sinai, NY) was significantly correlated with neutralization of authentic SARS-CoV-2 (Wajnberg et al, 2020). The half-life of the anti-SARS-CoV-2-directed IgG antibody (n = 353, April–October 2020, Bilthoven, The Netherlands) was 158 days (den Hartog et al, 2021) and 85 days (n = 452, March–April 2020, Oxfordshire, UK) (Lumley et al, 2021).
An authentic neutralizing assay against hCoV-19/Austria/CeMM1012/2020 (n = 34, between April and June 2020, Wuerzburg, Germany) showed 74% neutralizing antibodies for up to 10 months (Sonnleitner et al, 2022). In another study, a pseudotype neutralization assay (n = 148, April 28, 2020, Athens, Greece) demonstrated a median of 8.3 months after first symptom onset (Terpos et al, 2021). An authentic neutralizing activity against the “D614G” mutation significantly decreased 6–8 months postinfection (n = 23, between April 2021 and May 2021, Kakogawa, Hyogo, Japan) (Kurahashi et al, 2022).
In this study, we investigated how long time the SARS-CoV-2 serum-neutralizing antibodies (CoV-2-SNAb) were maintained in the serum and how efficiently these CoV-2-SNAb neutralized the SARS-CoV-2 virus.
Materials and Methods
Study design
The inclusion of the study was for all convalescent and noninfected persons who met the following criteria: (1) individuals who demonstrated detection of nasopharyngeal swab samples by fluorescence RT‒PCR; (2) individuals with CoV-2-SNAbs confirmed by an authentic virus neutralization assay; (3) individuals who demonstrated CoV-2-SNAb binding to the SARS-CoV-2 spike protein extracellular domain (S-ECD), spike-receptor-binding domain (S-RBD), and N-terminal domain (S-NTD); and (4) Thoracic computed tomography. Exclusion criteria are all convalescent patients who tested positive for COVID-19 and noninfected persons who had received a COVID-19 vaccination during the time of the study.
Blood donors
In this study, a total of 57 volunteer blood donors who were SARS-CoV-2 infected (from December 2019 to January 2020) and then convalescent (January 2020 to January 2021) participated. Serum samples were collected from Xiangtan Central Hospital (n = 3, February 2020 to January 2021) and Renmin Hospital of Wuhan University (n = 54, May 2020 to January 2021). The serum samples were collected for 12 months. Some of the 146 control participants (n = 14) were tested with an authentic SARS-CoV-2 virus assay in a biosafety level 3 (BSL-3) laboratory. Ten of the 14 persons in the control group were closely related to SARS-CoV-2-infected persons. Three persons worked with the medical staff, and seven persons were family members. Four of the 14 persons had no contact with any infected people.
Classification of SARS-CoV-2-infected persons
The classification was performed according to the National Health Commission of the People's Republic of China. Diagnosis and Treatment Protocol for COVID-19 (Trial Version 7), as the classification of the infected, convalescent, and noninfected persons, are shown in detail (National Health Commission of the People's Republic of China, 2020). The information on the infected, convalescent, and noninfected persons is summarized in Table 1.
Characterization of Infected, Convalescents, and Noninfected Persons
The nasopharyngeal swab sample: RT-PCR detection Kit was provided from Data Gene Antigen Rapid Test Kit, Wuhan Easy Diagnosis Biomedicine Co. Ltd., China. The IgM/IgG kits were provided by Nanjing Vazyme Medical Technology Co., Ltd. or Shenzhen Yhlo Biotech Co. Ltd., China.
Fourteen: detection limit of 14/146 (9.6%) for the authentic SARS-CoV-2 virus assay. Classification of the SARS-CoV-2-infected persons: mild, moderate, serious, serious-critical, ND.
CT, computed tomography; IgG, immunoglobulin G; IgM, immunoglobulin M; ND, no data; RT-PCR, real-time reverse transcription-polymerase chain reaction.
Collection of blood samples
Blood samples of 5 mL were collected between 7 and 9 a.m. from blood donors individually who had not eaten any breakfast. The drawn venous blood was stored for 2–3 h at room temperature (RT) in nonheparin tubes, centrifuged at 4,000 rpm for 8–10 min, the residual virus was inactivated at 56°C for 1 h (Tseng et al, 2012) and then frozen at −80°C until analysis at approximately one month.
Enzyme-linked immunosorbent assay titer of CoV-2-SNAb
When this project started in early January 2020, there were no available kits for S-ECD, S-RBD, or S-NTD in China to verify the specific CoV-2-SNAb. The antigens (S-ECD, S-RBD, and S-NTD) were purchased from Nanjing Jinsirui Biotechnology Co., Ltd., China). The binding of Cov-2-NpAb to S-ECD, S-RBD, or S-NTD was determined by enzyme-linked immunosorbent assay in our laboratory.
According to the gradient dilution method, we repeated the assays using different gradient dilutions of serum samples from convalescent- and noninfected parsons. We confirmed that the optimal dilution was 500 × for serum samples at a fixed excessive concentration of S-ECD, S-RBD, or S-NTD of 0.5 μg/mL. In brief, 96-well plates (Thermo Fisher Scientific, USA) were coated with 0.5 μg/mL S-ECD, S-RBD, S-NTD, or phosphate-buffered saline (PBS) (control) overnight at 4°C. The wells were rinsed once and blocking buffer (5% nonfat dried milk in PBS) was added for 1 h at 37°C and washed once thereafter.
The wells were rinsed three times and incubated for 1 h at 37°C with 100 μL per well of 1:10,000 HRP-conjugated goat-IgG followed by three washes in PBS buffer. Tetramethylbenzidine (100 μL per well) was added to the wells and incubated at 37°C for 10 min. Then, 2 M H2SO4 was added (100 μL per well) to terminate the reaction. The plates were then scanned in a spectrophotometer at an absorbance of 450 nm (chromogenic substrate). The data were replicated in two wells for all samples, and the curriculum vitae (CV) of the mean value was <5%.
CoV-2-SNAb response against authentic SARS-CoV-2 virus by RT-PCR
Reliable SARS-CoV-2 RT-PCR assays were based on the genome sequence of SARS-CoV-2 in blood donors (Li et al, 2020). This study was tested in laminar-flow-safety cabinets at BSL-3 at Disease Control, Shenzhen, China. In brief, 1 × 105 Vero E6 (TMPRSS2) cells per well were seeded in 96-well tissue culture plates in 100 μL Dulbecco's Modified Eagle Medium (DMEM) per well (10% fetal bovine serum [FBS] in DMEM with penicillin/streptomycin) and incubated overnight at 37°C in a CO2 incubator. According to 100 median tissue culture infectious dose (TCID)50, 90 μL of heat-inactivated sera and 10 μL of “WH-Human 1” coronavirus (Veyrenche et al, 2021) were mixed in DMEM and incubated at 37°C for 1 h and then added to the seeded Vero E6 cells per well in 96-well tissue culture plates in maintenance DMEM (2% FBS in DMEM with penicillin/streptomycin) and incubated at 37°C in a 5% CO2 incubator for an additional 48 h.
PBS was used as a control and ran in parallel. The total RNA of the virus from the DMEM was extracted according to the method described by the manufacturer (Roche's High Pure Viral RNA Purification Kit, USA; No. 11859992001). The copy number of SARS-CoV-2 RNA in the total RNA was detected with the fluorescence quantitative RT-PCR method (Shanghai Berger's Coronavirus Nucleic Acid Detection Kit, Shanghai, China). The nucleic acid concentration in the virus sample was calculated based on a formula, including the Ct value (number of PCR cycles giving detectable values) of the open reading frame 1 ab (ORF1ab) and the nucleocapsid protein (N-gene). The primers and probes for the SARS-CoV-2 virus were used to target both the ORF1ab and N genes. The values below the threshold of the ID50 calculation were entered as 0.5. The values below the assay's detection limit were considered negative and entered as 0.1.
All data were plotted after background subtraction and normalized to the controls. The data were repeated twice for all samples, and the CV of the mean value was <5%. The neutralization rate was accepted when the virus reached ≥95% (Li et al, 2020).
Statistical analysis
The statistical analysis was performed using the statistical program SPSS 25.0 (IBM Corp., New York, NY), including the mean ± standard deviation, Pearson correlation, Pearson's chi-square test, and two-tailed t-test. A box-whisker plot was used for descriptive data of median, maximum/min, and Q1/Q3. p-Value <0.05 was considered statistically significant. A Pearson correlation with an r-value >0.75 was considered a significant correlation.
Results
Time course analysis of the CoV-2-SNAb of convalescent persons
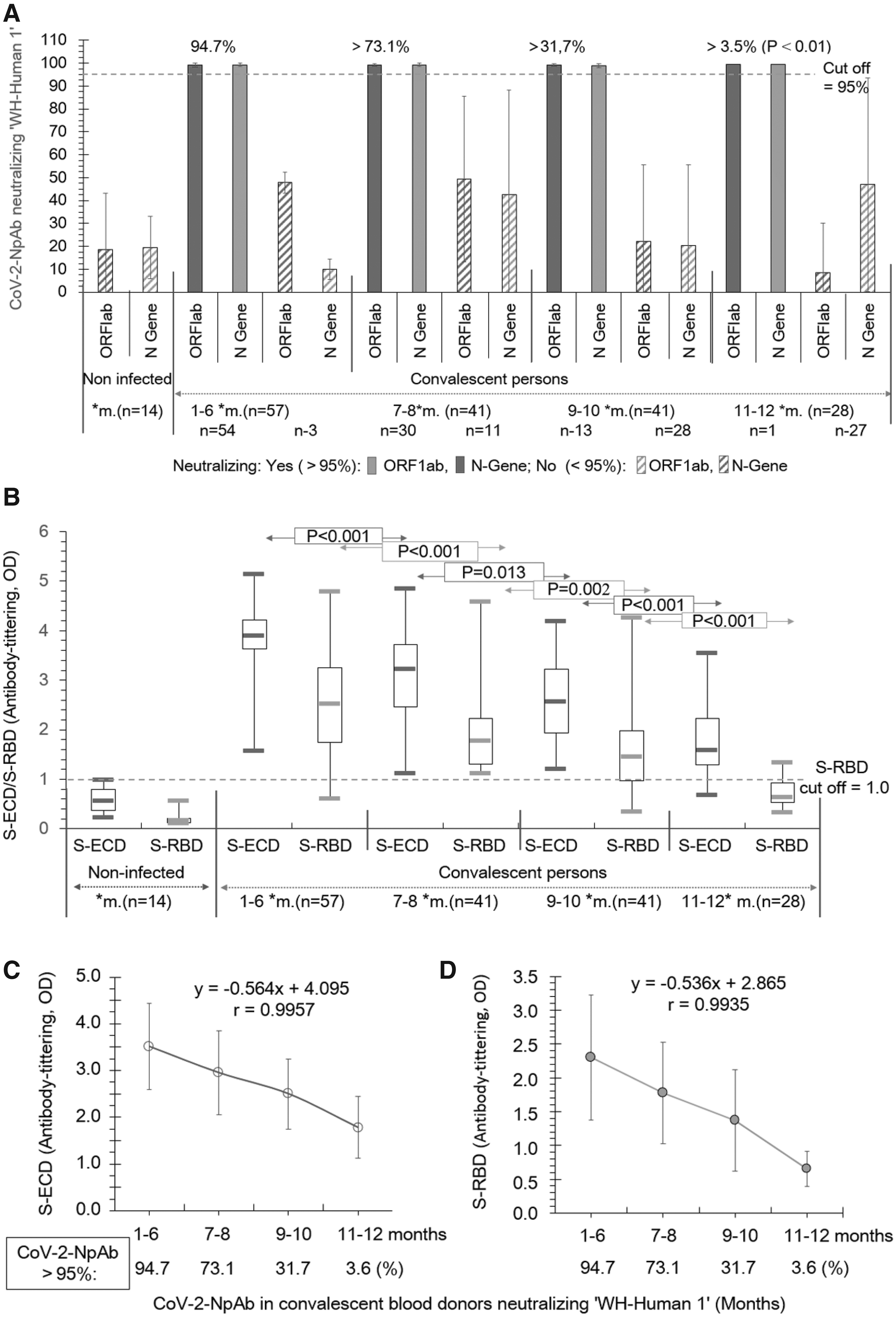
A total of 57 convalescent persons participate in this study. Fifty persons showed a mild/moderate degree of infection, while only seven persons showed a serious or serious-critical degree of infection. According to the inclusion and exclusion criteria in the study design, we summarized all data based on four-time groups at 1–6, 7–8, 9–10, and 11–12 months (Tables 1 and 2; Figs. 1 and 2).
Duration of SARS-CoV-2 against the WH-Human 1 virus in convalescent blood donors.

CoV-2-SNAb neutralizing the WH-Human 1 virus in correlation with the kinetic change in the CoV-2-NpAb binding S-RBD (antibody titration, OD) in 7 convalescent blood donors individually. *Only one convalescent person at 12 months showed 100% neutralization of the SARS-CoV-2 virus.
Number of Convalescent Persons with Neutralizing Score Using the Authentic Virus Assay for 12 Months
Pearson chi-square. p-Value <0.05 was considered as a statistically significant difference.
The CoV-2-SNAb against the authentic Wuhan human virus (“WH-Human 1”) was tested by reliable SARS-CoV-2-RT-PCR assays (Li et al, 2020) based on the Ct value of the ORF1ab and the N-gene, as well as the S-ECD and S-RBD by antibody-tittering.
The results in Figure 1A and Table 2 show that CoV-2-SNAb neutralized the “WH-Human 1” coronavirus (≥95%) significantly decline (p < 0.01) in the following manner: 94.7% for up to 6 months, 73.1% for up to 8 months, 31.7% for up to 10 months, and 3.6 for up to 12 months, in relationship to a significant decrease in S-ECD and S-RBD concentrations (p < 0.01, Fig. 1B). A suitable cutoff value of S-RBD (antibody-tittering, optical density [OD] = 1.0) is suggested. The percentage of convalescent persons with neutralizing effects (>95%) decreased significantly, corresponding to the concentrations of S-ECD and S-RBD (Fig. 2C, r = 0.984 and Fig. 2D, r = 0.982), respectively. Only one person with a serious degree still showed neutralized virus (>95%) at 12 months (Fig. 1A). No neutralizing effect was found in 14 noninfected persons (<95%) (Fig. 1A, p > 0.05).
The results in Figure 2 show an example of the CoV-2-NpAb neutralizing “WH-Human 1” virus (>95%) with the kinetic change of the CoV-2-NpAb binding S-RBD individually during the 10 months (n = 7). The CoV-2-SNAb binding to S-RBD was approximately sixfold higher than the cutoff value of S-RBD (antibody-tittering, OD = 1.0) compared with the noninfected persons (OD = 0.17).
Analysis of SARS-CoV-2-SNAb neutralization of SARS-CoV-2 by targeting S-RBD
The next set of experiments aimed to further elucidate the ability of CoV-2-SNAb to neutralize the “WH-Human 1” virus in correlation with the kinetic change of the CoV-2-NpAb binding S-RBD individually. A total of 52 convalescent persons during the 5–10 months of data collection were divided into two groups: CoV-2-SNAb effective neutralizing (≥95%) and ineffective neutralizing (>95%) the virus. The results showed that the CoV-2-SNAb bound to S-RBD 100% (antibody titter, OD >1.0) in Group 1, while the CoV-2-SNAb binding to S-RBD was still high (OD >1.0) in 66% of the people in Group 2, although it was ineffective at neutralizing the virus (Fig. 3).

Correlation between CoV-2-SNAb neutralizing SARS-CoV-2 virus and targeting S-RBD (antibody titration, OD) from convalescent blood donors (n = 52) over a period of 7–10 months. Group 1: Neutralizing antibodies (≥95%, n = 28); Group 2: Non-neutralizing antibodies (<95%, n = 24). t-Test was used.
Thus, some other mechanism(s) might be involved in the interaction with S-RBD binding rather than neutralization of CoV-2-SNAb, such as B- and T cells. We did not investigate B- and T cells, but did investigate another immune-related phagocytic blood cell in 44 cases after symptom onset and found both elevated and decreased immune-related phagocytic blood cells (Table 3).
Information of Phagocytic Blood Cells Half-1 Month After Symptom Onset
Cases had high CoV-2-SNAb against the authentic “WH-Human 1” except for cases 45a and 46 b (black letter: unnormal value of phagocytic blood cells).
CoV-2-SNAb, SARS-CoV-2-specific neutralizing antibodies in serum.
In this study, the RT-PCR test of nasopharyngeal swab samples was based on routine testing. All 57 convalescents and 146 noninfected persons were RT-PCR negative, and no side effects were observed.
Discussion
In this study, we investigated the persistence of CoV-2-SNAb in 57 convalescents and how efficiently CoV-2-SNAb protected against the “WH-Human 1” coronavirus. A comprehensive analysis of all results showed that the CoV-2-SNAb significantly neutralized the “WH-Human-1” coronavirus in the following manner: 94.7% for up to 6 months, 73.1% for up to 8 months, and 31.7% for up to 10 months (p < 0.001), in agreement with previous studies (Hyseni et al, 2020; National Health Commission of the People's Republic of China, 2020; Park et al, 2020). The concentration of CoV-2-SNAb correlated significantly with the decrease in the concentrations of S-ECD and S-RBD (p < 0.01). However, CoV-2-SNAb was almost unable to neutralize the virus at 12 months.
There was no neutralizing effect (<95%) when the S-RBD OD-value was <1.0, indicating that the threshold of S-RBD can be used to evaluate the effect of CoV-2-SNAb binding to S-RBD. In addition to neutralizing the “WH-Human 1” coronavirus, the SARS-CoV-2 mutant D614G was also neutralized, indicating cross-neutralization and protection from reinfection by either strain (Garcia-Beltran et al, 2021; Kurahashi et al, 2022).
We found that CoV-2-SNAb in some convalescent persons did not neutralize the virus, although CoV-2-SNAb showed high binding to the S-RBD (Fig. 3, Group 2). This finding indicates that in some cases, neutralization of the CoV-2 virus is performed by mechanism(s) other than CoVid-2-SNAb.
There are many possible mechanisms for the neutralization of the CoV-2 virus: The “cocktail” of neutralizing antibodies (Nabs) in our convalescent patients shows potent neutralization against the SARS-CoV-2 virus, indicating distinct conserved epitopes, including several S-RBDs (Bošnjak et al, 2021; Yan et al, 2020; Yuan et al, 2020) and S-NTD, as well as S1/S2 at the polybasic cleavage site (Brouwer et al, 2020; Chi et al, 2020; Wang et al, 2021), which are essential for S-protein-mediated cell‒cell fusion and virus entry into human lung cells and thus the ability to neutralize the virus (Brouwer et al, 2020; Hoffmann et al, 2020). The S-RBD conformations are involved in the attachment process and are in either an up and slightly rotated configuration or a down configuration, influencing the inactivation of the virus (Henderson et al, 2020). We observed that the “cocktail” of NAbs has variable activity against different VOCs, including some NAbs with loss of neutralization activity or reduced activity against the virus and its variants, in agreement with a previous report (Hingankar et al, 2022). As the distinct epitopes conserved in the S-protein domain are essential for S-protein-mediated cell–cell fusion and virus entry into human lung cells and thus the ability to neutralize the virus, there is potential variation at this domain (Brouwer et al, 2020; Hoffmann et al, 2020). S-RBD and S-NTD together have also been suggested as promising peptides for therapeutic neutralizing antibodies against the SARS-CoV-2 virus (Chi et al, 2020; Wang et al, 2021). The activation of B cells by CD4+ T cells also plays an important role in the elimination of COVID-19 at the onset of symptoms (Farshi et al, 2021; Wang et al, 2020a). In some of the convalescent persons, both elevated and decreased immune-related phagocytic blood cells were found months after the onset of symptoms (Table 3). A series of studies reported B cell responses up to 6 months (Sokal et al, 2021) and that B-/T cell responses developed within 6–8 months (Sherina et al, 2021) in mild and severe COVID-19 patients. Spike-specific memory B cells were more abundant at 6 months after symptom onset (Dan et al, 2021). SARS-CoV-2-specific CD4+ T-cells and CD8+ T-cells declined with a half-life of 3 to 5 months in an integrated manner (Dan et al, 2021).
In a recent study on T cell-mediated responses measured by ELISpot, memory T cells were detected, indicating that memory T cells were not found in all individuals and that no correlation was observed with neutralizing antibody responses, but the vast majority of the individuals showed SARS-CoV-2-specific antibodies and/or T-memory cells (Glück et al, 2022). Today, it is still unclear whether cellular or neutralizing antibody-mediated immunity is the dominant factor in long-term immunity against SARS-CoV-2 (Glück et al, 2022), whether the changes in dengue virus antibody (Hartley et al, 2020), and whether the differences between the serum-neutralizing antibody individual and various vaccines and the effect of pre-SARS-CoV-2 cross-reactive T cells on clinical outcomes are of clinical relevance (Kurahashi et al, 2022; Valdez-Cruz et al, 2021).
To further improve the fight against the COVID-19 pandemic, a combination of vaccines and nonimmunological treatments should also be tested. Such investigations are now ongoing in our research group. It is well known that inflammation is an essential response to the initiation of immune function (Wang et al, 2021). The excessive accumulation of inflammatory cells may lead to cell death (Park et al, 2020; Terpos et al, 2021). It has also been shown that patients infected with SARS-CoV-1, bird flu, or other types of viruses can produce large amounts of specific neutralizing antibodies.
The patients who recovered from mild to moderate coronavirus infection showed increased activation of CD4+ and CD8+ -T cells, antibody-secreting cells, and follicular helper T cells, resulting in the production of novel virus IgM- and IgG-neutralizing antibodies and inhibiting virus replication in vivo (Park et al, 2020; Terpos et al, 2021). The presence of memory B cells in convalescent patients at 6 months postsymptom onset has been reported (den Hartog et al, 2021; Hartley et al, 2020). Higher neutralizing antibody titters have also been reported (Akbar and Gilroy, 2020; Markmann et al, 2021; Valdez-Cruz et al, 2021).
In this study, the CoV-2-SNAb response against authentic SARS-CoV-2 virus was analyzed by CoV-2-SNAb RT-PCR assays using reliable RT-PCR detection assays of the genome sequence of SARS-CoV-2 (Ju et al, 2020). We found that the threshold of S-RBD = 1.0 OD can be used as an initial evaluation of the neutralization effect of CoV-2-SNAb on the virus or for the assessment of vaccination of people.
The microneutralization assay and the focus reduction neutralization test (Zhou et al, 2022) against SARS-CoV-2 are based on the plaque reduction neutralization test (Ju et al, 2020) and are considered the gold standard for measuring neutralizing antibodies in serum/plasma. However, those assays require BSL-3 facilities (Ju et al, 2020). To eliminate the use of BSL-3 facilities, the use of the pseudovirus neutralizing assay (PNA) is a valid alternative to the use of live SARS-CoV-2 virus, but the assay needs to be optimized and standardized (Bewley et al, 2021). We suggest to setting up a project to evaluate the use of the CoV-2-NpAb response neutralizing the CoV-2 virus in convalescent blood donors/vaccinated (Fig. 4).
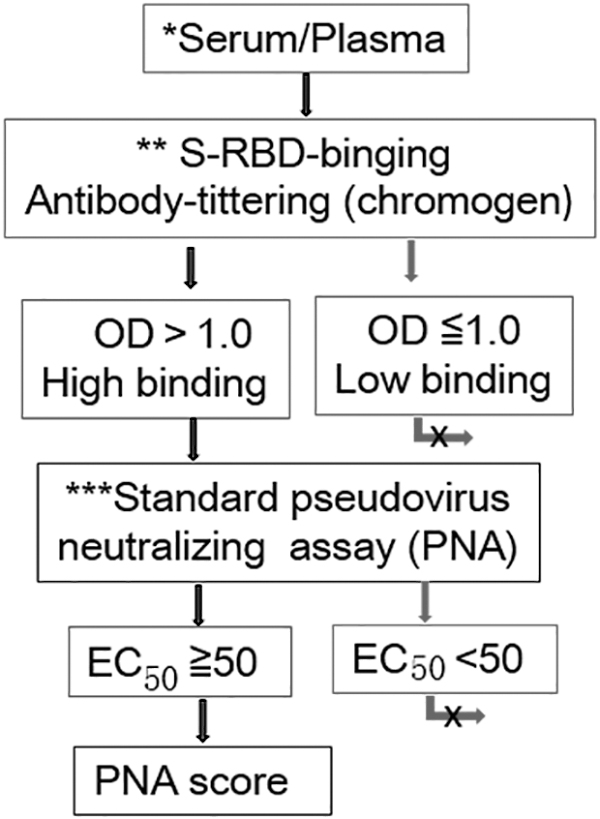
A schematic diagram of a project for assessment of CoV-2-NpAb response neutralizing CoV-2 virus of convalescent blood donors/vaccinated. *Serum: serum samples underwent heat inactivation at 56°C for 30 min and were stored at −80°C until tested. **S-RBD binging antibody titration: chromogen is a sensitive and simple method; the fluorescent assay is also recommended. ***The PNA system will be optimized and standardized in the future. EC50, half maximal effective concentration; PNA, pseudovirus neutralizing assay
In summary, the anti-CoV-2-SNAb neutralized the “WH-Human 1” coronavirus significantly for up to 10 months. We suggest that convalescents should contact their physician 6-month postinfection to test the function of serum neutralizing antibodies and determine whether administering a SARS-CoV-2 vaccine is necessary to prevent the development of severe illness in the future and use the threshold value of S-RBD as an evaluation of the effect of neutralization of the CoV-2 virus or the assessments of vaccination in combination with PNA.
Footnotes
Acknowledgments
We thank the Science and Development Department, Xiangtan City, China, the Centre of Disease Control, Shenzhen, China; the Shenzhen Ellen-Sven Precision Medicine Institute, China; and the Renmin Hospital of Wuhan University and Xiangtan Central Hospital, China for their support.
Authors' Contributions
Conceptualization: J.L., L.Z., S.L., Z.F., J.Z., E.H., and S.S.; methodology, analysis, and data interpretation: R.Z., H.L., P.Z., D.K., C.J., P.G., C.F., S.W., and E.H.; writing article: J.L., E.H., and S.S.; supervision: J.Z. and J.L.; project administration: J.L., L. Z., S.L., R.Z., Z.F., and J.Z.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by the Science and Development Department of Xiangtan City, New Crown Epidemic Emergency Special Project (Project No. SF-YB20201011).
Ethical Conduct of Research
Ethics approval and consent to participate. This study is in accordance with the ethical standards as laid down in the 1964 Declaration of Helsinki and its later amendments. All procedures in the study involving participants of the two hospitals were supervised by the National Ethical Guidelines of China and as a cooperation project approved by the Ethics Committee of Xiangtan Science and Development Department, Xiangtan city (Ethic no.-2020-03-18 No.3).