
Abstract
To study the clinical significance of manganese (Mn) in the serum of children with infectious mononucleosis (IM) caused by Epstein-Barr virus (EBV) infection, we analyzed the correlation between Mn and the cyclic GMP-AMP synthase (cGAS)–stimulator of interferon genes (STING) pathway and explored the immune pathogenesis of EBV infection. Children diagnosed with IM comprised the IM group, and healthy children during the same period were selected as the normal control group. Real-time reverse transcription-polymerase chain reaction was used to detect the mRNA expression levels of cGAS, STING, Tank-binding kinase 1 (TBK1), interferon regulatory factor 3 (IRF3), and related inflammatory factors, and Mn in serum was detected by inductively coupled plasma mass spectrometry. Interferon (IFN)-α and IFN-β expression levels in serum were detected by enzyme-linked immunosorbent assay, and the correlation between Mn levels and clinical manifestations and laboratory tests was analyzed. Mn levels and the expression levels of cGAS, STING, and related inflammatory factors were significantly higher in children with IM than in healthy children. Furthermore, Mn levels in children with IM were positively correlated with the expression levels of cGAS and related inflammatory factor
Introduction
Epstein-Barr virus (ebv) is a common pathogen that causes infectious mononucleosis (IM) in children, and more than 90% of people are infected with EBV at some point in their lives (Kuri et al, 2020). It has been confirmed that EBV is closely related to the occurrence and development of Burkitt's lymphoma, NK/T cell lymphoma, Hodgkin's lymphoma, nasopharyngeal carcinoma, and other tumors, which seriously threaten human health (Liu et al, 2013). The innate immune response is the body's first line of defense against infection, and the cyclic GMP-AMP synthase (cGAS)–stimulator of interferon genes (STING) pathway plays an important role in host resistance to viral infection (Cheng et al, 2020).
The homeostasis of trace elements is very important for biological organisms because they are cofactors involved in many processes. Manganese (Mn), magnesium (Mg), iron (Fe), copper (Cu), and zinc (Zn) are essential for all life. Research in recent years has demonstrated that Mn2+ is a critical substance for host resistance to DNA viruses. Mn2+ also enhances the sensitivity and enzymatic activity of cGAS to double-stranded DNA after virus infection (Wang et al, 2018). EBV is a typical double-stranded DNA virus.
In this study, we examined Mn levels in the serum of children with IM caused by EBV infection, as well as the expression levels of cGAS, STING, Tank-binding kinase 1 (TBK1), interferon regulatory factor 3 (IRF3), interferon-induced proteins with tetratricopeptide repeats 2 (IFIT2), C-X-C motif chemokine ligand 10 (CXCL10), interferon beta 1 (IFNB1), and interferon (IFN). Furthermore, we investigated the clinical significance of Mn in IM and the immune mechanism of EBV infection.
Materials and Methods
Research subjects
Thirty children who were hospitalized between January 2019 and June 2020 at our hospital and were confirmed to have IM caused by EBV infection were selected as the case group (IM group), and 20 healthy children were selected as the normal control group (NC group) during the same period. The IM group inclusion criteria were as follows: (1) IM diagnostic criteria were met (2016); and (2) peripheral blood EBV-DNA load >400 copies/mL. Exclusion criteria were a history of repeated infections, immunodeficiency diseases, genetic metabolic diseases, and incomplete medical records.
Specimen collection
Venous blood was drawn from the enrolled children before the use of antiviral drugs on the day of admission to the hospital.
Blood cell and platelet count detection
The leukocyte count was determined by BC-7500CRP auto hematology analyzers using electrical impedance; The leukocyte classification was determined using fluorescence staining and flow cytometry; Also, the platelet (PLT) count was determined using Sheath flow electrical impedance.
Lymphocyte subpopulation detection
Add 20 μL lymphocyte subset detection reagent (CD3-FITC/CD8-APC-Cy7/CD45PerCP-Cy5.5/CD4-PC7/CD16+CD56-PE/CD19-APC) to the bottom of the absolute counting tube; Pipette 50 μL of fully mixed ethylenediaminetetraacetic acid (EDTA) anticoagulated whole blood by reverse pipetting and add it to the bottom of the absolute counting tube; incubate for 15 min at room temperature, avoid light after shaking and mixing well; Add 450 μL of hemolytic reagent for flow cytometry into each tube and incubate at room temperature, avoid light for 15 min after mixing well; The percentage and absolute number of lymphocyte subsets were obtained and analyzed by flow cytometry (FACS canto II, BD).
Immunoglobulin detection
IgM, IgG, and IgA contents in serum were determined by Mindray BS2000 using immunoturbidimetry.
EBV DNA detection
Venous blood was drawn from each patient and anticoagulated with EDTA, and the EBV-DNA load was detected by real-time polymerase chain reaction (PCR). The reagents were produced by Hunan Shengxiang Biotechnology Co. Ltd (Hunan, China). Samples were analyzed using the Roche LightCycler 480 real-time PCR machine (Roche Pharmaceutical Co. Ltd, Shanghai, China) according to the manufacturer's instructions.
Trace element detection
The concentration of Mn in serum was measured using an inductively coupled plasma high-resolution mass spectrometer (ELEMENT 2). The concentration of Mg in serum was detected by the xylidine blue method with a Hitachi automatic biochemical analyzer 7600, and the reagent was produced by Fujifilm Wako Pure Chemical Corporation. The concentration of Fe in serum was detected by the complexing method, and the reagent was produced by German Desay Diagnostics. Cu concentration in serum was produced by the complexing method, and the reagent was produced by SHINO-TEST. The concentration of Zn in serum was detected by alkaline phosphatase reaction colorimetry, and the reagent was produced by Beijing Century World Biotechnology.
Detection of cGAS, STING, TBK1, IRF3, IFIT2, CXCL10, and IFNB1
The peripheral blood samples were centrifuged in a low temperature high-speed centrifuge at 15,000 g at 4°C for 5 min, and 0.5 mL TRIzol was added to the bottom of the tube to extract total RNA, reverse transcribed to synthesize cDNA. Using 18s as an internal reference, real-time PCR was used to determine the expression levels of cGAS, STING, TBK1, IRF3, IFIT2, CXCL10, and IFNB1. The sequence of primers is shown in Table 1. The expression difference of each gene was compared by the 2−△△Ct method.
Forward and Reverse Primers Used for Real-Time Polymerase Chain Reactions
cGAS, cyclic GMP-AMP synthase; CXCL10, C-X-C motif chemokine ligand 10; IFIT2, interferon-induced proteins with tetratricopeptide repeats 2; IFNB1, interferon beta 1; IRF3, interferon regulatory factor 3; STING, stimulator of interferon genes; TBK1, Tank-binding kinase 1.
Detection of IFN-α and IFN-β
IFN-α and IFN-β cytokines in serum were detected by enzyme linked immunosorbent assay kit (Neobioscience, Shenzhen, China). All procedures were conducted according to the manufacturer's instructions.
Statistical methods
Statistical analysis was performed using SPSS 24.0 (IBM Corp., Armonk, NY). Data conforming to a normal distribution were expressed as mean ± SD. Student's t-test was used for comparison between groups, and Pearson correlation was used for correlation analysis. Data not conforming to the normal distribution were expressed as the median (IQR), and the Mann–Whitney U test was used for pairwise comparisons between groups, while Spearman's correlation was used for correlation analysis. The enumeration data were expressed as percentages, and the chi-squared test or Fisher's exact test was used between groups. p < 0.05 was considered statistically significant.
Results
There were 30 children in the IM group, which comprised 17 males and 13 females with an average age of 4.62 ± 0.97 years, and 20 children in the NC group, with 10 males and 10 females with an average age of 4.56 ± 1.33 years. There was no significant difference in sex and age between the two groups (p > 0.05).
Comparison of routine blood tests between the IM and NC groups
White blood cell (WBC) and lymphocyte (LYM) counts in the IM group were significantly higher than in the NC group (p < 0.05), and PLT count in the IM group was significantly lower than that in the NC group (p < 0.05). There was no significant difference in Neutrophil count between the two groups (p > 0.05; Table 2).
Comparison of Routine Blood Tests Between the Infectious Mononucleosis and Normal Control Groups
Indicates significant p value.
IM, infectious mononucleosis; LYM, lymphocyte; NC, normal control; NEU, neutrophil; PLT, platelet; WBC, white blood cell.
Comparison of immune function in the IM and NC groups
The absolute counts of CD3+, CD3+CD4+, and CD3+CD8+ T cells in the IM group were higher than those in the NC group, and the ratio of CD4+/CD8+ in the IM group was lower than that in the NC group (p < 0.05). IgA, IgG, and IgM levels in the IM group were significantly higher than those in the NC group (p < 0.05; Table 3).
Comparison of Immune Function in the Infectious Mononucleosis and Normal Control Groups
Indicates significant p value.
Comparison of trace elements in the IM and NC groups
Compared with the NC group, Mn and Cu levels in the IM group were significantly higher than those in the NC group, while Mg and Fe in the IM group were significantly lower than those in the NC group (both p < 0.05). The difference in Zn between the two groups was not statistically significant (p > 0.05; Table 4).
Comparison of Trace Elements in the Infectious Mononucleosis and Normal Control Groups
Indicates significant p value.
Cu, copper; Fe, iron; Mg, magnesium; Mn, manganese; Zn, zinc.
Comparison of cGAS, STING, TBK1, and IRF3 mRNA expression levels in the IM and NC groups
Compared with the NC group, the mRNA expression levels of cGAS, STING, IFIT2, CXCL10, and IFNB1 in the IM group were significantly higher than those in the NC group (p < 0.05; Table 5 and Figs. 1 and 2). However, the differences in the mRNA expression levels of TBK1 and IRF3 between the two groups were not statistically significant (p > 0.05; Table 5 and Fig. 1).
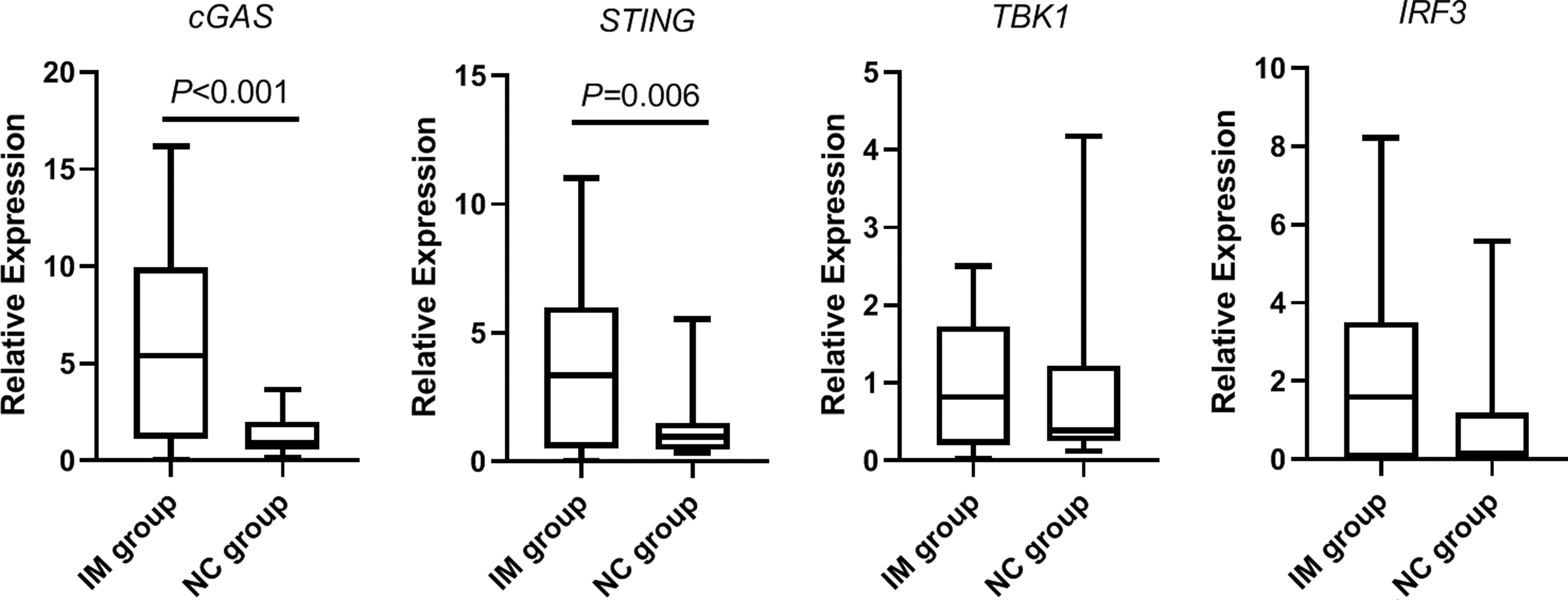
Comparison of cGAS, STING, TBK1, and IRF3 mRNA expression levels in the IM and NC groups. cGAS, cyclic GMP-AMP synthase; IM, infectious mononucleosis; IRF3, interferon regulatory factor 3; NC, normal control; STING, stimulator of interferon genes; TBK1, Tank-binding kinase 1.
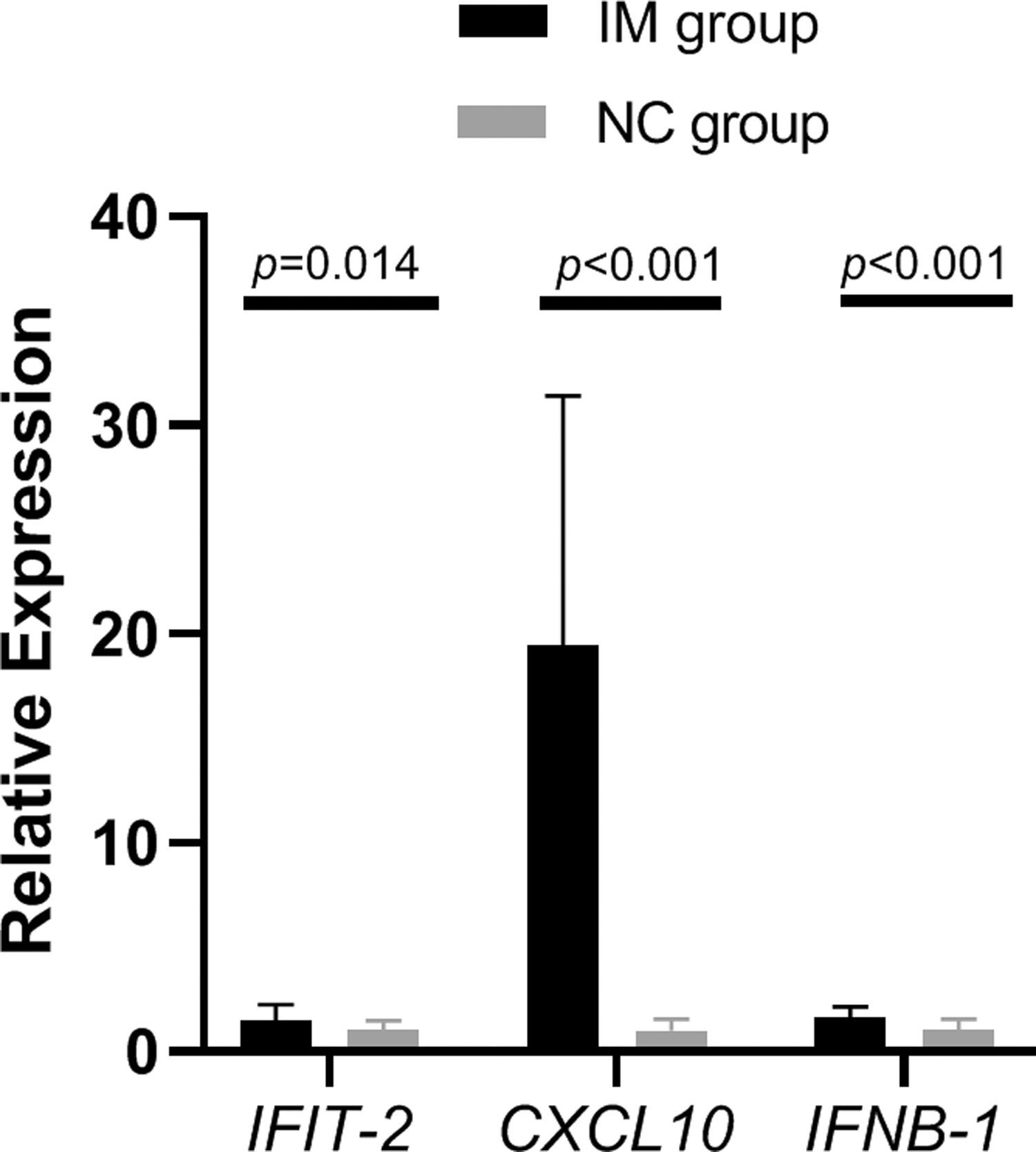
Comparison of IFIT2, CXCL10, and IFNB1 mRNA expression levels in the IM and NC groups. CXCL10, C-X-C motif chemokine ligand 10; IFIT2, interferon-induced proteins with tetratricopeptide repeats 2; IFNB1, interferon beta 1.
Comparison of cGAS, STING, TBK1, IRF3, IFIT2, CXCL10, and IFNB1 mRNA Expression Levels in the IM and NC Groups
Indicates significant p value.
Comparison of IFN-α and IFN-β between the IM and NC groups
Compared with the NC group, IFN-α and IFN-β expression in the IM group were significantly higher (p < 0.05; Table 6).
Comparison of Interferon-α and Interferon-β in the Infectious Mononucleosis and Normal Control Groups
Indicates significant p value.
IFN, interferon.
Correlation of trace elements with cGAS, IFIT2, CXCL10, IFNB1, IFN-α, and IFN-β in the IM group
Mn levels in the IM group were positively correlated with the expression of cGAS, IFIT2, CXCL10, IFNB1, IFN-α, and IFN-β (r = 0.573, r = 0.624, r = 0.683, r = 0.663, r = 0.692, and r = 0.409, respectively; all p < 0.05). There was no correlation between the expression of STING (p > 0.05). Furthermore, Mg, Fe, and Cu levels in the IM group were not correlated with the expression of cGAS, STING, IFIT2, CXCL10, IFNB1, IFN-α, and IFN-β (p > 0.05; Table 7).
Correlation of Trace Elements with cGAS, STING, IFIT2, CXCL10, IFNB1, IFN-α, and IFN-β in the IM Group
Indicates significant p value.
Correlation between Mn and other indicators in the IM group
Mn levels in the IM group were significantly positively correlated with WBC count, LYM count, CD3+ T cell absolute count, and CD3+CD8+ T cell absolute count (r = 0.486, r = 0.478, r = 0.468, and r = 0.543, respectively; all p < 0.05). Mn levels in the IM group were negatively correlated with the ratio of CD4+/CD8+ (r = −0.497; p < 0.05). There was no correlation between Mn levels and PLT, CD3+CD4+ T cell absolute count, IgA, IgG, and IgM in the IM group (p > 0.05; Table 8).
Correlation Between Manganese and Other Indicators in the Infectious Mononucleosis Group
Indicates significant p value.
Discussion
EBV is a human lymphotropic herpes virus that was first discovered in 1964 and is a pathogen in various diseases, with more than 90% of people infected at some point during the course of their life (Kuri et al, 2020). After EBV enters the body, it binds to CD21 receptor on the membrane of B lymphocytes via gp350 (Cui et al, 2013; Hutt-Fletcher, 2007), upon which the antigenicity of B cells changes and stimulates the proliferation of lymphocytes (Cohen, 2018). In addition, there is an EBV epitope on the platelet membrane glycoprotein GPIIb. After EBV infects the body, antiviral antibodies are produced that can bind to platelets, which may lead to the destruction of platelets and clearance through the reticuloendothelial system (Jin et al, 2012; Wu et al, 2013). In the present study, WBC and LYM counts were higher and PLT counts were lower after EBV infection.
The body's cellular immunity plays an important role in resisting viral infection. After EBV infects the body, it enters cells by binding to CD21 on the surface of B lymphocytes. The proliferation of B lymphocytes can induce the activation of T lymphocytes, thereby eliminating the infected B lymphocytes (Li et al, 2012). Among them, the most significant increase is in CD8+ T lymphocytes (Wang et al, 2017). In the current study, CD3+, CD4+, and CD8+ T lymphocyte counts in the IM group were higher than those in the NC group, and the CD4+/CD8+ ratio was significantly lower than that in the NC group. Activated B lymphocytes produce anti-infection antibodies IgA, IgG, and IgM, which play an important role in the body's anti-infection (Fleisher and Oliveira, 2004). In this study, the IgA, IgG, and IgM levels of the children in the IM group were significantly higher than those in the NC group.
Trace elements are important factors that affect the immune function of the body, and their imbalance can affect human health and even cause disease. Mn, Cu, Zn, Fe, and Mg are essential for all life because most enzymes require metal cofactors (Gröber et al, 2015; Hood and Skaar, 2012; Kysenius et al, 2019). Mg acts as a cofactor for adenosine triphosphate, nucleic acids, and various enzymes in the body, and has various functions such as enhancing tolerance to oxidative stress and signal transduction. Fe directly participates in cellular respiration, biological oxidation, and other processes, and affects the activities of various enzymes. When the body is deficient in Mg and Fe, it will lead to a decline in immune function, increasing the risk of viral infection (Hu et al, 2017; Jing and Ren, 2019).
Elevated levels of Cu in serum compared with the control group may be related to biological defense mechanisms (Hu et al, 2021), and hosts can exploit Cu toxicity as an efficient innate response to infection (Fu et al, 2014). As an essential trace element in the human body, Mn is ubiquitously present in all cells of the human body (Mercadante et al, 2019; Scheiber et al, 2019). In recent years, studies have shown that Mn ions can function as danger-related molecular patterns or alarmins, transmit signals of virus invasion, and activate innate immunity to resist virus invasion (Wei et al, 2020).
This study found that the levels of Mg and Fe in the IM group were lower than those in the NC group, and the levels of Cu and Mn in the IM group were higher than those in the NC group. Activation and proliferation of B lymphocytes induced by EBV infection causes an increase in Zn content (Ollig et al, 2019). The results of this study showed that EBV infection caused an increase in serum Zn content, but the difference was not statistically significant, which may be related to the different disease course and the size of the sample.
When dsDNA infection occurs, the cGAS–STING signaling pathway can recognize cytoplasmic DNA and make the body produce IFN, thereby exerting an antiviral innate immune response (Ma and Damania, 2016). In this study, the expression levels of cGAS, STING, CXCL10, IFIT2, and IFNB1 in the IM group were higher than those in the NC group. Moreover, the levels of IFN-α and IFN-β in the IM group were significantly higher than those in the NC group, suggesting that cGAS, STING, type I IFN, and other inflammatory cytokines are involved in the immune mechanism of IM caused by EBV infection. TBK1 is a member of the noncanonical inhibitor of nuclear factor kappa-B kinase family. After infection with DNA viruses, STING binds to TBK1 and promotes its phosphorylation at serine 172 on the activation loop of TBK1 (Hu et al, 2020). Activated TBK1 phosphorylates IRF3, leading to its homodimer formation and translocation into the nucleus, driving the expression of antiviral IFN (Ahmad et al, 2016). There were no significant changes in the mRNA expression levels of TBK1 and IRF3 in the IM group, which may be related to the activation of the proteins (Li et al, 2019; Zhang et al, 2019).
Studies have reported that Mn generally exists in the form of divalent ions, and Mn2+ can resist DNA virus infection by increasing the sensitivity of the DNA sensor cGAS and its downstream adapter protein STING (Wang et al, 2018; Wei et al, 2020). This study also found that the Mn content in serum was positively correlated with cGAS, IFIT2, CXCL10, IFNB1, IFN-α, and IFN-β. Furthermore, Mn levels were positively correlated with WBC count, LYM count, CD3+, and CD8+, and negatively correlated with CD4+/CD8+, which suggested that the changes in Mn in children with IM may be related to immune cells in vivo. It has been reported that lymphocytes and macrophages may ingest Mn ions released extracellularly to enhance the innate immune response in cells (Wei et al, 2020).
Zhang et al (2021) developed a nano-Mn adjuvant (Mn salt, MnJ) with stable colloidal properties. MnJ can potently activate cellular immune responses, especially cytotoxic T lymphocytes. However, Gram et al showed that human B lymphocytes express cGAS, STING, and the signal transduction components TBK1 and IRF3, but they are unable to secrete type I IFNs after cytoplasmic DNA exposure (Gram et al, 2017). Whether T lymphocytes can induce the production of type I IFN through this pathway and how Mn2+ regulates this pathway require further study.
Conclusions
In summary, this study showed that serum Mn in children with IM caused by EBV infection was increased, and inflammatory cytokines such as cGAS, STING, and type I IFN were involved in the immune mechanism of IM caused by EBV infection. The limitations of this study include the small sample size and the fact that the clinical samples were easily affected by individual differences such as age and immune function. Moreover, this study only detected cGAS, STING, TBK1, IRF3, and related inflammatory factors in children with IM, and these findings have not been verified in vitro. In the future, EBV can be used to infect cells or build animal models with overexpression or inhibition of critical molecules for further exploration.
Footnotes
Acknowledgment
We thank H. Nikki March, PhD, from Liwen Bianji (Edanz) (
Data Availability
All data are available.
Authors' Contributions
J.S. performed the molecular genetic studies, participated in sequence alignment, and drafted the article; Y.L. selected the topic; D.Z. and M.Y. performed data analysis; W.Z. reviewed and revised the article. All authors read and approved the final article.
Author Disclosure Statement
No competing financial interests exist.
Ethics Approval
This study was conducted with the approval of the Institutional Human Ethics Committee of the Children's Hospital of Soochow University (article ethics approval number: 2019LW023).
Funding Information
The study was supported by the Scientific and Technological Development Project (Livelihood Technology) of Suzhou (no. SYS2019085).