
Abstract
Dengue virus (DENV) is the etiological agent of dengue, the most important mosquito-transmitted viral disease of humans worldwide. Enzyme-linked immunosorbent assays (ELISAs) designed to detect DENV IgM are commonly used for dengue diagnosis. However, DENV IgM is not reliably detected until ≥4 days after illness onset. Reverse transcription–polymerase chain reaction (RT-PCR) can diagnose early dengue but requires specialized equipment, reagents, and trained personnel. Additional diagnostic tools are needed. Limited work has been performed to determine whether IgE-based assays can be used for the early detection of vector-borne viral diseases, including dengue. In this study, we determined the efficacy of a DENV IgE capture ELISA for the detection of early dengue. Sera were collected within the first 4 days of illness onset from 117 patients with laboratory-confirmed dengue, as determined by DENV-specific RT-PCR. The serotypes responsible for the infections were DENV-1 and DENV-2 (57 and 60 patients, respectively). Sera were also collected from 113 dengue-negative individuals with febrile illness of undetermined etiology and 30 healthy controls. The capture ELISA detected DENV IgE in 97 (82.9%) confirmed dengue patients and none of the healthy controls. There was a high false positivity rate (22.1%) among the febrile non-dengue patients. In conclusion, we provide evidence that IgE capture assays have the potential to be explored for early diagnosis of dengue, but further research is necessary to address the possible false positivity rate among patients with other febrile illnesses.
Introduction
Dengue is a mosquito-borne viral disease that has a devastating impact on human health worldwide (Gubler, 2011; Guzman et al, 2016; Halstead, 2019). The etiological agent is dengue virus (DENV), which belongs to the genus Flavivirus (family Flaviviridae) (Simmonds et al, 2017). Current estimations indicate that 4 billion people are at risk of contracting dengue, with an average of 390 million new DENV infections every year (Brady and Hay, 2020). DENV occurs as four serotypes, designated DENV-1, DENV-2, DENV-3, and DENV-4. Infection with one serotype confers protective immunity against the homologous, but not heterologous, serotype. Secondary DENV infection greatly increases the risk of contracting severe disease due to possible antibody-dependent enhancement of infection (Halstead, 2014; Halstead et al, 2010; Sun et al, 2019).
DENV infections are usually asymptomatic or cause a self-limiting febrile illness accompanied by various other nonspecific symptoms, such as myalgia, arthralgia, headache, retro-orbital pain, and rash (Gubler, 2011; Guzman et al, 2016; Halstead, 2019). Symptoms can escalate to severe life-threatening complications, such as hemorrhagic manifestations and organ failure, particularly among individuals with secondary DENV infections (Halstead, 2014; Hu et al, 2013). Because of the physiological alterations that occur during DENV infections, rapid virus detection is critical (Md-Sani et al, 2018; Niriella et al, 2020). Dengue mortality can be reduced by up to 50% when diagnosis occurs promptly (Omar et al, 2018; Philip Samuel and Tyagi, 2006; WHO, 2012).
The DENV IgM capture enzyme-linked immunosorbent assay (MAC-ELISA) is often used for dengue diagnosis because of its high sensitivity and easiness to perform (WHO, 2009). The MAC-ELISA is often used in tandem with the DENV IgG capture ELISA in epidemiological studies (Lima et al, 2012). One limitation of these assays is that they cannot reliably detect dengue within the first few days of illness. DENV IgM and IgG are not usually detectable until 4–5 and 14 days after symptom onset, respectively (Koraka et al, 2001; Vazquez et al, 2014). Early detection of dengue is commonly achieved by reverse transcription–polymerase chain reaction (RT-PCR).
One limitation of this technique is that it requires specialized reagents and trained personnel. DENV NS1 detection assays are widely used for the early detection of dengue (Guzmán and Kourí, 2004; Muller et al, 2017; Raafat et al, 2019). NS1 is an attractive target because it is secreted from infected cells as early as 1 day post-infection (Muller and Young, 2013). Many NS1 detection assays have been developed, with sensitivities as low as 34% and as high as 96% (Bessoff et al, 2008; Blacksell et al, 2012; Castro-Jorge et al, 2010; Duong et al, 2011; Guzman et al, 2010; Kumarasamy et al, 2007; Najioullah et al, 2011; Ramirez et al, 2009). Virus isolation is the gold standard technique for dengue diagnosis, but it is laborious and time-consuming as it takes longer to perform compared with the aforementioned techniques.
IgE production occurs when an allergen acts as an antigen, resulting in an allergen-induced humoral response (Kelly and Grayson, 2016; Prussin and Metcalfe, 2003). IgE is distinguished from other antibody classes by its two heavy ɛ chains that bind with high affinity to a specific receptor (FcɛR1) on the plasma membrane of mast cells and basophils. Production of IgE is tightly regulated and linked to interleukin-4 and prostaglandin stimulation. Although the role of IgE in the development of allergic reactions after exposure to insect bites is well described (Barros et al, 2016; Singh and Mann, 2013), considerably less work has been performed to characterize the IgE responses that occur during DENV infection (Bachal et al, 2015; Inokuchi et al, 2018; Míguez-Burbano et al, 1999; Vazquez et al, 2007).
Mosquito saliva allergens can bind to specific ligands and DENV virions (Calvo et al, 2006; Cao-Lormeau, 2009), which could affect the kinetics of the host immune response during infection. Furthermore, antibody-mediated penetration of DENV into basophils and mast cells triggers the production of bioamines that have potent vasoactive activity, leading to vascular permeability (Mabalirajan et al, 2005). High levels of anti-DENV IgE have been detected in dengue patients at illness onset, persisting for several days after defervescence (Koraka et al, 2003). This phenomenon could be a consequence of the host immune response elicited during mosquito feeding and should be exploited for the development of a novel diagnostic tool for dengue disease.
In this study, we developed an ELISA for the detection of DENV IgE and assessed its performance for early dengue detection.
Materials and Methods
Study population and sample collection
The study was performed using sera from the repository maintained in the Laboratorio de Arbovirologia in the Dr. Hideyo Noguchi Regional Research Center in Merida, Mexico. Sera were collected from 260 human subjects in Merida in 2015–2017 during a community service program for dengue detection coordinated by researchers from the Universidad Autonoma de Yucatan and the Korean Cooperation for Infectious Diseases. Human subjects were separated into three groups (designated as groups 1–3). Group 1 consisted of 117 patients who met the criterion for dengue fever established by the World Health Organization (WHO) in 1997 (WHO, 1997). DENV RNA was detected in the sera of all patients by endpoint serotype-specific RT-PCR. The serotypes were identified as DENV-1 (57 patients) and DENV-2 (60 patients).
Sera were collected on the day 1 as the onset of illness or day 2 (first day after onset), day 3 (2nd day after onset), day 4 (3rd day after the onset of illness), and day 5 (4th day after the onset of illness) with 12, 26, 37, 29, and 13 patients, respectively. Group 2 consisted of 113 dengue-negative patients with febrile illness of undetermined etiology. These patients met the clinical criterion of dengue, but not the laboratory criterion of dengue because DENV RNA and IgM were not detectable in their sera. Sera were collected from these patients 3–10 days after illness onset. Group 3 consisted of 30 asymptomatic negative controls who were negative for both DENV RNA and IgM. All human subjects provided written consent that their sera could be used for research purposes. All sera were blind-coded, and patient-identifying information was removed. The study was approved by the institutional review boards of each of the participating research institutions.
Reverse transcription–polymerase chain reaction
Total RNA was extracted from each serum using the QIAamp Viral RNA Mini Kit (Qiagen, Germantown, MD, USA) according to the manufacturer's instructions. An aliquot of each total RNA was tested for DENV RNA using a single-step semi-nested RT-PCR where the amplification was first performed using DENV-specific consensus primers, followed by serotype-specific primers, as previously described (Seah et al, 1995). Amplicons were visualized by 2% agarose gel electrophoresis and ethidium bromide staining.
Preparation of viral antigen
Viral antigen for the ELISAs was prepared from all DENV serotypes and pooled before use. Antigen was produced by inoculating subconfluent monolayers of Aedes albopictus (C6/36) cells in 75-cm2 culture flasks with virus at a multiplicity of infection of 0.01. Cell culture media was replaced at 6 days post-infection, and cells were scraped from the flasks at 14 days post-infection. Antigens were precipitated using PEG 8000 following established protocols (Keasey et al, 2017). Precipitates were resuspended in nuclease-free distilled H2O to give a final concentration of 1.0 mg/μL, and equal volumes of each were pooled. Uninfected cells were prepared in parallel to prepare negative control antigens.
MAC-ELISA
MAC-ELISAs were performed as previously described, with minor modifications (Loroño-Pino et al, 2004). Briefly, sheep anti-human IgM (Sigma–Aldrich, St. Louis, MO, USA) was used as the capture antibody, and 6B6C-1 (BioRad Laboratories, Hercules, CA, USA) was used as the detector antibody. The MAC-ELISA antigen was prepared from all four DENV serotypes and pooled before use. A serum sample was considered positive for antibodies to DENV if the optical density (OD) value was at least twofold higher than the mean OD for the negative control sera.
IgE capture ELISA
The inner 60 wells of 96-well flat-bottom microtiter plates (Nalge Nunc International, Rochester, NY, USA) were coated with 50 μL (30 ng) of goat anti-human IgE (KPL, Gaithersburg, MD, USA) in carbonate–bicarbonate buffer, pH 9.6 (Sigma–Aldrich). Coated plates were incubated overnight at 4°C. Plates were washed three times with 250 μL of wash buffer (phosphate-buffered saline [PBS], pH 7.5, containing 0.1% Tween 20), then 100 μL of blocking buffer (PBS containing 0.1% Tween 20 and 4% skim milk) was added to each well, and incubated for 15 min at room temperature. Next, 50 μL of serum diluted 1:40 in PBS containing 0.5% bovine albumin was added to each well and incubated for 1 h at 37°C.
Plates were washed three times and 50 μL (200 ng) of pooled antigen prepared from all four DENV serotypes was added to each well and incubated for 1 h at 37°C. Plates were washed and 50 μL of anti-flavivirus 4G2 antibody (0.1 ng/mL) labeled with horseradish peroxidase (Novus Biologicals, Littleton, CO, USA) diluted in PBS with 0.5% bovine albumin was added to each well and incubated at 37°C for 1 h. After five more washes, equal volumes of 2,2′ azino-bis(3-ethylbenzthiazoline-6-sulfonic acid) (ABTS) and peroxidase solutions from the ABTS Microwell Peroxidase Substrate System (KPL) were mixed and 100 μL added to each well. Plates were incubated at 37°C for 30 min, and ODs were measured at a wavelength of 450 nm. All serum samples were tested in triplicate.
Data analysis
Statistical analyses were performed using the IBM SPSS Statistics version 22 software for Windows (IBM Corporation, Armonk, NY, USA). Results were considered significant when p ≤ 0.05. ELISA results were analyzed using a receiver operating characteristic (ROC) curve to determine the optimal OD value for DENV IgE detection. OD ratios of P/N were calculated, where P represents the OD of the test serum and N represents the mean OD of the negative controls. Sera with OD ratios of ≥1.4 were considered positive (Vazquez et al, 2014). The Kruskal–Wallis test was used to compare OD ratios between different groups. The Wilcoxon test was used to analyze the OD ratios of dengue patients from different demographic groups. Sensitivity was calculated as (number of DENV IgE-positive dengue patients/total number of dengue patients) × 100. Specificity was calculated as (number of DENV IgE-negative non-dengue patients/total number of non-dengue patients) × 100.
Results
Sample population
The capture ELISA was evaluated using sera from three groups of human subjects.
Group 1 consisted of 117 dengue fever patients, group 2 consisted of 113 non-dengue patients with febrile illness of unknown origin, and group 3 consisted of 30 otherwise healthy controls. Group 1 contained 61 (52.1%) females and 56 (47.9%) males. The mean age was 22.9 ± 10.8 years (22.5 ± 10.8 years for females and 23.3 ± 10.9 years for males). RT-PCR revealed that the serotypes responsible for the infections were DENV-1 (57 patients) and DENV-2 (60 patients). Sera were collected on the day of illness onset (designated day 1) or one of the 4 days that followed (days 2 to 5), with 12, 26, 37, 29, and 13 patients, respectively.
Patients were diagnosed using the criterion for dengue fever established by the WHO in 1997. Group 2 contained 64 (56.6%) females and 49 (43.4%) males. The mean age was 28.2 ± 14.1 years (30.6 ± 14.7 years for females and 25.0 ± 12.7 years for males). Sera were collected 3–10 days after illness onset and none contained DENV RNA or IgM. Group 3 contained 14 (46.6%) females and 16 (53.4%) males, but their ages were not documented. All these individuals were negative for DENV RNA and DENV IgM.
Optimization of the DENV IgE capture ELISA
For the development of the DENV IgE capture ELISA, various parameters were optimized.
We selected 50 μL of capture antibody (goat anti-human IgE) because it showed the best signal-to-noise ratio. We selected the amount of viral antigen and an anti-flavivirus antibody that showed negligible binding to the capture antibody when tested by checkboard titration. Various dilutions of the anti-flavivirus antibody were also tested in ELISAs performed using antigens harvested from uninfected control cells to monitor for potential cross-reactivity and high signal-to-noise ratios. Additional optimization was performed using reference positive and negative samples; for example, sera from convalescent dengue patients who presumably had DENV-specific IgE and sera from select healthy controls.
Diagnostic performance of the DENV IgE capture ELISA
The mean OD of the sera obtained from the dengue patients (0.106 ± 0.034) was significantly higher than those of the healthy controls (0.053 ± 0.005) and non-dengue patients (0.051 ± 0.003) (χ 2 = 124.67, df = 2, p = 0.00) (Fig. 1). The results are expressed as P/N values and analyzed by the ROC curve (Fig. 2). Sera with OD ratios of ≥1.4 were considered positive. Using this diagnostic criterion, the DENV IgE capture ELISA differentiated between most dengue patients and healthy controls. Ninety-seven (82.3%) sera from dengue patients were positive for DENV IgM, while none of the sera from healthy controls was positive. However, there was a high rate of false positives among the non-dengue febrile patients, with 25 (22.1%) yielding apparent positive test results. Therefore, the sensitivity and specificity of our assay were 82.3% and 77.9%, respectively (Table 1).

Box plot diagram comparing the mean optical densities of the sera from healthy controls, non-dengue febrile patients, and dengue patients. Horizontal black bars indicate median values. Boxes represent Q1–Q3. Vertical lines indicate standard deviations. *Significantly different values, as determined by the Kruskal–Wallis test, followed by the Bonferroni test (p ≤ 0.05).
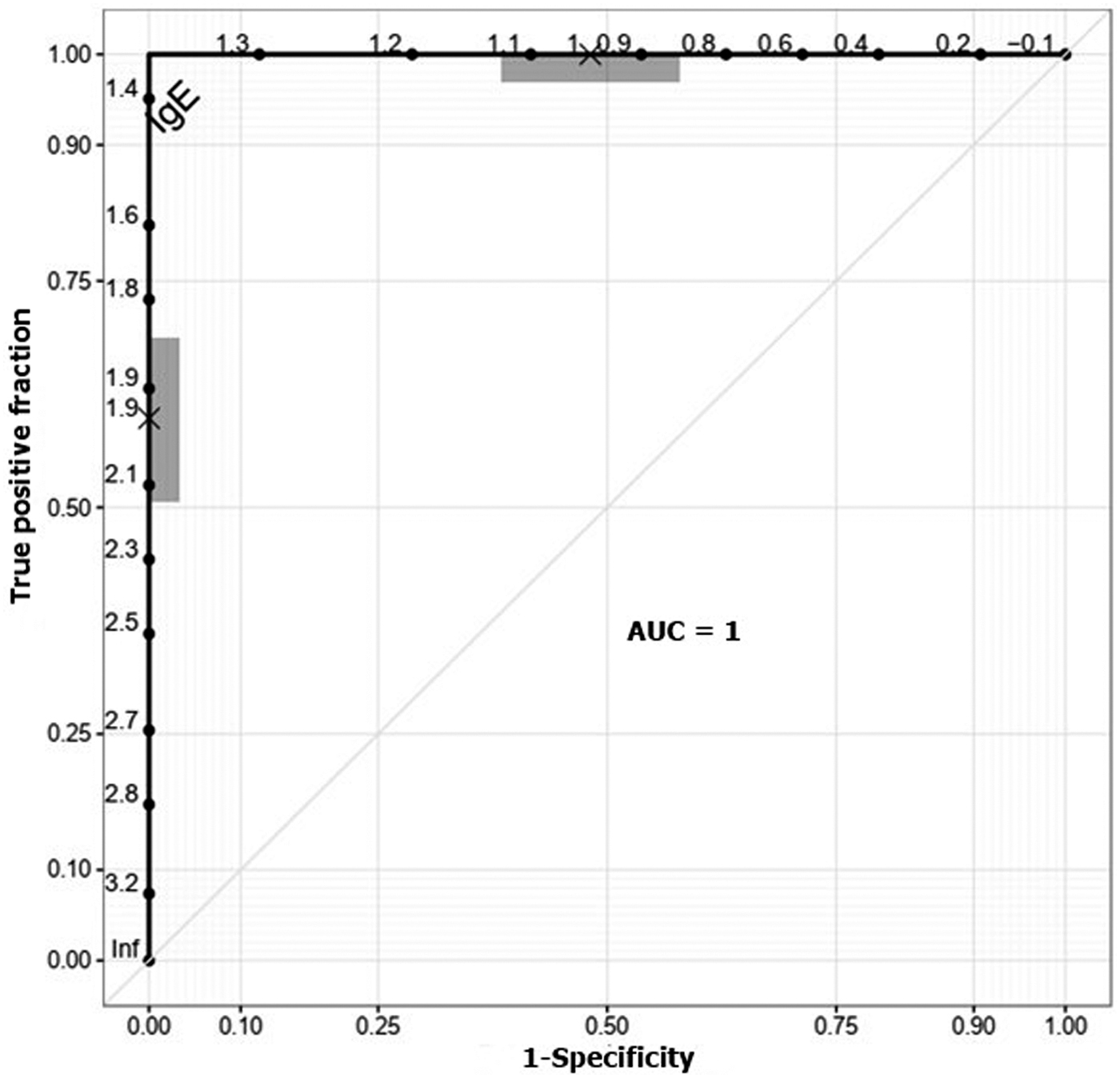
Receiver operating characteristic curve indicating the optimal cutoff value of the DENV IgE capture ELISA. AUC, area under the receiver operating characteristic curve; DENV, dengue virus; ELISA, enzyme-linked immunosorbent assay.
Performance of the Dengue Virus IgE Capture Enzyme-Linked Immunosorbent Assay
ELISA, enzyme-linked immunosorbent assay.
There was no significant difference in the performance of the ELISA when dengue patients were compared according to the time of sera collection (day 1 to 5; p = 0.5871), age (≤20 or >20 years; p = 0.4184), sex (male or female; p = 0.5871), or serotype (DENV-1 or DENV-2; p = 0.5633) (Fig. 3). There was also no significant difference in the performance of the ELISA when dengue patients were compared according to disease severity (dengue fever or dengue hemorrhagic fever [DHF]; p = 0.3145), although the small number of DHF patients (n = 7) did not allow for a robust statistical analysis.
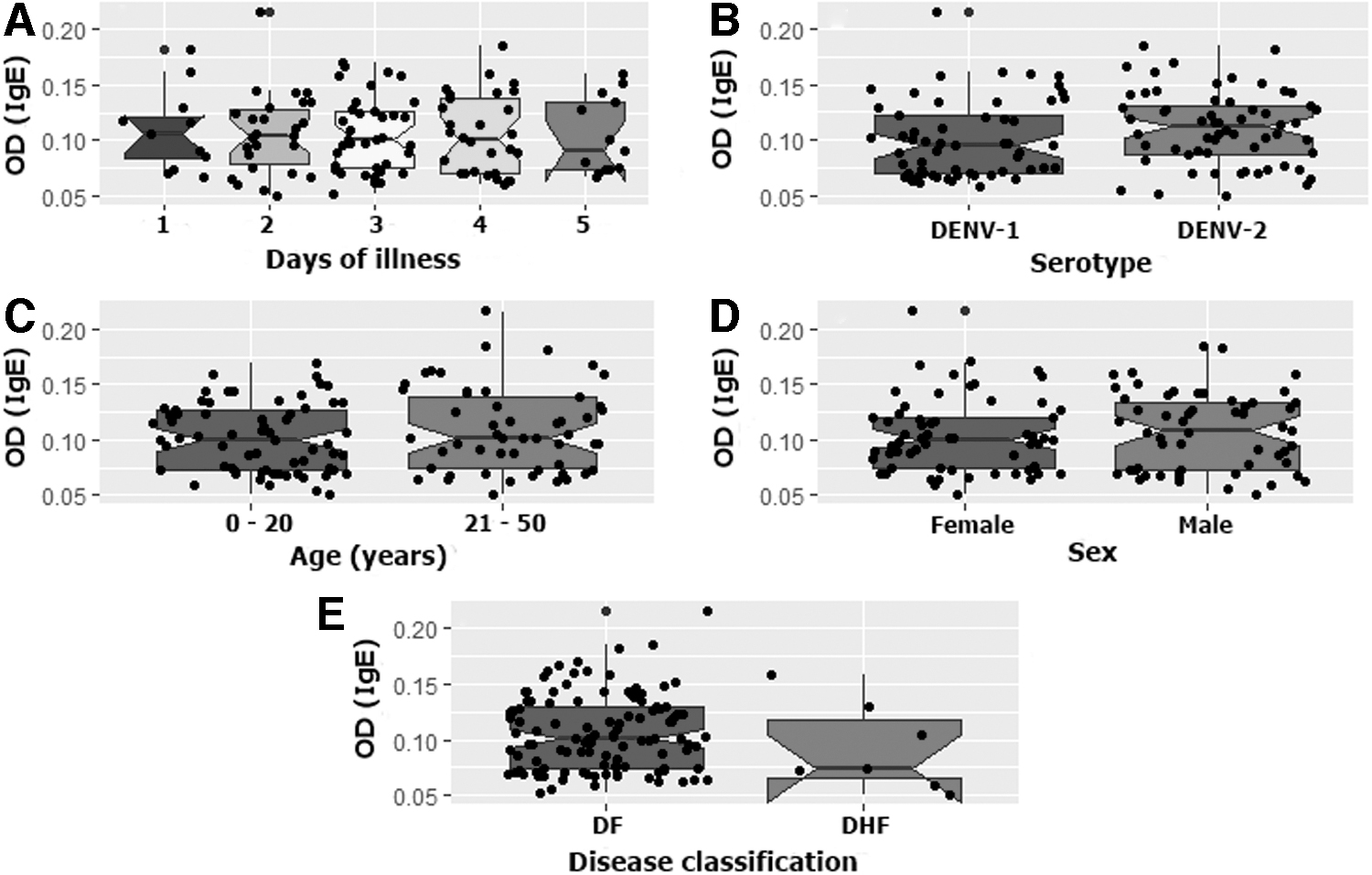
Comparison of DENV IgE presence in dengue patients according to select demographic variables and virus serotype where
Discussion
IgE is an important component of the antiviral immune response (Kelly and Grayson, 2016). Findings from the present study concur with earlier studies demonstrating IgE responses in dengue (Abubakar et al, 1997; Bachal et al, 2015; Inokuchi et al, 2018; Pavri, 1989; Pavri et al, 1980; Shaikh et al, 1983; Vazquez et al, 2014; Vazquez et al, 2007; Vazquez et al, 2005). IgE responses during viral infections have been best characterized for respiratory pathogens, such as a respiratory syncytial virus (RSV) and human rhinoviruses (HRVs) (Agrawal et al, 2014; Kantor et al, 2016; Tam et al, 2013; Welliver et al, 1986; Welliver et al, 1981). Recurrent wheezing is more common among RSV-infected infants with high levels of RSV-specific IgE (Welliver et al, 1986; Welliver et al, 1981).
HRV-specific IgE is produced during HRV infections and potentially mediates the severity of virus-induced asthma exacerbation (Agrawal et al, 2014; Tam and Grayson, 2017; Tam et al, 2013). IgE production during infections with non-respiratory viral pathogens, such as human immunodeficiency, hepatitis, and herpes simplex viruses, has also been studied (Cabanillas et al, 2020; Fletcher et al, 2000; Silva et al, 2021; Vierucci et al, 1984). Less is known about the contribution of IgE during flavivirus infections, and most work has been performed on dengue and Japanese encephalitis viruses (Abubakar et al, 1997; Bachal et al, 2015; Inokuchi et al, 2018; Koraka et al, 2003; Pavri, 1989; Pavri et al, 1980; Shaikh et al, 1983; Vazquez et al, 2014; Vazquez et al, 2007; Vazquez et al, 2005).
One particular gap in our knowledge is the utility of IgE-based assays for flavivirus diagnosis, leading us to evaluate whether they can be used for the detection of early dengue. Our ELISA exhibited a sensitivity of 82.3%, which can be considered high since the sera were collected within the first 5 days of illness. DENV IgM especially in secondary dengue is often not detected in dengue patients at this time (Hu et al, 2011; Suleman et al, 2016; Teoh et al, 2015; Wang and Sekaran, 2010). DENV IgM was identified in 42.9% of dengue patients in China at 3 days post-illness onset (Hu et al, 2011). Less than 60% of patients from Malaysia with confirmed dengue contained detectable DENV IgM at 4 days post-illness onset (Wang and Sekaran, 2010).
Our assay displayed a high rate of potentially false positivity (22.1%) among non-dengue febrile patients. Many human viruses increase total IgE production (Alexeyev et al, 1994; Bahna et al, 1984; Frick, 1986; Koraka et al, 2003; Mazza et al, 1995; Skoner et al, 1995), and thus, the lack of specificity could be because some IgE bound nonspecifically to the DENV antigen. An increase in DENV-reactive IgE was detected in non-dengue patients with acute febrile illness in a hospital-based prospective study in Vietnam (Inokuchi et al, 2018). Another explanation is that our assay displays increased sensitivity compared with the RT-PCR and that a subset of patients assumed to have an illness of undetermined etiology could actually had DENV infections.
This explanation is unlikely because DENV viremias are high during the first few days of illness onset and RT-PCRs for dengue diagnosis are usually highly sensitive (Guzman et al, 2016; Guzman et al, 2010; Raafat et al, 2019; Vaughn et al, 2000). Another explanation is that a continuous cold chain was not maintained for some sera, resulting in the degradation of viral RNA, but not viral antigen as it is more stable (Rainen et al, 2002). It is important to mention that the ELISA was performed using crude antigen prepared from DENV-infected C6/36 cells. The use of recombinant viral proteins as antigens would likely improve the specificity of the assay.
Another limitation of our study is that information on patient allergies was unavailable. A patient allergic to common allergens, that is, mosquito salivary proteins, could contain antibodies that cross-react with proteins in the crude antigen preparation. The availability of this information could assist in the development of an assay with improved specificity. The inclusion of DENV IgM and IgG ELISAs would have strengthened the study by allowing a direct comparison between our assay and traditional assays. The inclusion of IgE standards would have also strengthened the study. However, this is a proof-of-concept pilot study designed to test the feasibility of the idea to decide whether a larger study is warranted.
Our study has several additional limitations. Sera were not assayed for DENV IgG, and therefore, it is not known whether any healthy controls or non-dengue patients had previous DENV infections. Furthermore, dengue patients were not classified into those with primary and secondary DENV infections. Some studies have reported higher levels of DENV-specific IgE in patients with secondary infections (Abubakar et al, 1997; Vazquez et al, 2014; Vazquez et al, 2007), while others have reported no significant difference (Koraka et al, 2003). Another limitation is the lack of sera from patients with severe dengue. Higher levels of DENV-specific IgE have been detected in patients with dengue hemorrhagic fever compared with dengue fever (Bachal et al, 2015; Koraka et al, 2003).
DENV-specific IgE levels did not differ among patients when they were analyzed according to their age and sex. Others have also shown that DENV-specific IgE levels are not dependent on these demographic variables (Koraka et al, 2003). DENV-specific IgE levels also did not differ when patients were analyzed according to the DENV serotype responsible for the infection. To the best of our knowledge, the present study represents the first to compare DENV-specific IgE levels in patients infected with different serotypes. Additionally, we observed that DENV-specific IgE levels did not differ according to the time since illness onset. In contrast, DENV-specific IgE levels steadily increased with time in patients examined from 2 to 5 days after illness onset (Inokuchi et al, 2018). In another study, DENV-specific IgE levels were higher at 8–13 and ≥14 days compared with 1–7 days after illness onset (Koraka et al, 2003).
Conclusions
We performed a pilot study to evaluate whether IgE-based capture assays provide a suitable complementary assay to conventional assays for the early diagnosis of dengue. Our DENV IgE ELISA exhibited a high rate of sensitivity but was limited in its ability to differentiate between dengue and non-dengue febrile illnesses. Therefore, we conclude that IgE-based capture assays for the early diagnosis of dengue have potential, but further work is needed to reduce the high rate of false positivity.
Footnotes
Acknowledgments
We thank Rosa Cetina Trejo and Carlos Baak Baak at the arbovirology laboratory for their technical assistance on the ELISA test and providing the tools for the statistical analysis.
Authors' Contributions
This work was approved by all the co-authors. C.M.-W. and S.A. made significant contributions to conception and study design. Data and logistics for sample acquisition were completed by V.L.-T., A.R.D.-R., and G.C.R.-S. Data analysis and interpretation were performed by B.J.B., C.M.-W., S.A., and S.K. Also, C.M.-W., B.J.B., and S.A. have contributed to the writing the draft of the article and critically revised; whereas C.M.-W., S.K., and S.A. contributed with grant resources.
Author Disclosure Statement
The authors declare that there are not conflict of interests. I, Carlos Machain-Williams, state on behalf of my co-authors that the work described in this document is original research, it has not been previously published and is not considered for publication elsewhere, partially or completely. All authors read and approved the final work.
Funding Information
This work was supported in part by Bio & Medical Technology Development Program of the National Research Foundation (NRF) funded by the Korean government (MSIT) (no. 2016M3A9B8942289) (C.M.-W. and S.K.) and in part by the Ministry of Higher Education Malaysia, HICOE Program M0002-2019 (S.A.).