
Abstract
The ongoing COVID-19 pandemic poses a significant threat to human health. Many hypotheses regarding pathogenesis have been proposed and are being tried to be clarified by experimental and clinical studies. This study aimed to reveal the roles of the innate immune system modulator GAS6/sAXL pathway, endothelial dysfunction markers vascular endothelial growth factor (VEGF) and hypoxia-inducible factor (HIF)-1α, and antiviral effective TRIM25 and TRIM56 proteins in pathogenesis of COVID-19. The study included 55 patients with COVID-19 and 25 healthy individuals. The serum levels of GAS6, sAXL, VEGF, HIF-1α, TRIM25, and TRIM56 were measured using commercial ELISA kits and differences between COVID-19 patients and healthy controls, and the relationship to severity and prognosis were evaluated. GAS6, sAXL, TRIM56, and VEGF were found to be higher, while TRIM25 was lower in patients. There were strong positive correlations between GAS6, sAXL, TRIM25, TRIM56, and VEGF. None of the research parameters other than HIF-1α was associated with severity or prognosis. However, HIF-1α was positively correlated with APACHE II. We speculate that the antiviral effective TRIM25 and TRIM56 proteins, as well as the GAS6/sAXL pathway, act together as a defense mechanism in COVID-19. We hope that our study will contribute to further studies to elucidate the molecular mechanism associated with TRIM56, TRIM25, GAS6, sAXL, and VEGF in COVID-19 patients.
Introduction
Many mechanisms are blamed in the pathogenesis of SARS-CoV-2. The innate immune system plays an important role in the response of patients to viral infection (Bohn et al, 2020; Kasuga et al, 2021). Most human viruses, including SARS-CoV-2, try to avoid triggering the innate immune signal by developing a variety of mechanisms (Amara and Mercer, 2015; Amor et al, 2020; Hershko and Ciechanover, 1998; Morizono and Chen, 2014).
Tripartite motif-containing (TRIM) family proteins have E3 ubiquitin ligase activity and play a crucial role in apoptosis, pyroptosis, and innate immunity (Wan et al, 2021). TRIM proteins regulate inflammatory signaling pathways, mostly by targeting important viral components for destruction (Giraldo et al, 2020; Yang et al, 2016).
TRIM25 and TRIM56 proteins have been investigated in many virus infections (Chen et al, 2020a, Chen et al, 2020b; Hu et al, 2017; Liu et al, 2016, Liu et al, 2014; Rajsbaum et al, 2012). In particular, TRIM56 has been shown to have antiviral effects against both positive and negative single-stranded RNA viruses; Flaviviridae, Retroviridae, Coronaviridae, Paramyxoviridae, and Orthomyxoviridae, respectively (Liu et al, 2016, Liu et al, 2014; Tsuchida et al, 2010; Wang et al, 2011).
To our knowledge, there is no study present investigating the TRIM25 and TRIM56 levels in COVID-19 patients so far.
Tyro3, AXL, and Mertk, which are TAM receptors, play a role in numerous physiological and pathological processes, and their relationship with viral infections has been noteworthy in recent years (Nguyen et al, 2013; Rothlin et al 2015; Wang et al, 2020). It has been reported that AXL receptors act as entry receptor/cofactor for many enveloped viruses, such as filoviruses and flaviviruses, and regulate immune and inflammatory responses during viral infection (Wang et al, 2021a; Wang et al, 2021b).
Recently, it has been reported that AXL may increase in viral infection, thus being a potential target in the future (Wang et al, 2021a; Wang et al, 2021b). Growth arrest-specific 6 (GAS6) binds to all three TAM receptors and has the highest affinity for AXL receptor (Martín and García de Frutos, 2008). GAS6 is expressed in many tissues, including capillary endothelial cells, vascular smooth muscle cells, and bone marrow cells. GAS6/AXL pathway also modulates acute inflammation (Bellan et al, 2019; Morales et al, 2021; Recarte-Pelz et al, 2013).
Hypoxia is a primary pathophysiological feature and a major cause of death in severe COVID-19 patients and hypoxia-inducible factor-1α (HIF-1α) is the transcriptional regulator of metabolic processes, including metabolism, cell proliferation, and angiogenesis under hypoxic conditions (Chen et al, 2020a, Chen et al, 2020b; Serebrovska et al, 2020; Ziello et al, 2007). HIF-1α is a direct transcriptional repressor of interferon regulatory factors, which are transcriptional activators of type I interferons. Activation of HIF-1α results in activation of many pathological pathways (Hashimoto et al, 2015; Tian et al, 2021).
Vascular endothelial growth factor (VEGF), a potent inducer of vascular permeability, is involved in pathological processes such as cancer, stroke, cardiovascular diseases, as well as COVID-19-associated pulmonary edema, sepsis, and acute lung injury (Claesson-Welsh, 2015). Since SARS-CoV-2 inhibits its receptor angiotensin-converting enzyme-2 (ACE2), it causes upregulation of VEGF, thus leading to increased vascular permeability and exacerbation of endothelial damage. Besides, it has been reported that high levels of VEGF in SARS-CoV-2 infected lung tissue trigger plasma extravasation, leading to pulmonary edema (Jin et al, 2020).
In this study, we aimed to reveal the roles of GAS6/sAXL, VEGF, HIF-1α, and TRIM proteins in the pathogenesis of the disease in COVID-19 patients.
Materials and Methods
Study population
This study included RT PCR (+) COVID-19 patients admitted to Erciyes University Faculty of Medicine Hospital COVID-19 inpatient unit or COVID-19 intensive care unit between February 1 and May 1, 2021. Exclusion criteria were HIV-positivity, immune deficiency, and hematologic malignancy. The control group consisted of COVID-19 RT PCR (−) 25 healthy participants.
Biochemical measurement
Blood samples were collected from all patients within the first 24 h of admission to the inpatient or intensive care unit. After centrifuging at 2,000 g for 10 min, serum samples were aliquoted and kept at −80°C. On the analysis day, the levels of GAS6 (Bioassay Technology Laboratory;
Fifty microliters serial diluted standards and 50 μL streptavidin-HRP enzyme were added to the standard wells. Forty microliters serum samples, 10 μL antibody, and 50 μL streptavidin-HRP enzyme were added to each of the sample wells. Then, the plate was gently mixed and plate was covered with seal plate membrane and incubated for 1 h at 37°C. The plate was washed five times after incubation. Then 50 μL chromogen A and 50 μL chromogen B solutions were added to each well and incubated for 10 min in a light-free incubator for color changes. Finally, 50 μL stop solution was added to each well. We measured the optical density (OD) of each well under 450 nm wavelength within 10 min after having added stop solution. Nonlinear curve was obtained according to standard concentrations and GAS6, sAXL, TRIM25, TRIM56, VEGF, and HIF-1α serum protein concentrations were calculated from this curve.
Data collection
The following data of all patients at first admission were recorded: the heart rate (beats/min), mean arterial pressure (MAP, mm-Hg), respiratory rate (breaths/min), oxygen saturation %, body temperature (°C), Sequential Organ Failure Assessment (SOFA), and The Acute Physiology and Chronic Health Evaluation (APACHE) II scores.
In addition, the following data from laboratory parameters evaluated within the first 24 h were reviewed retrospectively: white blood cell (WBC), neutrophil, lymphocyte, platelet, procalcitonin, C-reactive protein (CRP), ferritin, D-dimer, fibrinogen, prothrombin time (PT), activated partial thromboplastin time (aPTT), and international normalized ratio (INR) values. Complete blood count (WBC, neutrophil, lymphocyte, platelet) on XN-9000 (Sysmex Co., Kobe, Japan) by impedance and flow cytometric method, CRP on Cobas c701 (Roche Diagnostics, Mannheim, Germany) by immunoturbidimetric method, procalcitonin, and ferritin on Cobas e801 (Roche Diagnostics) by electrochemiluminescence method, coagulation parameters (D-dimer, fibrinogen, PT, aPTT, INR) on CN 2000 (Sysmex Co.) by the coagulometric, and immunoturbidimetric methods were analyzed.
Statistical analysis
Statistical analysis was performed on SPSS 23.0 package program. The distribution of data was evaluated with the Shapiro-Wilk test. Continuous variables are presented as mean ± SD or median (25–75. quartiles), with and without a normal distribution, respectively. Categorical variables were given as n (%). Variables were compared with independent samples t-test or Mann–Whitney–U test according to distribution characteristics. Categorical parameters were compared with the chi-square test. The relationship between the parameters was evaluated with Spearman correlation analysis. A p-value <0.05 was considered statistically significant.
Results
We included 55 confirmed COVID-19 patients (26 females, 29 males) with a mean age of 57.25 ± 12.59 years and 25 healthy controls (12 females, 13 males) with a mean age of 55 ± 5.55 years. The age and gender distributions of the two groups were similar (p < 0.220 and p < 0.571, respectively). When we compared patient and control groups, we found that GAS6, sAXL, TRIM56, and VEGF levels were significantly higher, while TRIM25 levels were significantly lower in patients (Table 1 and Fig. 1).
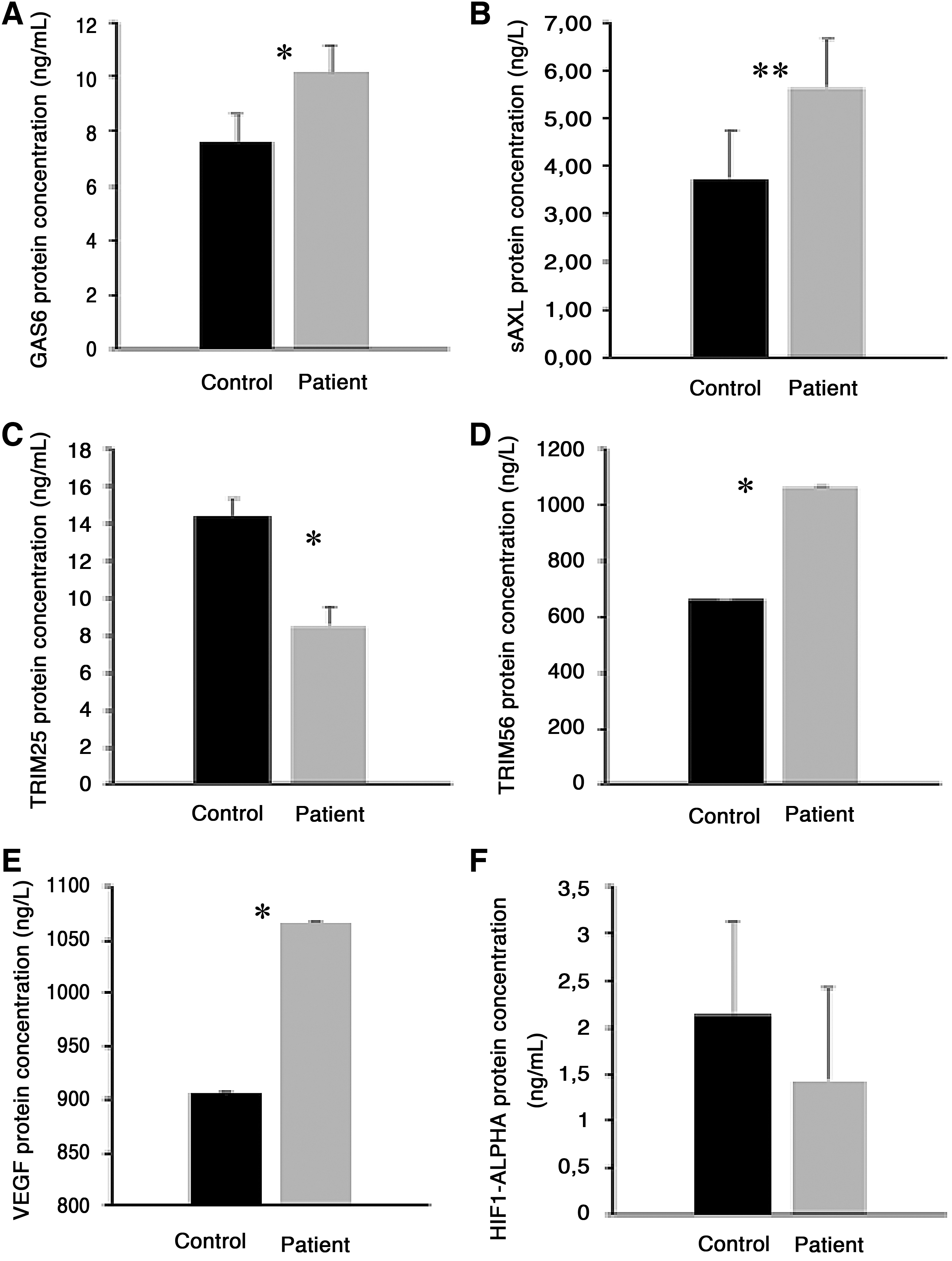
Bar diagram of GAS6, sAXL, TRIM25, TRIM56, VEGF, and HIF1-alpha in patient and control groups
Demographic Characteristics and Research Parameter Levels of Patient and Control Groups
Bold font indicates statistical significance.
GAS6, growth arrest-specific 6; sAXL, soluble AXL; TRIM, tripartite motif; VEGF, vascular endothelial growth factor; HIF, hypoxia-inducible factor
We divided the patient group into inpatient (n = 30) and intensive care unit (n = 25) subgroups and compared. While mean age of intensive care patients was significantly higher (p < 0.019), the gender distributions were similar (p < 0.104). We observed significant differences in clinical parameters with respiratory rate and APACHE II being higher, oxygen saturation being lower in the intensive care group. WBC, neutrophil, procalcitonin, CRP, ferritin, and D-dimer levels were significantly higher, while lymphocytes were significantly lower in intensive care unit patients but there was no significant difference in any of the research parameters between both groups (Table 2 and Fig. 2).
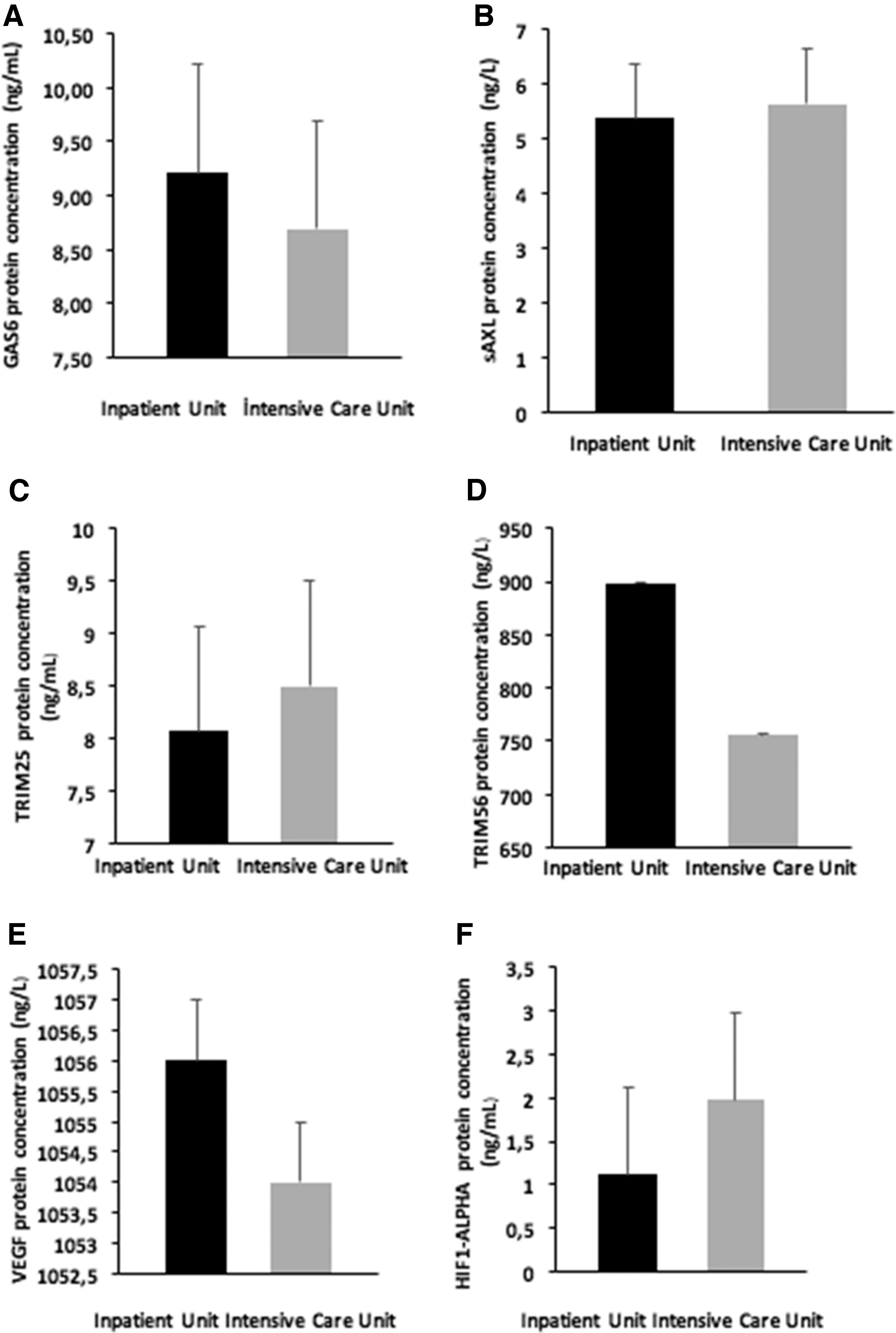
Bar diagram of GAS6, sAXL, TRIM25, TRIM56, VEGF, and HIF1-alpha in inpatient unit care and intensive care unit groups.
Demographic Characteristics, Routine, and Research Parameter Levels of Patients on Inpatient and Intensive Care Unit
Bold font indicates statistical significance.
MAP, mean arterial pressure; GKS, Glasgow coma scale; APACHE, The Acute Physiology and Chronic Health Evaluation; SOFA, Sequential Organ Failure Assessment; WBC, white blood cell; CRP, C-reactive protein; PT, prothrombin time; aPTT, activated partial thromboplastin time; INR, international normalized ratio.
All of the inpatient (6, nasal; 23, mask; 1, reservoir mask) and intensive care unit (3, mask; 2, reservoir mask; 10, noninvasive mechanical ventilation; 4, high flow; 6, invasive mechanical ventilation) patients received respiratory support. At the end of the 3-month follow-up, 12 COVID-19 patients died (inpatient unit, n = 2; intensive care unit, n = 10, p < 0.004).
We divided patients into survivor and nonsurvivor subgroups and compared demographic characteristics, levels of routine, and research parameters. The mean age of the nonsurvivors was higher than the survivors (p < 0.019). Heart, respiratory rate, APACHE II, and SOFA scores were higher in nonsurvivor patients. Besides, among the routine parameters, WBC, neutrophil, procalcitonin, ferritin, and D-dimer levels were higher. None of the research parameters was different between the nonsurvivors and survivors (Table 3).
Demographic Characteristics, Routine, and Research Parameter Levels of Patients with Survivors and Nonsurvivors
Bold font indicates statistical significance.
We performed correlation analysis to evaluate the relationship between research parameters in COVID-19 patients. There were strong positive correlations between GAS6, sAXL, TRIM25, TRIM56, and VEGF (Table 4).
Correlation Analysis Results in Patients
Bold font indicates statistical significance.
When the correlation of research parameters with APACHE II and SOFA scores was evaluated, we found a positive correlation only between HIF-1α and APACHE II (r = 0.287, p = 0.034) (Table 5). Besides, sAXL and TRIM56 levels were statistically increased in female patients compared to the controls, while only sAXL levels increased statistically in male patients compared to the control group (Table 6 and Fig. 3).

Bar diagram of sAXL and TRIM56, in male/female patient group.
Correlation of Research Parameters with APACHE II and SOFA Scores
Bold font indicates statistical significance.
Research Parameter Levels of Female/Male Patients and Control
Bold font indicates statistical significance.
Discussion
The ongoing COVID-19 pandemic poses a significant threat to human health, especially in terms of morbidity and mortality. Many hypotheses regarding pathogenesis have been proposed and are being tried to be clarified by experimental and clinical studies (Sharma et al, 2020). In the present study, we aimed to evaluate the molecular mechanism underlying the immune-inflammatory responses induced by SARS-CoV-2.
Recently, TRIM proteins have attracted attention for playing a critical role in host defense against viral infection (Gack et al, 2007; Uchil et al, 2008).
Although the TRIM family is largely cellular proteins, it is also found in the extracellular domain (Williams et al, 2019). For example, serum levels of TRIM72 were evaluated in colon cancer patients. However, the number of studies investigating these proteins at blood level is quite limited. In this study, we aimed to evaluate TRIM 25 and 56 in the serum of COVID-19 patients.
Recent studies draw attention to TRIM25 and TRIM56, which is known for its innate antiviral effects. Many studies have demonstrated the direct antiviral effect of TRIM25 andTRIM56 against a variety of viruses, including the Coronaviridae family (de Groot et al, 2013; Liu et al, 2016, Liu et al, 2014). Besides, molecular studies have shown that TRIM25 plays an antiviral role and that viruses follow various ways to antagonize TRIM25 activity (Martín-Vicente et al, 2017). Possibly, decreased TRIM25 levels may indicate insufficient antiviral defense in SARS-CoV-2, and we speculate that TRIM56 may be increased in COVID-19 patients to show its antiviral effects in physiological response due to its role in innate and adaptive immune mechanisms. However, TRIM25 and TRIM56, against a variety of viruses, and their mechanism of action have remained enigmatic.
As it is known, another possible mechanism involved in the innate immune system in the first defense against infections is the GAS6/AXL pathway. Therefore, at the present study, we also tried to investigate the role of GAS6/AXL pathway on the pathogenesis of COVID-19.
Increased levels of GAS6 and AXL have been reported in various spectrums of inflammatory states such as sepsis, septic shock, and systemic inflammatory response syndrome (Ekman et al, 2010; Stalder et al, 2016; Yeh et al, 2017). It has also been thought that therapeutic GAS6 replacement may be beneficial by reducing inflammation (Giangola et al, 2015). There are few studies evaluating GAS6 and AXL levels in SARS-CoV-2.
It has been earlier reported that GAS6 levels were increased in COVID-19 patients and GAS6 correlated with severity. The patients with the highest GAS6 levels had the lowest survival rate. In addition, sAXL was also higher in nonsurvivor patients than in survivors. They also proposed the idea that GAS6 and sAXL levels reflect the severity of COVID-19 (Morales et al, 2021).
In addition, it was reported that GAS6 and AXL levels were increased in intensive care unit patients with COVID-19 compared to healthy controls and were associated with poor clinical outcomes (Huckriede et al, 2021). Moreover, blood samples taken on day 12 revealed that GAS6 decreased over time in survivor patients.
Meanwhile, it is necessary to reemphasize the difference between cellular that is membrane-bound-AXL and sAXL. sAXL is formed by cleavage of cellular AXL extracellular domain with ADAM17. sAXL binds to the corresponding ligand in plasma before binding to cellular AXL and prevents binding to cellular AXL. This could be a host-developed defense system.
We thought and speculated that GAS6 and sAXL might be increased as a host defense system; in other words, GAS6 may have been increased in response to inducing inflammation, which is involved in pathogenesis of SARS-CoV-2. Accordingly, the defense mechanism of sAXL may have been to prevent related to the SARS-CoV-2 from binding to cellular AXL.
Activation and dysfunction of endothelial cells that express high levels of ACE2, which are key regulators of inflammation and coagulation, are thought to play a critical role in the pathogenesis of COVID-19 by altering vascular barrier integrity, promoting procoagulant status, and inducing endothelial inflammation (Del Turco et al, 2020; Jin et al, 2020).
In the present study, we also evaluated the role of VEGF and HIF-1α on the pathogenesis of COVID-19.
VEGF, located in the plasma, or in the blood-borne cells (Kut et al, 2007), is known to be a potent modulator of vascular biology and an important marker of endothelial dysfunction, it is upregulated by HIF-lα and is the primary cytokine involved in angiogenesis (Ylä-Herttuala et al, 2007). HIF-lα is a critical factor whose expression is induced in response to hypoxia at the site of inflammation and promotes phagocytic activity (Imtiyaz and Simon, 2010).
It was earlier reported that the level of VEGF was increased in COVID-19 patients and correlated with severity and prognosis (Rovas et al, 2021). Similarly, it was also shown that the bevacizumab, a VEGF inhibitor, may be beneficial in COVID-19 treatment (Pang et al, 2021).
HIF-1α and inflammatory cytokine expression were increased in COVID-19 patients (Tian et al, 2021), and similarly, it has been shown that elderly patients exhibited extreme inflammatory responses associated with mortality.
In the present study, we have shown that VEGF levels were increased in COVID-19 patients than in healthy controls, but there was no difference in HIF-1α levels. Possible reasons for increased VEGF may include microcirculatory disorders and hypoxia due to SARS-CoV-2 binding to ACE2-expressing endothelial cells.
However, the lack of elevation of HIF-lα suggested that VEGF might be triggered by another mechanism. ACE2 antagonizes VEGF-A to reduce vascular permeability during acute lung injury. Since SARS-CoV-2 downregulates ACE2, it likely cancels the VEGF-A-antagonizing effect of ACE2, leading to upregulation of VEGF and thus the increase of vascular permeability and aggravation of endothelial damage. Thus, cellular ACE2 reduction secondary to SARS-CoV-2 infection could be another possible reason of increased VEGF. In addition, VEGF is also thought to be the earliest marker for endothelial dysfunction, because VEGF levels may increase before active intravascular inflammation begins (Turkia, 2020). Increased VEGF levels indicate the presence of endothelial dysfunction and may play a role in the pathogenesis of endothelial damage in COVID-19.
Finally, there are many studies that have evaluated routine hematological and biochemical parameters in SARS-CoV-2. Increased leukocyte, neutrophil, PCT, CRP, ferritin, and D-Dimer and decreased lymphocyte are the common findings of almost all studies in COVID-19 patients, and it is well known that they are associated with severity and mortality (Henry et al, 2020; Izcovich et al, 2020). A multicenter prospective cohort study reported that APACHE II and age were predictive of mortality (Ferrando et al, 2020). A systematic review showed that tachycardia, tachypnea, and a high SOFA score were prognostic predictors. The findings of our study are quite compatible with studies up to now.
As a result, we speculate that the antiviral effective TRIM25 and TRIM56 proteins, as well as the GAS6/sAXL pathway, which plays an important role in inflammation and virus entry into the cell, act together as a defense mechanism in COVID-19 patients. Therefore, we thought that the GAS6/sAXL system treatment strategies may be promising in COVID-19. In addition, TRIM56, an antiviral peptide, increased as expected, but unexpectedly decreased TRIM25 levels may indicate that SARS-CoV-2 uses it as a virulence mechanism. The positive correlation between VEGF and GAS6, sAXL, and TRIMs can be considered as another possible mechanism, indicative of the repair of impaired endothelial function, one of the major challenges of COVID-19. According to this mechanism, GAS6, sAXL, and TRIM levels may have increased for the restoration of endothelial function.
Furthermore, we found that none of the research parameters other than HIF-1α was associated with the severity and prognosis of COVID-19, as previous studies have reported conflicting results for GAS6 and sAXL. Therefore, we still think that more studies are needed on this subject although as we have done with this study. Finally, we should emphasize that HIF-1α levels were positively correlated with APACHE II. Therefore, HIF-1α may have a crucial role in patients with severe endothelial dysfunction and hypoxia. We hope that our study will contribute to further studies to elucidate the molecular mechanism in COVID-19 patients.
Footnotes
Ethical Approval
Informed consent was obtained from all volunteers or, if unable to give consent, from the patients' relatives. This study was approved by Erciyes University Clinical Research Ethics Committee.
Authors' Contribution
G.B., A.B., A.K.A., S.M., P.S., and E.K. contributed to the idea and design of the study. G.B. drafted the initial article. A.B., A.K.A., S.M., P.S., and E.K. revised the article. M.O. and H.S. performed the presented analyses, had complete access to all the data, and take responsibility for the integrity and accuracy of the presented results. H.S., B.U., G.K.U., and S.O. contributed to data acquisition and interpretation. All authors reviewed and edited the article and approved the final version of the article.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by grants from the Fundamental Research Funds of Erciyes University (Project ID: THD-2021-11333) of Turkey.