
Abstract
Coronavirus disease 2019 caused by severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) initially emerged in December 2019 and has subsequently expanded globally, leading to the ongoing pandemic. The extensive spread of various SARS-CoV-2 variants possesses a serious public health threat. An extensive literature search along with deep analysis was performed to describe and evaluate the characteristics of SARS-CoV-2 variants of concern in relation to the effectiveness of the current vaccines and therapeutics. The obtained results showed that several significant mutations have evolved during the COVID-19 pandemic. The developed variants and their various structural mutations can compromise the effectiveness of several vaccines, escape the neutralizing antibodies, and limit the efficiency of available therapeutics. Furthermore, deep analysis of the available data enables the prediction of the future impact of virus mutations on the ongoing pandemic along with the selection of appropriate vaccines and therapeutics.
Introduction
Severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) is an enveloped virus with a positive-sense single-stranded RNA of ∼30 kilobases (Duan et al, 2020). The virus is made up of structural and nonstructural proteins, each is significantly contributing to the viral infectivity and transmission. The virus's structural proteins are the spike (S), envelope (E), membrane (M), and nucleocapsid (N) proteins (Durmaz et al, 2020; Mittal et al, 2020). Several variants of this virus have emerged during the last year and half (Chakraborty et al, 2022). Some of these variants are more transmissible, potentially more infectious, and can escape the neutralizing antibodies generated from previous infection or vaccination. These variants have been classified as variants of concern (VOCs) by the World Health Organization (WHO), and include Alpha (B.1.1.7), Beta (B.1.351), Gamma (P.1), Delta (B.1.617.2), and Omicron (B.1.1.529) (Petersen et al, 2022).
Alpha (B.1.1.7) variant originated in the United Kingdom, and is considered more infectious owing to the variant's high transmissibility (Chakraborty et al, 2022). Beta (B.1.351) variant was discovered in South Africa (Tang et al, 2021). Because of the E484K mutation in S protein, this variant possesses the ability of immune evasion and can avoid the neutralization by antibodies (Zhou et al, 2021). Gamma (P.1) variant was discovered in Brazil and is related to the reinfection in Manaus (Chakraborty et al, 2022). Delta (B.1.617.2) variant was first detected in India (Petersen et al, 2022). Omicron (B.1.1.529) variant was first discovered in South Africa and possesses the highest mutation rate among other VOCs (Petersen et al, 2022). Other significant variants include B.1.525 (U.K./Nigeria), P.2 (Brazil), P.3 (Philippines/Japan), and B.1.616 (France) (Chakraborty et al, 2022). A comprehensive analysis on the diversity of SARS-CoV-2 at the genomic level during the early second wave in Europe was also conducted (Miljanovic et al, 2021). Among 41 Serbian isolates, 148 different variants were detected. Seven mutations were very rare, while one was unique.
Spike-Y707H is an extremely rare substitution that was found for the first time in the European samples, before being detected in the United States (Deng et al, 2021). Mutation in ORF1ab, an essential gene for viral replication and immune evasion, constitutes a very rare variant identified in Brazil, United States, United Kingdom, Switzerland, and Serbia (Miljanovic et al, 2021). Furthermore, mutated ORF3a, an accessory protein located in between nsp1 and nsp16, constitutes a variant with rare mutations at Y109H, D183N, and C133S, which was also first detected in Europe (Bianchi et al, 2021; Miljanovic et al, 2021).
SARS-CoV-2 Structural Proteins and Their Role in Viral Infection
SARS-CoV-2 structural proteins (Fig. 1) play an important role in viral replication, attachment, and entry into the host cell in addition to spreading of infection (Satarker and Nampoothiri, 2020). S protein is a large surface glycoprotein (Satarker and Nampoothiri, 2020) that protrudes from the virion envelope giving a crown shape to the virus (Coutard et al, 2020). S protein recognizes and binds to the host cell receptors, providing a subsequent fusion between the viral envelope and host cell membrane (Satarker and Nampoothiri, 2020). S protein also plays a role in viral spreading since it facilitates the adhesion of the infected cells with adjacent uninfected cells (Schoeman and Fielding, 2019). S protein is a trimer that consists of an ectodomain, transmembrane part, and an intracellular C fragment (Satarker and Nampoothiri, 2020). The ectodomain is composed of two independent subunits; S1 that is responsible for receptor binding and S2 that enables membrane fusion (Satarker and Nampoothiri, 2020).
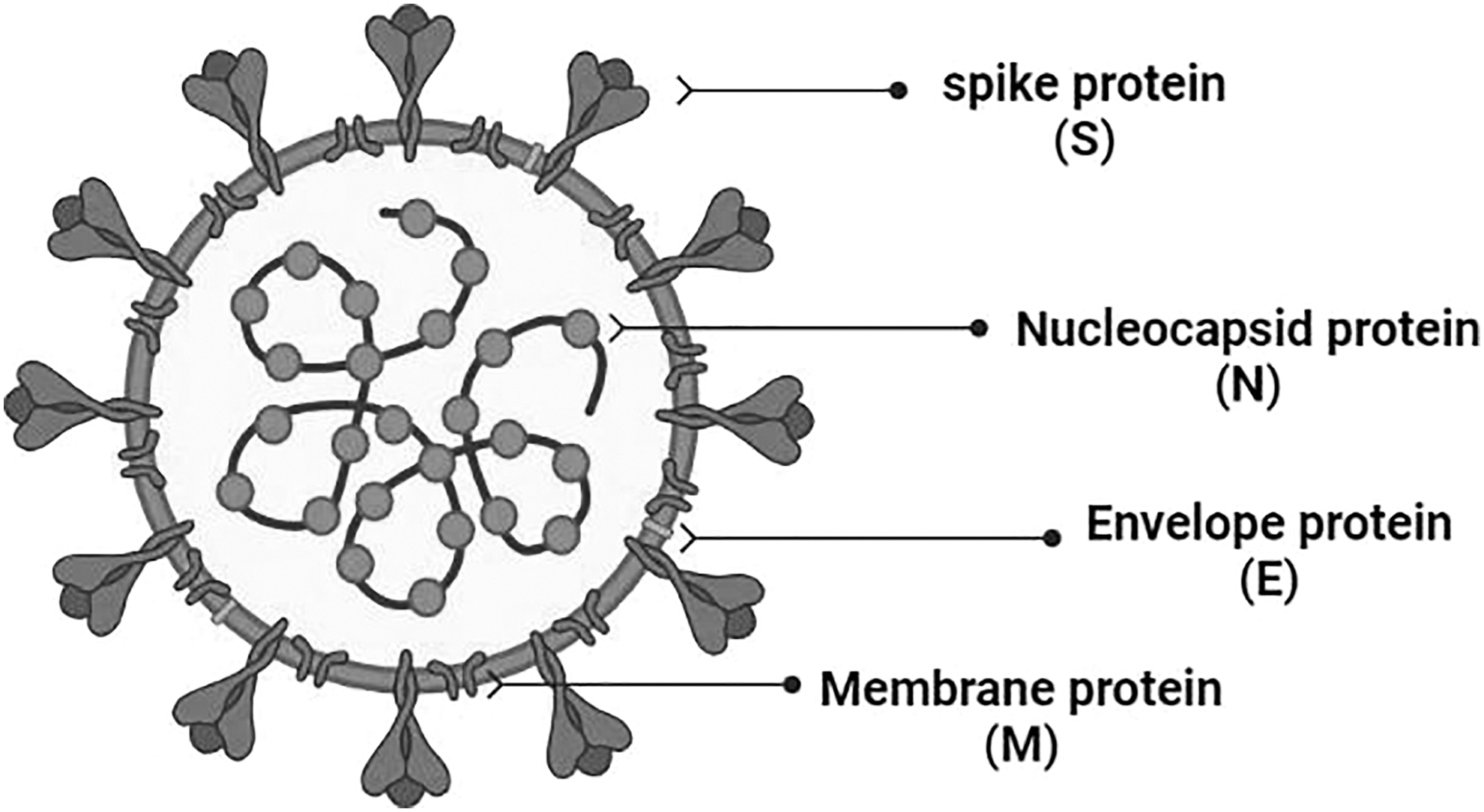
The different structural proteins of SARS-CoV-2 virus. SARS-CoV-2, severe acute respiratory syndrome coronavirus 2.
S protein undergoes conformational changes upon binding with angiotensin-converting enzyme 2 (ACE-2) receptor (Satarker and Nampoothiri, 2020). Upon binding with ACE-2, it changes to a stable postfusion state (Wrapp et al, 2020).
Nucleocapsid is the most abundant viral protein (Satarker and Nampoothiri, 2020). Its primary function is to encapsidate the viral genome (Surjit and Lal, 2008) to form the ribonucleoprotein (Satarker and Nampoothiri, 2020). Additionally, N protein has a role in (i) COX-2 activation and upregulation, which leads to lung inflammation (Satarker and Nampoothiri, 2020; Surjit and Lal, 2008), (ii) blocking the cell cycle by binding to cyclin–CDK complex that results in hypophosphorylation of retinoblastoma protein (Surjit and Lal, 2008), and (iii) inhibition of interferon production (Surjit and Lal, 2008). The envelope protein is a small integral protein that consists of an N-terminal domain (NTD), a hydrophobic domain, and a C terminal chain (Ruch and Machamer, 2011). Envelope protein forms ion channels called viroporins, which are essential for viral assembly and release (Satarker and Nampoothiri, 2020). It is also involved in pathogenic processes and can induce cell cytotoxicity (Ye and Hogue Brenda, 2007).
E protein may also disturb the epithelium of lungs (Teoh et al, 2010). The membrane protein, also known as matrix protein or E1 membrane glycoprotein (Hu et al, 2003), is one of the major membrane proteins in coronavirus (Lapps et al, 1987). It defines the shape of the viral envelope, forms ribonucleoproteins, and mediates inflammatory responses (Satarker and Nampoothiri, 2020). M protein is linked to the viral infectivity due to its binding potential to the viral S protein and host surface receptors to promote the membrane fusion (Hu et al, 2003). Other SARS-CoV-2 ORF, including nsp13, nsp14, nsp15, and orf6 are interferon antagonists (Yuen et al, 2020).
SARS-CoV-2 Structural Mutations
SARS-CoV-2 is a single-stranded RNA virus that possesses some level of a proof-reading activity (Yu et al, 2021), thus, it has no efficient RNA repairing mechanism (Sanches et al, 2021). The virus also spreads rapidly among a large number of susceptible hosts (Sanches et al, 2021). These two characteristics enable the virus to exhibit high genetic mutations, and consequently, generates new variants, including alpha, beta, gamma, and delta (Sanches et al, 2021). S protein is responsible for the majority of observed mutations in SARS-CoV-2 variants (Table 1) (Kannan et al, 2021). These mutations could affect the binding affinity to the target ACE-2 receptor or evade the neutralizing antibodies formed due to past infections or vaccination (Sanches et al, 2021). Consequently, the variants may exhibit more infectivity, transmissibility, and evasiveness (Joshi et al, 2021; Lauring and Hodcroft, 2021; Sanches et al, 2021).
S Protein Mutations in Each Variant (Sanches et al, 2021)
Major mutations in SARS-CoV-2 variants
Alpha variant mutations
SARS-CoV-2 Alpha (B.1.1.7) variant has 23 mutations (Mohammadi et al, 2021), including 6 mutations in the S protein (Meng et al, 2021). These mutations result in the alteration in the interaction with ACE-2 receptor; thus, increasing the infectivity, reducing the efficacy of neutralizing antibodies, and finally compromising the efficacy of the treatment (Mohammadi et al, 2021). The important mutations in S protein of this variant include E484K, S494P, and N501Y, which are located in the receptor binding domain (RBD), in addition to other mutations including 69del, 70del, D614G, 144del, A570D, S982A, P681H, D1118H, T716I, and K1191N (Chakraborty et al, 2022). The most significant mutations in this variant are H69-V70del, N501Y, and P681H, which are located in RBD, NTD, and close to the S1/S2 cleavage site (Muttineni et al, 2022).
Beta variant mutations
The Beta (B.1.351) variant has 12 mutations (Mohammadi et al, 2021), 77% of them occur in the S protein (L18F, D80A, D215G, LAL 242–244 Del, R246I, K417N, E484K, N501Y, D614G, and A701V) and the remaining mutations are present in ORF1a, E, and N viral proteins (Mohammadi et al, 2021). Most of these mutations are located within the immune-related regions such as RBD and NTD domains (Muttineni et al, 2022). K417N, E484K, and N501Y mutations are able to escape the neutralizing effect of monoclonal antibodies (mAbs) (Zhou et al, 2021), and decrease the vaccine efficacy (Chakraborty et al, 2022; Muttineni et al, 2022). This variant also shows stronger binding affinity to ACE-2 receptor (Ramanathan et al, 2021).
Gamma variant mutations
Gamma (P.1) variant has the highest number of mutations in S protein compared with the original Wuhan strain (Chakraborty et al, 2022; Muttineni et al, 2022). The mutations in the RBD region are K417T, E484K, and N501Y (Chakraborty et al, 2022). Other mutations in S protein include T20N, R190S, D614G, P26S, D138Y, H655Y, L18F, and T1027I (Chakraborty et al, 2022). The Gamma variant shows an increase in the resistance to neutralizing antibodies (Hirotsu and Omata, 2021). The most significant mutations in the genome sequence of this variant are E484K, K417T, and N501Y (Hirotsu and Omata, 2021).
Delta variant mutations
The Delta (B.1.617.2) variant was first detected in India and became the dominant variant worldwide (Fig. 2) (Ramesh et al, 2021). It is responsible for severe symptoms of the disease and increased risk of hospitalization. It is classified as VOC due to its rapid spread and severe symptoms (Ramesh et al, 2021). This variant has an advantage over the others because of the multiple mutations making it more transmissible (Ramesh et al, 2021). Delta variant S protein contains eight mutations (Planas et al, 2021), four of them are located in NTD (T19R, G142D, Δ156–157, and R158G), two (L452R and T478K) in the RBD, one (P681R) mutation is close to the furin cleavage site, and one (D950N) mutation in the S region (Planas et al, 2021). These mutations are responsible for the increased transmissibility and evading the immune system (Ramesh et al, 2021).
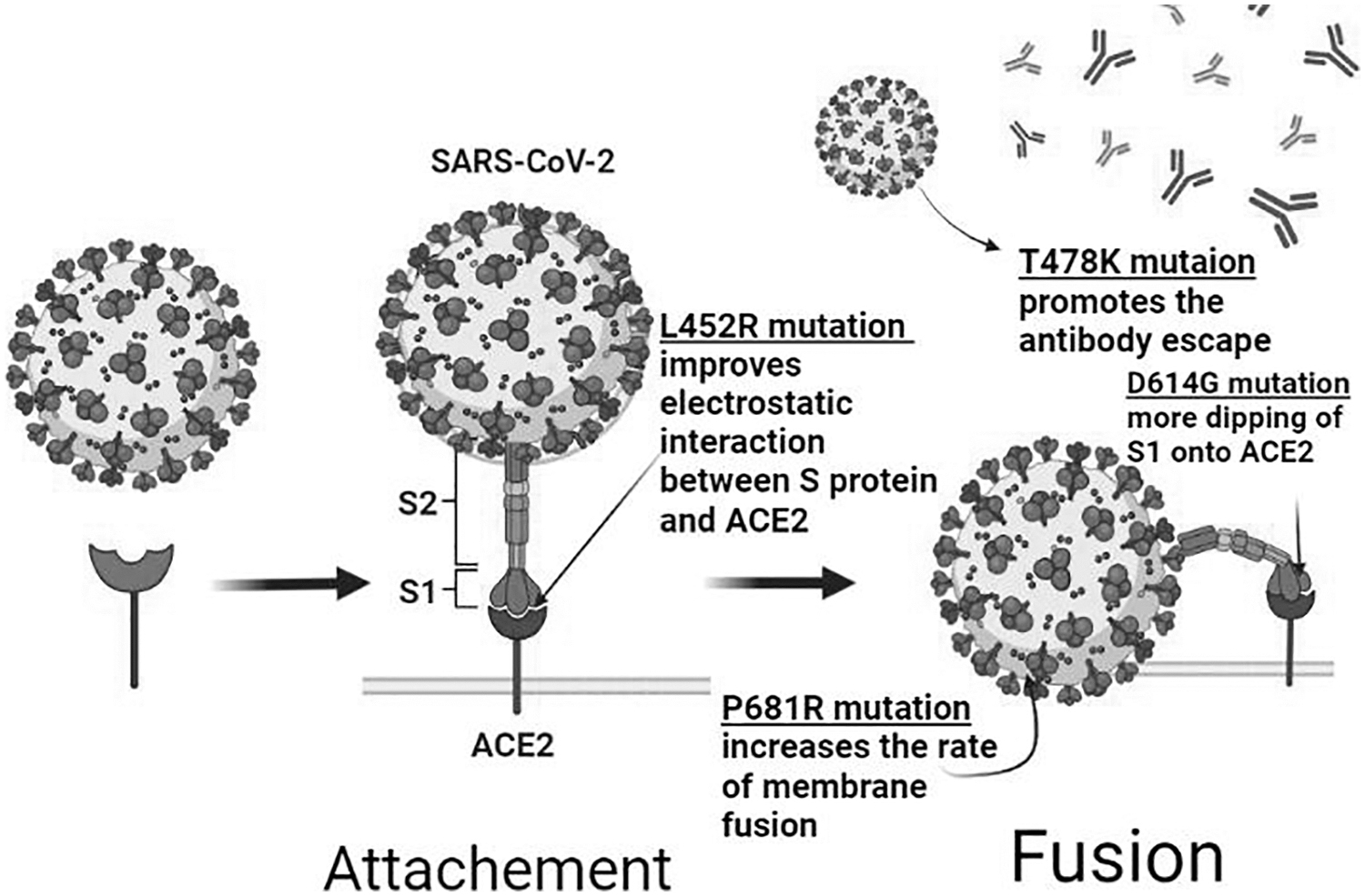
Structural mutations of the delta variant and their impact. D614G mutation in the S1 subunit results in improved virus–receptor binding and infectivity. L452R mutation in the RBD promotes the transmissibility by improving the virus–ACE-2interaction and reducing virus-neutralizing ability of antibodies. P681R mutation is adjacent to S1–S2 cleavage site, it increases the rate of membrane fusion, transmissibility, and infection. T478K mutation presents in RBD–ACE2 interface and is possibly related to immune escape. ACE-2, angiotensin-converting enzyme 2; RBD, receptor-binding domain.
Omicron variant mutations
The high transmissibility of Omicron (B.1.1.529) variant is due to more than 30 mutations in the S protein (Callaway, 2021). N501Y mutation enhances the binding affinity with ACE-2 receptor and when combined with Q498R, the binding affinity is further increased, leading to more accessibility to the host cells and an increased transmission (CDC COVID-19 Response Team, 2021). H655Y and N679K mutations are located near the furin cleavage site and can accelerate the cleavage of S protein, rendering the virus more infectious (Hossain et al, 2022). Numerous mutations in the RBD of S protein, such as Q493R, N501Y, S371L, S373P, S375F, Q498R, and T478K, are responsible for this increased affinity to ACE-2 receptor (Kumar et al, 2022; Wang and Cheng, 2021). Additionally, this variant produces a false negative result in polymerase chain reaction testing due to “S gene target failure,” leading to an undetected viral spread among the host (Torjesen, 2021). Furthermore, the likelihood of reinfection of previously SARS-CoV-2-infected individuals by Omicron is quite high (Pulliam et al, 2022).
Table 2 summarizes the most important viral variants and the significant and predominant mutations. Mutations displayed in one variant may appear in the other variants especially when these mutations provide an increased viral fitness (Gong et al, 2021). For example, N501Y mutation that is found in Alpha variant and provides an improved virus-ACE-2 binding, is also present in the Beta and Gamma variants. Similarly, E484K and K417N/T mutations were first detected in the Beta and Gamma variants, are also present in several other lineages (Gong et al, 2021).
Severe Acute Respiratory Syndrome Coronavirus 2 Variants and Their Specific Mutations
ACE-2, angiotensin-converting enzyme 2; RBD, receptor-binding domain; SARS-CoV-2, severe acute respiratory syndrome coronavirus 2.
Characteristics of the Major Mutations
D614G mutation
In this mutation, aspartic acid is replaced by glycine in the S1 domain (Korber et al, 2020). The overall effect is increasing the flexibility in the S1–S2 binding region because glycine is less bulky than aspartic acid, in addition to the loss of H-bond and salt bridge bindings (Joshi et al, 2021). Besides, this mutation increases the affinity of S1–S2 cleavage site toward the host transmembrane protease serine 2 (TMPRSS2) enzyme leading to more dipping of S1 domain onto the human ACE-2 (Joshi et al, 2021). These changes can decrease the stabilization of the locked conformation, thus, promoting the opening of RBD, and subsequently, improves the virus–receptor binding and the membrane fusion (Hu et al, 2020). It has been observed that D614G mutation increases S protein stability, thus, enhancing the overall fitness of SARS-CoV-2 (Hu et al, 2020).
L452R mutation
In L452R mutation, leucine, a hydrophobic amino acid, is substituted by arginine a hydrophilic amino acid, which abolishes the hydrophobic interactions (Joshi et al, 2021). This substitution generates a conformational alteration in the RBD (Joshi et al, 2021). This leads to a significant improvement in the electrostatic interaction between the S protein and ACE-2 (Deng et al, 2021), which is subsequently increasing the transmissibility of the virus (Joshi et al, 2021). Furthermore, it can cause an alteration in the binding of S protein with the neutralizing antibody, leading to a significant reduction in the virus-neutralizing ability of the antibodies (Muttineni et al, 2022).
P681R mutation
P681 residue is located near the S1–S2 furin cleavage site (Joshi et al, 2021). In P681R mutation, proline (P) is replaced by arginine (R) (Joshi et al, 2021). This might lead to an increase in the rate of membrane fusion and better transmissibility (Joshi et al, 2021). One study suggested that the increased coronavirus disease 2019 (COVID-19) severity and unusual symptoms caused by the delta variant are partly due to the increased fusogenicity that resulted from P681R mutation (Saito et al, 2022). Additionally, it was demonstrated that this mutation plays a critical role in the replication of delta variant (Liu et al, 2022). Furthermore, it may be responsible for the increased fitness of the delta variant (Liu et al, 2022). Detailed structural analysis showed that L452R and E484Q mutations along with P681R may result in an increased binding with the human ACE-2 and enhanced transmissibility (Joshi et al, 2021).
T478K mutation
T478K mutation is unique to the delta variant (Planas et al, 2021). Similar to E484K mutation that promotes antibody escape (Planas et al, 2021), the 478th residue is presented at the RBD–ACE-2interface (Xu et al, 2021). In this mutation, threonine is replaced by lysine (Joshi et al, 2021). Threonine is a polar uncharged amino acid, thus, the substitution by the positively charged amino acid lysine, results in a more positive surface with higher electrostatic potential of the S protein (Joshi et al, 2021). Experimental investigations demonstrate that T478K is also possibly engaged in escaping the immune recognition (Muecksch et al, 2021).
SARS-CoV-2 Variants and Major Vaccines
The immunological responses elicited by SARS-CoV-2 vaccinations and/or viral infections are critical in controlling the COVID-19 pandemic (Wu et al, 2020). The molecular interpretation of the antibody to S glycoprotein, stimulated through vaccination and/or prior viral infection, is identified as a key factor in explaining the efficacy of SARS-CoV-2 vaccines against the emergence of the new SARS-CoV-2 variants and their resistance to neutralization (Martínez-Flores et al, 2021). At least 590 neutralizing antibodies targeting the SARS-CoV-2 S glycoprotein are registered in the Protein Data Bank.
In many regions of the world, Omicron is currently the dominant variant, while the current vaccines that rely on the S protein of the original Wuhan virus is still in use (Hammerschmidt et al, 2021). These include mRNA vaccines (Pfizer-BioNTech BNT162b2 and Moderna mRNA-1273), viral vector vaccines (Oxford/AstraZeneca AZD1222 and J&J/Janssen's Ad26.COV2.S), and the recombinant S protein vaccine (Novavax NVX-CoV2373), in addition to the inactivated viral vaccines by Sinovac and Sinopharm BBIBP-CorV (Bian et al, 2021b). Since they are dependent on the wild-type virus (sequenced in January 2020), their efficacy against the new variants may be compromised to some extent.
BNT162b2 (Pfizer–BioNTech) mRNA vaccine
Pfizer and BioNTech collaborated on the development of BNT162b2, a mRNA SARS-CoV-2 vaccine (Teo, 2021) that consists of a modified RNA, encoding the full SARS-CoV-2 S protein, along with two proline mutations to secure the prefusion conformation (Wrapp et al, 2020). T4 fibritin-derived foldon trimerization domain added to facilitate the trimer formation, consequently potentiates the immune responses (Kaur and Gupta, 2020). Negatively charged mRNA is encapsulated into a positive lipid nanoparticle that acts as endocytosis facilitator and RNase evader (Li and Li, 2021; Reichmuth et al, 2016).
mRNA-1273 (Moderna TX, Incorporation)
Moderna and the Vaccine Research Center at the National Institute of Allergy and Infectious Diseases (NIAID) in the National Institutes of Health (NIH) collaborated on the development of mRNA-1273, a mRNA SARS-CoV-2 vaccine (Corbett et al, 2020). mRNA-1273, like BNT162b2, contains the sequence encoding the full S protein sequence with two proline mutations (Jackson et al, 2020; Kaur and Gupta, 2020). An alteration in the furin cleavage site (682-RRARS-686) was created to maintain a stable prefusion state of S protein. Arginine amino acid residues (R) are substituted by glutamine (Q), serine (S), or glycine (G) to hinder the cleavage process (Hoffmann et al, 2020a). This mediates the ability to trigger an antiviral response that is largely S-protein specific (Tu et al, 2020).
To keep the S protein in its stable prefusion state, a modification to the furin cleavage site (682-RRARS-686) was made. To slow down the cleavage process, glutamine, serine, or glycine are substituted for the arginine amino acid residues (R), which inhibits the cleavage (Hoffmann et al, 2020a). This mediates the ability to elicit an S-protein-specific antiviral response (Tu et al, 2020).
NVX-CoV2373 (NOVAVAX) protein subunit vaccine
Novavax Incorporate developed NVX-CoV2373, a SARS-CoV-2 protein subunit vaccine (Patel et al, 2021). On July 2022, the U.S. Food and Drug Administration (FDA) issued an Emergency Use Authorization (EUA) for the Novavax COVID-19 Vaccine. It is composed of recombinant S protein generated from SARS-CoV-2 in a prefusion state in a nanoparticle formulation (Patel et al, 2021). S protein is stably expressed using the Baculovirus Expression Vector System (Patel et al, 2021; Tang et al, 2020). NVX-COV2373 utilizes two proline substitutions at K986P/V987P, in addition to alteration in the furin cleavage site by replacing three arginine amino acids with glutamines (R682Q, R683Q, and R685Q) (Bangaru et al, 2020). The subunit vaccine offers weak immunogenicity and requires the inclusion of an adjuvant to boost the induced immune function (Cao et al, 2018).
It utilizes Matrix-M adjuvant, which is composed of Quillaja saponins coupled with cholesterol and lipids into nanoparticles to boost Th1 and Th2 responses. This generates various subclasses of antibodies to improve the immune cell migration and enables dose reduction (Kyriakidis et al, 2021; Madhun et al, 2009; Magnusson et al, 2018; Rajput et al, 2007). Adaptive immune responses, including helper T cells play a major role in disease control, including SARS-CoV infection; they activate the B cells to secrete sufficient antibodies and macrophages to combat microbial infections and to kill infected target cells (Moss, 2022).
ChAdOx1 nCoV-19 (Oxford-AstraZeneca) replication-defective viral vector vaccines
Oxford University and AstraZeneca collaborated on the development of ChAdOx1 (AZD1222), a viral vector vaccine (Sharma et al, 2020). ChAdOx1 consists of a replication-defective chimpanzee adenoviral vector ChAdOx1, encoding SARS-CoV-2 structural surface S protein, which triggers the immune responses (van Doremalen et al, 2020). The sequence utilizes a codon optimization for the S glycoprotein in the amino acids 2-1273 (Patel et al, 2021).
Ad26.COV2.S/JNJ-78436735 (Johnson & Johnson–Janssen vaccine)
Johnson & Johnson–Janssen collaborated on the development of Ad26.COV2.S.1, a single-dose nonreplicating adenoviral vector vaccine (Barouch et al, 2021). Ad26.COV2.S expresses a full-length SARS-CoV-2 S protein in a prefusion stabilized conformation using a human adenovirus type 26 (Ad26) viral vector (Abbink et al, 2007; Bos et al, 2020). To achieve the prefusion conformation, S protein employs two proline substitutions at K986P/V987P and employs an alteration in the furin cleavage site by introducing R682S and R685 amino acids (Bos et al, 2020).
BBIBP-CorV/CoronaVac (Sinopharm/Sinovac) inactivated viral vaccines
Sinopharm developed BBIBP-CorV, a two-dose SARS-CoV-2 inactivated viral vaccine (Alderson et al, 2021). Sinovac also developed CoronaVac, a two-dose SARS-CoV-2 inactivated viral vaccine (Alderson et al, 2021). BBIB-CorV is inactivated by propiolactone, whereas CoronaVac is inactivated by a combination of propiolactone and formaldehyde (Alderson et al, 2021). Both employ plaque-purified isolates produced in Vero cells (Alderson et al, 2021). Both vaccinations are given intramuscularly with aluminum hydroxide as an adjuvant (Alderson et al, 2021).
Evaluation and Enhancement of Vaccine Efficacy Against VOCs
Evaluation of the vaccine efficacy and effectiveness in relation to VOCs
A literature search was performed to investigate the vaccine effectiveness against symptomatic infection and severe disease or patient hospitalization by B.1.1.7, B.1.351, P.1, and B.1.617.2 variants. A Blobbogram/forest plot for each variant (Fig. 3A–E), constructed using Microsoft Excel, was utilized to pool several clinical trials of various study designs, including cohort, case–control, pivotal efficacy trial, randomized controlled trials (RCT), and test-negative design (Supplementary Table S1). To facilitate data location, “COVID-19,” “SARS-CoV-2,” “vaccine,” “variant,” “B.1.1.7/Alpha,” “B.1.351/Beta,” “P.1/Gamma,” “B.1.617.2/Delta,” and “B.1.1.529/Omicron” keywords were used to narrow the search. Data from nonmedical websites, non-English articles, and animals in vivo or in vitro trials were excluded. Studies involving individuals of age 16 and above from both genders were included.
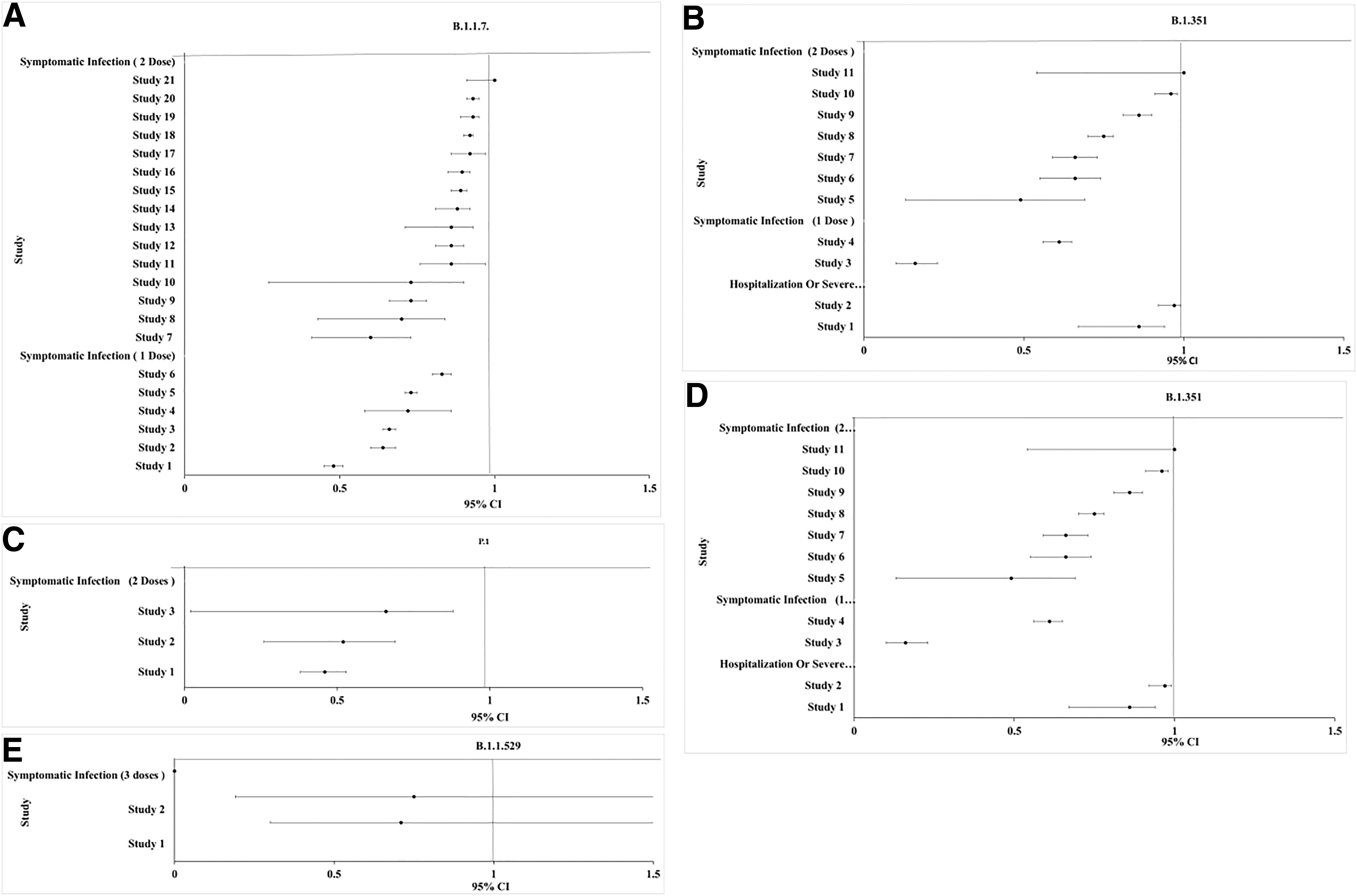
Blobbogram/forest plot pools of clinical trials studying the vaccine effectiveness against VOC. The Y-axis represents each study. The X-axis denotes each study according to the upper limit of confidence interval, effect size, and the lower limit of confidence interval.
Effectiveness of SARS-CoV-2 vaccines against Alpha variant
The effectiveness of SARS-CoV-2 vaccines against Alpha variant was evaluated based on 16 test-negative case–control (TNCC), 1 case–control, 2 RCT, and 2 cohort independent studies (Fig. 3A). This analysis included BNT162b2, mRNA-1273, ChAdOx1, and NVX-CoV2373 vaccines from clinically independent studies in several countries, including U.K., England, Scotland, Canada, France, and Qatar. In terms of symptomatic infection after two doses, mRNA-1273 appeared 100% (most effective) in a TNCC study. In another independent study, ChAdOx1 appeared 60% (least effective). In terms of symptomatic infection after one dose, the efficacy was less than two doses. The efficacy ranged from 88% in mRNA-1273 to 48.7% in BNT162b2/ChAdOx1.
Effectiveness of SARS-CoV-2 vaccines against Beta variant
The effectiveness of SARS-CoV-2 vaccines against Beta variant was evaluated based on two TNCC, six RCT, two cohort, and one pivotal independent study (Fig. 3B). This analysis included Ad26.COV2.S, BNT162b2, and mRNA-1273 vaccines from clinically independent studies across several countries, including USA, Qatar, France, and an International study. In terms of effectiveness against hospitalization and severe illness after two doses, BNT162b2 was 97.4% effective. In terms of symptomatic infection after two doses, BNT162b2 appeared 100% (most effective) in a pivotal study. In another independent study, BNT162b2 appeared as 49% (least effective). In terms of symptomatic infection after one dose, the efficacy was less than after two doses ranging from 61% in mRNA-1273 to 16% in BNT162b2.
Effectiveness of SARS-CoV-2 vaccines against Gamma variant
The effectiveness of SARS-CoV-2 vaccines against Gamma variant was evaluated based on one TNCC and two cohort independent studies (Fig. 3C). This analysis included mRNA-1273 and CoronaVac vaccines from clinically independent studies across countries, including Canada and Brazil. In terms of symptomatic infection after two doses, mRNA-1273 appeared as 66.2% (most effective) in a cohort study. In another independent study, CoronaVac appeared as 46.8% (least effective).
Effectiveness of SARS-CoV-2 vaccines against Delta variant
The effectiveness of SARS-CoV-2 vaccines against Delta variant was evaluated based on 16 TNCC, 7 cohort, and 2 RCT independent studies (Fig. 3D). This analysis included BNT162b2, Ad26.COV2.S, mRNA-1273, ChAdOx1, and inactivated vaccines from independent studies in China, U.K., Qatar, USA, and Canada. In terms of effectiveness against hospitalization after two doses, Ad26.COV2.S was 76.7% and 85.4% effectiveness at reducing hospitalization and severe illness after 14 and 28 days, respectively.
In terms of symptomatic infection after three doses, homologous regimen of BNT162b2 appeared 92.6% effective 14 days after a booster dose. Nevertheless, heterologous regimen of two ChAdOx1 and one BNT162b2 appeared 93.8% effective 14 days after a booster dose. In terms of symptomatic infection after two doses, BNT162b2 appeared 88% (most effective) in a TNCC study. In another independent study, BNT162b2 appeared as 41.8% (least effective). In terms of symptomatic infection after one dose, the efficacy was less than after two doses ranging from 67.7% in BNT162b2 to 13.8% in inactivated vaccines.
Effectiveness of SARS-CoV-2 vaccines against Omicron variant
The effectiveness of SARS-CoV-2 vaccines against Omicron variant was evaluated based on two TNCC independent studies (Fig. 3E). This analysis included ChAdOx1 and BNT162b2. In terms of symptomatic infection after three doses, heterologous regimen of two ChAdOx1 and one BNT162b2 appeared 75.5% in a TNCC study 14 days after a booster dose. Nevertheless, homologous regimen of BNT162b2 appeared 71.4% effective 21 days after a booster dose.
Enhancement of vaccine strategies against SARS-CoV-2 variants
The decrease in neutralization capacity and protective effectiveness of existing vaccinations suggest that VOC may escape the immune system to some extent (Bian et al, 2021a). Escaped SARS-CoV-2 variants might represent an unforeseen threat to the whole world, necessitating rapid global vaccine research and implementation measures (Bian et al, 2021a). As a result, FDA has announced that it is critical to update the evaluation of current vaccines against viral variants (Bian et al, 2021a).
Rapid and extensive vaccination campaigns using existing SARS-CoV-2 vaccines
According to the WHO, the effectiveness of SARS-CoV-2 vaccinations must meet a minimum threshold of 50% (Li et al, 2021). Moreover, in reference to the previous blobbograms against several VOCs, most of the vaccines can achieve the minimum threshold after a two-dose regimen against symptomatic infection. However, limited studies' findings suggest an effectiveness at lower than 50% threshold in Beta, Gamma, and Delta after a two-dose regimen. Nevertheless, the current greatest approach to manage the threat of the emerging variations still remains to accelerate the distribution of existing vaccinations to immunize the major segments of the global population (Bian et al, 2021a).
Approaches to enhance the immunogenicity of existing SARS-CoV-2 vaccines against VOC
Based on the aforementioned data in relation to the vaccination effectiveness against SARS-CoV-2 variants, increasing the level of vaccine-induced immune response might help to prevent the variant transmission (Bian et al, 2021a). Many approaches can be evaluated in this regard such as increasing the number of doses and mixing vaccinations (heterologous vaccination regimen) to enhance the immunological responses (Bian et al, 2021a). For instance, giving a third dose of BNT162b2 vaccine, whereby some countries are adopting this strategy on high-risk groups, including health care workers, could boost the immunity against VOC (Juno and Wheatley, 2021; Saiag et al, 2021). Furthermore, mixing ChAdOx1 and BNT162b2 stimulated a larger T cell response and was more effective against VOC compared with two doses of BNT162b2 (Pozzetto et al, 2021). For instance, in a real-world observational study, individuals using heterologous vaccination of ChAd/BNT are less likely to get infected in comparison to BNT/BNT.
The sera were examined for their capacity to inhibit VeroE6 cell infection by SARS-CoV-2 isolates (Pozzetto et al, 2021). Vaccinated individuals with ChAd/BNT showed serum-neutralizing antibody titers 2.3- to 3.6-fold greater than those immunized with BNT/BNT. These results suggest that S protein mutations in VOC have a less significant effect on the neutralizing capacity of antibodies produced by heterologous immunization than by homologous immunization (Pozzetto et al, 2021).
SARS-CoV-2 Variants and Major Therapeutics
Despite the availability of vaccinations, therapeutics with significant efficacy against SARS-CoV-2 virus are desperately needed; especially those effective against the current variants (Painter et al, 2021). In this study, we review the available therapeutics and their efficacy against the current variants. Different therapeutics and their efficacy against the variants are summarized in Supplementary Table S3. The impact of variant mutations on the drug efficacy is also summarized in Supplementary Table S4.
Effect of variants on the efficacy of S protein-targeted therapeutics
Since S protein is the site for the majority of mutations in the viral variants (Kannan et al, 2021), the efficacy of S protein-targeted therapeutics against the viral variants is expected to be influenced (Pokhrel et al, 2021). On the other hand, the efficacy of drugs that target the conserved regions of S protein should be largely maintained (Pokhrel et al, 2021).
mAbs against SARS-CoV-2 variants
Neutralizing antibodies are one of the most potent therapies that target the RBD of S protein (Walls et al, 2020; Wrapp et al, 2020) by inhibiting its attachment to ACE-2 receptors (Cerutti et al, 2021; Starr et al, 2021a). The RBD–ACE-2interface is a short sequence of 25 amino acids (Benton et al, 2020). Sequence mutations in this region, which is identified in all variants, can significantly cause immune escape (Benton et al, 2020). Various clinically approved mAbs and their efficacy on different variants are summarized in Figure 4.

Box-Whisker blot representing fold reduction in the neutralization efficacy of different mAbs against different variants in comparison to the reference strain.
Bamlanivimab (LY-CoV555) is a mAb that targets the RBD. It was granted an EUA by FDA (Starr et al, 2021b). Several in vitro studies showed its inability to neutralize Beta, Gamma, and Delta variants, despite its ability to inhibit the Alpha variant (Fig. 4A–D) (Group et al, 2021; Planas et al, 2021). Similarly, bamlanivimab was unable to neutralize the Omicron variant (Takashita et al, 2022) (Fig. 4E).
Q493R mutation (changes in the amino acid spatial structure) of the Omicron variant can cause the removal of hydrogen bonds or may cause the collision of antibody to CDRH3 region, which may explain the failure of neutralization by bamlanivimab and etesevimab (LY-CoV016) (Takashita et al, 2022). Etesevimab is another mAb that can be used in combination with bamlanivimab against the Omicron variant (Takashita et al, 2022). The loss of activity of bamlanivimab against the Delta variant is attributed to L452R, E484Q, and E484K mutations that are located at the S protein receptor-binding motif (RBM), the epitope recognized by bamlanivimab (Supplementary Table S4 and Fig. 6) (Hoffmann et al, 2021b; Starr et al, 2021b).
On the other hand, E484K mutation is responsible for the inactivity of the bamlanivimab against Beta and Gamma variants (Raman et al, 2021; Tao et al, 2021) (Supplementary Table S4 and Fig. 6). Moreover, the use of this drug was revoked later due to its failure in viral neutralization caused by the emergent variants (Stosor and Angarone, 2021).
Casirivimab–imdevimab is a cocktail of mAbs termed REGN-COV that was granted an EUA by the FDA (Aleem and Slenker, 2022). Despite its inability to neutralize the Omicron variant (Takashita et al, 2022), it shows significant neutralizing efficacy against Alpha, Beta, Gamma, and Delta variants (Fig. 4) (Planas et al, 2021; Tzou et al, 2020). Intravenous infusion of casirivimab and imdevimab to hospitalized COVID-19 patients significantly reduces the mortality in patients infected with those susceptible variants (Group et al, 2021).
Casirivimab–imdevimab combination along with sotrovimab, an authorized mAb, bind to a region at the core of RBD (Tao et al, 2021), which is more evolutionarily conserved than the highly evolving RBM; hence the mutations are less likely to affect their neutralizing efficiency (Stosor and Angarone, 2021). However, given the high transmission frequency of the Omicron variant, which accounts for more than 99% of SARS-CoV-2 cases in the United States, according to Centers for Disease Control and Prevention data, FDA has limited the use of casirivimab–imdevimab combination to patients infected with or exposed to variants that are susceptible to these therapies (Food and Drug Administration, 2022).
Bamlanivimab–etesevimab, a mAb combination (Starr et al, 2021b), was unable to neutralize Beta variant, in contrast to casirivimab–imdevimab (Hoffmann et al, 2021a). Additionally, patients infected with Alpha or Gamma variants were at lower risk of hospitalization when they received casirivimab–imdevimab combination compared with bamlanivimab-etesevimab combination (Falcone et al, 2021). Similarly, the Omicron variant was shown to be resistant to bamlanivimab-etesevimab combination (Takashita et al, 2022). Furthermore, because of the reduced susceptibility of most variants to bamlanivimab–etesevimab combination, FDA restricted its use only to areas where the combined frequency of variants is ≤5% (Kumar et al, 2021). Later, owing to the high prevalence of Omicron, for which the bamlanivimab–etesevimab is ineffective (Takashita et al, 2022), FDA has restricted its use to patients infected with or exposed to variants that are susceptible to these mAbs (Food and Drug Administration, 2022).
In fact, bamlanivimab–etesevimab combination binds to different but overlapping epitopes of RBD (Falcone et al, 2021), therefore, mutations conferring resistance to one of them would confer the resistance to the other (Stosor and Angarone, 2021). On the other hand, casirivimab–imdevimab combination binds to nonoverlapping epitopes on the RDB, hence, the likelihood of a single mutation to confer its effectiveness is low and consequently, they were shown to be effective against most VOC (Stosor and Angarone, 2021), despite their diminished efficacy against the Omicron variant (Li et al, 2022). Therefore, the emergence of antibody-resistant SARS-CoV-2 variants, which is restricting the efficacy of mAbs, can be mitigated by using a combination of antibodies that bind to distinct nonoverlapping epitopes (Weisblum et al, 2020).
The AstraZeneca (tixagevimab–cilgavimab) combination is human anti-SARS-CoV-2 recombinant mAb that bind to nonoverlapping epitopes of the RBD of S protein (Li et al, 2022). Tixagevimab (COV2-2196), cilgavimab (COV2-2130), and S309 (precursor of sotrovimab) were found to preserve their neutralizing efficacy against Beta, Gamma, and Omicron; however, the 50% Focus Reduction Neutralization (FRNT50) values of these mAb were 3.7 to 198.2 greater for Omicron than for Beta or Gamma (Takashita et al, 2022). Furthermore, the AstraZeneca (tixagevimab–cilgavimab) combination was found to inhibit Beta, Gamma, and Omicron variants; however, the FRNT50 values of this combination were greater for Omicron by a factor of 24.8 to 142.9 than for Beta or Gamma (Takashita et al, 2022).
Sotrovimab (VIR-7831) is a mAb targeting an epitope shared by both SARS-CoV-1 and SARS-CoV-2 (Pinto et al, 2020). It has been granted an EUA by FDA (Starr et al, 2021b). This neutralizing action of sotrovimab does not directly interfere with ACE-2 binding because it does not overlap with it on the RBD-binding interface (Tiecco et al, 2022). Sotrovimab shows a neutralizing activity against Delta variant and is expected to be active against Alpha, Beta, and Gamma variants (Fig. 4) (Planas et al, 2021; Tzou et al, 2020). Although sotrovimab exhibited less than twofold reduction in the neutralizing activity against Omicron, it is still defined as a “no change” criterion as specified by the FDA (the FDA fact sheet for sotrovimab denotes no change as a reduction of less than fivefold in susceptibility) (Cameroni et al, 2022). Therefore, sotrovimab appears to be potent against the Omicron variant (Cameroni et al, 2022).
VIR-7832 is another mAb, which is identical to sotrovimab, except for a three-amino acid GAALIE (G236A, A330L, I332E) change in the Fc domain (Cathcart et al, 2021). This enhances the affinity of sLeA-targeting antibodies, while reducing the binding to the inhibitory receptor, hFcγRIIB. Additionally, VIR-7832 shows an enhanced CD8+T cell response and demonstrates a high efficacy against Alpha, Beta, and Gamma variants (Kumar et al, 2021). It is expected to retain its neutralization activity against the Omicron variant according to preliminary data (Ferré et al, 2022).
Bebtelovimab (LY-CoV1404) is a recently discovered mAb that binds to an epitope within the RBD (Tiecco et al, 2022). As of February 11, 2022, it was granted an EUA by the FDA to be used for the treatment of adults and children with mild-to-moderate COVID-19 symptoms who are at high risk of progressing COVID-19 symptoms (Eli Lilly, 2020). Bebtelovimab can potentially neutralize the original SARS-CoV-2 virus and different variants, including Alpha, Beta, Gamma, and Omicron. The binding and neutralizing activity of bebtelovimab appear to be unaffected by the most prevalent mutations found in the known VOC, making it a suitable therapeutic agent for the treatment of the Omicron variant (Westendorf et al, 2022).
DXP-604, is a newly synthesized mAb (Tiecco et al, 2022). Preliminary studies indicate that the K417N single-site mutation in the Omicron variant RBD does not confer resistance to DXP-604 (Tiecco et al, 2022). However, the combination of K417N with several other mutations namely S477N, Q493R, G496S, Q498R, N501Y, and Y505H can reduce the binding affinity of DXP-604 to the Omicron RBD by nearly 30-fold compared with the original SARS-CoV-2 (Tiecco et al, 2022).
Despite the compromised effectiveness of several mAbs against SARS-CoV-2 variants (Planas et al, 2021), therapies targeting more conserved or structurally distinct epitopes of the S protein can be more effective against SARS-CoV-2 variants (Sun et al, 2021). Examples of such therapies can include nanobodies, soluble ACE-2 (sACE-2), and designed ankyrin repeat proteins (DARPins).
Nanobodies against SARS-CoV-2 variants
Nanobodies or variable heavy-chain domains are small recombinant single-domain antibodies (Zupancic et al, 2021). Owing to their small size, they can target SARS-CoV-2 epitopes cavities of the RBD (Zupancic et al, 2021). Nanobodies in multivalent forms can neutralize variants by binding to multiple nonoverlapping epitopes (Zupancic et al, 2021), and preventing the conformational changes required for viral entry (Zupancic et al, 2021). The nanobody 7A3 binds to a highly conserved region of RBD in its up and down conformation (Hong et al, 2021), and is effective against Beta and Delta variants (Hong et al, 2021). On the other hand, 8A2 nanobody, which interferes with the up conformation of the RBD, can neutralize SARS-CoV-2 variants to a lesser extent (Hong et al, 2021). Despite the decreased efficacy of 8A2 nanobody, a cocktail of 7A3 and 8A2 showed high affinity to RBD and enhanced activity against Beta and Delta variants (Hong et al, 2021).
Additionally, Nb22, an alpaca-derived nanobody, has 8.4-fold increased neutralization efficacy against Delta variant compared with Alpha variant, with an IC50 of 0.41 ng/mL (Wu et al, 2021). L452R, is a RBD mutation of Delta variant, which forms an additional hydrogen bond with Nb22 hydroxy group; thus, increasing the Delta variant-neutralizing activity (Supplementary Table S4 and Fig. 6) (Wu et al, 2021). Despite the effectiveness of nanobodies against SARS-CoV-2 variants, they are still under clinical trials (Hong et al, 2021).
sACE-2 against SARS-CoV-2 variants
sACE-2 is an engineered form of ACE-2 receptor that can neutralize the S protein of SARS-CoV-2 (Chan et al, 2020; Monteil et al, 2020). APN01, a recombinant human sACE-2 (Zoufaly et al, 2020), neutralizes the S protein of all variants with greater affinity when compared with original SARS-CoV-2 (Wirnsberger et al, 2022; Zoufaly et al, 2020). Furthermore, APN01 can neutralize different VOC with IC50 10 to 20 times higher than the original SARS-CoV-2 (Wirnsberger et al, 2022; Zoufaly et al, 2020). This is attributed to the increased infectivity of the variants (Wirnsberger et al, 2022) (Fig. 5). sACE-2-based therapies utilize the increased S protein-ACE-2 receptor binding affinity caused by the variant's mutations such as D614G to increase the S protein binding affinity to sACE-2 (Supplementary Table S4 and Fig. 5) (Monteil et al, 2020).
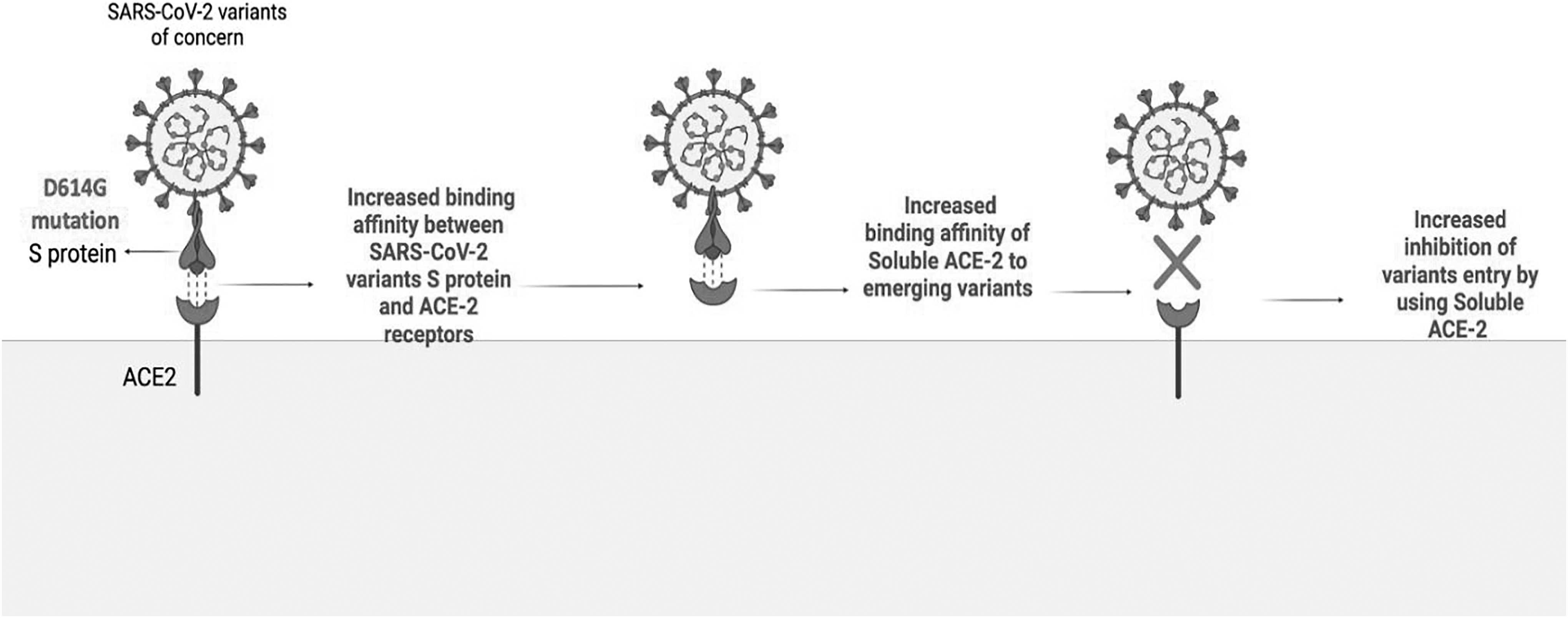
The mechanism of increased efficacy of soluble ACE-2 therapies against SARS-CoV-2 variants.
Since most mutations are more likely to increase rather than decrease the S protein-ACE-2 receptor-binding affinity (Barton et al, 2021), sACE-2-based therapies appeared as promising against the viral variants. Furthermore, the few mutations that decrease the S protein-ACE-2-binding affinity such as K417N is compensated by N501Y mutation (Raman et al, 2021); hence, the variants are less likely to hinder the effectiveness of sACE-2 (Supplementary Table S4) (Zoufaly et al, 2020).
DARPins against SARS-CoV-2 variants
DARPins are small protein scaffolds (Stumpp et al, 2008; Wirnsberger et al, 2022) that can be quickly designed against SARS-CoV-2 virus (Rothenberger et al, 2021; Wirnsberger et al, 2022). Multivalent DARPin is a group of connected DARPins, which unlike mAbs, can bind to different epitopes of the viral S protein; thus, minimizing the escape of the variants (Rothenberger et al, 2021; Wirnsberger et al, 2022). Ensovibep, is a multivalent DARPin having three domains that bind to the RBD (Walser et al, 2020). In vitro studies indicated that ensovibep is effective against most SARS-CoV-2 mutations and most emerging VOC (Rothenberger et al, 2021).
MP0423, is another antiviral DARPins that targets three different S protein domains namely RBD, NTD, and S2 domain (Rothenberger et al, 2021). It shows a declined efficacy against Alpha, Beta, and Gamma variants (Rothenberger et al, 2021). The reduced potency of MP0423 is attributed to D80A and R246I mutations of the Beta variant and P681H mutation of the Alpha and Delta variants at the NTD and S2 domains, respectively (Supplementary Table S4 and Fig. 6), which are the binding domains for MP0423 (Rothenberger et al, 2021).

Summary of different therapeutics employed against SARS-CoV-2 variants. It represents the SARS-CoV-2 variant therapeutics that act at different sites of the virus life cycle. Drugs highlighted with different colors indicating their efficacy against the viral variants. Grey-highlighted therapies have retained efficacy against the variants, light grey-highlighted therapies showed reduced efficacy and dark grey-highlighted therapies have lost their activity against the variants. Different mutations affecting the therapies are indicated.
On the other hand, F486V was the only mutation conferring resistance to ensovibep (Supplementary Table S4 and Fig. 6) (Rothenberger et al, 2021). However, F486V mutation is not very common among the spreading variants; thus, the efficacy of ensovibep against the viral variants is still maintained (Rothenberger et al, 2021).
Effect of variants on the efficacy of viral replication-targeted therapeutics
Proteins involved in viral replications such as RNA-dependent RNA polymerase (RdRp) enzyme and the viral main protease (Mpro or 3CL protease) are more evolutionarily conserved than the highly mutated S protein (Abdelnabi et al, 2021; Vangeel et al, 2022); hence, drugs targeting them are expected to be more effective against the emerging viral variants (Fig. 6) (Abdelnabi et al, 2021).
Molnupiravir against SARS-CoV-2 variants
Molnupiravir, is an oral prodrug with broad-spectrum antiviral activity (Fischer et al, 2022; Painter et al, 2021). It acts as a competitive nucleoside analog that targets the RdRp enzyme, increasing the frequency of the viral RNA mutations (Tiecco et al, 2022), thereby impairing the viral replication (Cannalire et al, 2022; Singh et al, 2021). FDA granted molnupiravir an EUA to COVID-19 patients who are at high risk and for whom other COVID-19 treatments are not available or clinically inappropriate (Vangeel et al, 2022). Recently, in phase 3 clinical trials, molnupiravir reduced the mortality and hospitalization by 50% (Singh et al, 2021). This efficacy was consistent against SARS-CoV-2 variants, including Gamma and Delta (Singh et al, 2021). Additionally, molnupiravir can reduce the viral loads of Alpha and Beta variants compared with the parent strain B.1-G (Fischer et al, 2022; Painter et al, 2021). Later, in vitro data proved the efficacy of molnupiravir against all VOCs, including the Omicron (Vangeel et al, 2022).
Remdesivir against SARS-CoV-2 variants
Remdesivir, is another nucleoside analog, and it is the first FDA-approved antiviral for the treatment of COVID-19 patients (Szemiel et al, 2021). Remdesivir retained its antiviral activity against all VOC, including Omicron, according to an in vitro study (Vangeel et al, 2022). Moreover, remdesivir equally inhibits the viral replication of Alpha and Beta variants compared with the original SARS-CoV-2 (Szemiel et al, 2021). Although several mutations in the RdRp are associated with remdesivir resistance namely (V557L, V473F, N491S, F480L/S/C, P323L, or E802D), none has been found in the Omicron variant (Tiecco et al, 2022). Additionally, there are no reports for the widespread presence of remdesivir-resistant mutations (Szemiel et al, 2021). This is indicating the effectiveness of remdesivir against the viral variants (Supplementary Table S4 and Fig. 6) (Szemiel et al, 2021).
Nirmatrelvir against SARS-CoV-2 variants
Mpro is a cysteine protease that cleaves the two SARS-CoV-2 polyproteins (pp1a and pp1ab) at several sites, forming several nonstructural proteins that are essential for viral replication (Tiecco et al, 2022). Nirmatrelvir (PF-07321332) is an irreversible inhibitor of SARS-CoV-2 Mpro that is coadministered with ritonavir to slow its metabolism, providing higher drug concentrations and enables an oral administration (marketed as Paxlovid) (Tiecco et al, 2022). When treatment is started during the first few days of symptom manifestation, it results in nearly a 90% reduction in the risk of severe COVID-19 and hospitalization (Vangeel et al, 2022). Although evolutionary changes might have a little impact on the Mpro gene, nirmatrelvir retained its antiviral effectiveness against all VOC, including the Omicron, according to an in vitro study (Vangeel et al, 2022).
The fact that several antiviral drugs, namely molnupiravir, remdesivir, and nirmatrelvir retained their efficacy against many SARS-CoV-2 variants is consistent with the high conservation of these drugs' target proteins (Tiecco et al, 2022; Vangeel et al, 2022). For instance, the RdRp when compared with the original lineage (NC 045512) has just two amino acid modifications (P323L in all VOC and G671S in Delta variant) (Vangeel et al, 2022). Additionally, since they are far from the active site of remdesivir and molnupiravir, they are expected to be effective against those variants (Vangeel et al, 2022). On the other hand, Mpro has two amino acid alterations namely K90R in Beta and P132H in Omicron (Vangeel et al, 2022). These mutations, like the RdRp, are not near the active region of the Mpro, hence, nirmatrelvir efficacy is not expected to be reduced against different VOC (Vangeel et al, 2022).
It is important to mention that at the global population level, the use of antivirals targeting Mpro and RdRp is very limited in comparison to the widespread neutralization antibodies generated through vaccination or postinfection. Thus, widespread use or misuse of these antivirals can in theory create a selective pressure that selects for mutations in Mpro or RdRp making them resistant to the antiviral action of small-molecule inhibitors. Thus, it is important to closely monitor the use of these antivirals and maintain genomic surveillance to preserve their efficiencies.
ATV006 against SARS-CoV-2 variants
ATV006, is a RdRp inhibitor at the preclinical stage that shows significant antiviral effectiveness against Delta variant by reducing the viral loads, lung damage, and mortality in animal studies (Cao et al, 2021). Therefore, ATV006 has a potential as an oral medication candidate against SARS-CoV-2 variants (Cao et al, 2021).
Prophylactic antiviral CRISPR in human cells against SARS-CoV-2 variants
Prophylactic antiviral CRISPR in human cells (PAC-MAN) is a CRISPR-based strategy targeting and inhibiting the viral RdRp conserved regions (Abbott et al, 2020). It demonstrated a robust inhibition of viral replication in human lung cells (Abbott et al, 2020), and is a beneficial strategy in combating the existing and emerging variants (Abbott et al, 2020). However, the lack of safe and effective in vivo delivery methods is the biggest barrier for its clinical development (Abbott et al, 2020).
Effect of variants on the efficacy of host protein-targeted therapeutics
As the host genes have lower mutation rate, therapies targeting host proteins that directly interact with the SARS-CoV-2 proteins can be effective against different variants (Fig. 6) (Egloff et al, 2004; Reuschl et al, 2021).
TMPRSS2 inhibitors against SARS-CoV-2 variants
TMPRSS2 inhibitors prevent the cleavage of SARS-CoV-2 S protein; thus, inhibiting its S2-mediated membrane fusion (Hoffmann et al, 2020b; Hu et al, 2021; Lee et al, 2021). Camostat and nafamostat are in phase 2 and 3 clinical trials (Lee et al, 2021). Camostat and nafamostat, in addition to bromhexine and aprotinin, were equally effective against Alpha and Beta variants compared with the reference lineage A in cell-based studies (Lee et al, 2021). Similarly, Omicron variant is sensitive to several TMPRSS2 inhibitors, including camostat, nafamostat, and aprotinin, according to preliminary data (Tiecco et al, 2022).
Plitidepsin against SARS-CoV-2 variants
Plitidepsin is an inhibitor of eukaryotic translation factor eIF4H that interacts with SARS-CoV-2 Nsp9 (nonstructural protein 9) (Egloff et al, 2004). It is in phase 2 clinical trials for COVID-19 management (Egloff et al, 2004). Nsp9 is involved in RNA synthesis and viral replication (Egloff et al, 2004). Plitidepsin potently suppressed the viral replication of both Alpha variant and the original SARS-CoV-2 equally in cell line-based analysis (Reuschl et al, 2021).
CD147 inhibitors against SARS-CoV-2 variants
CD147 is a receptor for SARS-CoV-2 and its variants (Geng et al, 2021). Mepolizumab is a humanized antibody that blocks the CD147 receptor and can inhibit the entry of SARS-CoV-2 variants, including Alpha, Beta, Gamma, and Delta in cell line-based studies with inhibition rates of 68.7%, 75.7%, 52.1%, 52.1%, and 62.3%, respectively (Geng et al, 2021). It can also reduce the inflammatory cytokine storm caused by SARS-CoV-2 and its variants (Geng et al, 2021). Infection of SARS-CoV-2 through CD147 can initiate the JAK-STAT pathway, which in turn induced the expression of cyclophilin A (CyPA). CyPA triggers the MAPK pathway, hence, regulates the expression of cytokines and promoted the development of cytokine storm (Geng et al, 2021).
Summary of the Proposed Effective Therapies Against Omicron
Although many single mAbs have either lost (bamlanivimab, etesevimab, and imdevimab) or significantly reduced (casirivimab, DXP-604) their neutralization activity against the Omicron variant, there are different therapeutic mAbs that can still be effective against the Omicron variant. For instance, sotrovimab, VIR-7832, and bebtelovimab retained their anti-Omicron activity. Moreover, tixagevimab and cilgavimab can maintain potential activity against Omicron variant as indicated earlier. Many combinations of mAbs, such as bamlanivimab–etesevimab and casirivimab–imdevimab have lost their neutralization efficacy against the Omicron variant, while the AstraZeneca tixagevimab–cilgavimab combination retained its activity (but with a reduced efficacy) against the Omicron variant. Unlike the mAbs, most antiviral drugs, which could inhibit the replication of Omicron, were effective against the Omicron variant, including molnupiravir, remdesivir, and nirmatrelvir.
Furthermore, different TMPRSS2 inhibitors namely camostat, nafamostat, and aprotinin can also be a potential therapeutic option against the Omicron variant, according to preliminary data. These results suggest that if further VOC emerge, they are likely to remain susceptible to existing (and presumably future) antivirals that are not targeting the S protein (Vangeel et al, 2022).
Emerging Variants and the Dynamic Pandemic Landscape
During the time of preparing this article, several new variants that have grabbed scientific and media attention have emerged. Among the several lineages of the Omicron variants, Omicron BA.2 (B.1.1.529.2) variant has received most attention. BA.2 was first reported by WHO from South Africa on November 24, 2021 (WHO, 2021). BA.2 shares 32 mutations with BA.1 but have 19 signature mutations of which 7 are found in the S protein (Majumdar and Sarkar, 2022). As of today, this variant has become dominant in the United States and across many countries in the world (CDC, 2022; Chen and Wei, 2022). Due to its recent emergence, there is little data on the transmission and antibody escape of BA.2. However, preliminary data suggest that BA.2 variant can be better at transmission and vaccine escape than BA.1 (Callaway, 2022; Chen and Wei, 2022; Rahimi and Talebi Bezmin Abadi, 2022). In addition to mutations due to RdRp infidelity, recombination between variants have also been documented.
A combination of Delta (AY.4) and Omicron (BA.1) variants named BA.1 × AY.4 recombinant or “Deltacron” was first detected in France in January 2022 (Mahase, 2022). Another recombinant variant that has received attention is the BA.1/BA.2 recombinant variant (XE), which was first detected in the United Kingdom on January 19, 2022 (Authority, 2022). Data on the transmissibility and vaccine escape of BA.1/BA.2 recombinant variant (XE) remain unavailable due to its recent emergence. The constant emergence of variants through mutation or recombination are a testament to the adaptability of this virus and the importance of genomic surveillance. Recently, sublineages, Omicron BA.4 and BA.5, are dominating, which are closely related to BA.2, while they contain further mutations in the RBD (Tuekprakhon et al, 2022).
Prediction of Future Variants
There is too much uncertainty about the probability and frequency of emergence of new variants (Murray and Piot, 2021). However, it is predicted that future SARS-CoV-2 mutations may enhance the viral infectivity and transmissibility (García de Alcañíz et al, 2021) as the virus quickly adapts and evolves (García de Alcañíz et al, 2021). A quick literature look can indicate that new mutant strains are emerging quickly and that they are more infective than the ancestral strains (García de Alcañíz et al, 2021). Factors that expose this virus to mutations and variants' development can include (i) lack of exonuclease proofreading activity, (ii) human immune system intervention, and (iii) rapid global spread and transmission, which give the virus a chance for the natural selection of rare-acted but favorable mutations (Chen et al, 2020).
Furthermore, the relatively high mutation rate in the S protein specifically in the RBD can lead to the development of several other variants (Chen et al, 2020). A study stated that most of the new mutants will disappear soon and only few will succeed to stay for a long period (García de Alcañíz et al, 2021). The failure or success of a mutant highly depends on the natural selection (García de Alcañíz et al, 2021). For a mutant to rise, it needs to have a selective advantage as in the case of S477N, N439K, and N501Y variants (García de Alcañíz et al, 2021).
The Top Net Tree method, which is based on algebraic topology and deep learning, was used to predict the binding free energy (BFE) changes generated by mutations in the S protein and the host ACE-2 receptor (Chen et al, 2020). Findings showed that mutations have rendered all SARS-CoV-2 variants more virulent than the original virus discovered in Wuhan (Chen et al, 2020). Furthermore, the BFE changes in all 3,686 probable future mutations were systematically estimated, revealing that the majority of mutations will improve SARS-CoV infectivity even more (Chen et al, 2020). According to sequence alignment, probability calculation, and BFE analysis, residues 452, 489, 500, 501, and 505 on the RBM have a high chance of mutations to more infectious strains of SARS-CoV-2 (Chen et al, 2020). The RBM Y495N mutation was said to have the biggest free energy change; thus, it may significantly improve the infectivity of this virus (Chen et al, 2020).
Other similar possible mutations are Y423C, Y423F, and Y423S in the residue 423 (Chen et al, 2020). It is also worth noting that residue 489 is a likely hotspot, since 5 mutations, including Y489H, Y489D, Y489F, Y489C, and Y489N, are predicted to improve the S protein's capacity to bind to the ACE-2 receptor (Chen et al, 2020). Another method for prediction is SpikePro, a simple computational model that predicts the impact of S protein mutations on the future of SARS-CoV-2 fitness, infectivity, and escape from the host immune system (Almehdi et al, 2021; Pucci and Rooman, 2021). SpikePro is a genomic surveillance program that predicts the development of new variants particularly in relation to the selective pressure generated by vaccination procedure on this virus (Burioni and Topol, 2021; Pucci and Rooman, 2021).
Conclusion
The future of the current COVID-19 pandemic remains largely uncertain. Continuous surveillance for the emergence of hazardous variants, as well as extensive study concerning their impact on the public health are essential. Because of the damaging effect of the current COVID-19 pandemic on public health, business, and society, vaccine development has become a top priority for the health care system. Although emerging SARS-CoV-2 variants can escape vaccine-induced immunity, novel approaches are being developed to take maximum advantage of the current vaccines in reducing morbidity and mortality. Additional exploration and analysis are needed to analyze the emerging VOCs, the effectiveness of vaccinations, the potency of potential therapeutics to aid in minimizing the hospitalization, morbidity, and mortality.
Footnotes
Authors' Contributions
All authors contributed equally.
Data Availability Statement
The data presented in the article have been made available.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This research has been funded by Sandooq Al-Watan and University of Sharjah to Sameh S.M. Soliman under grant #133006 and by the University of Sharjah to Mohamad Hamad under grant #CoV19-0306.
Supplementary Material
Supplementary Table S1
Supplementary Table S2
Supplementary Table S3
Supplementary Table S4
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.