
Abstract
Every year, dengue is responsible for 400 million infections worldwide. Inflammation is related to the development of severe forms of dengue. Neutrophils are a heterogeneous cell population with a key role in the immune response. During viral infection, neutrophils are mainly recruited to the infection site; however, their excessive activation is linked to deleterious results. During dengue infection, neutrophils are involved in the pathogenesis through neutrophils extracellular traps production, tumor necrosis factor-alpha, and interleukin-8 secretion. However, other molecules regulate the neutrophil role during viral infection. TREM-1 is expressed on neutrophils and its activation is related to increased production of inflammatory mediators. CD10 is expressed on mature neutrophils and has been associated with the regulation of neutrophil migration and immunosuppression. However, the role of both molecules during viral infection is limited, particularly during dengue infection. Here, we report for the first time that DENV-2 can significantly increase TREM-1 and CD10 expression as well as sTREM-1 production in cultured human neutrophils. Furthermore, we observed that treatment with granulocyte-macrophage colony stimulating factor, a molecule mostly produced in severe cases of dengue, is capable of inducing the overexpression of TREM-1 and CD10 on human neutrophils. These results suggest the participation of neutrophil CD10 and TREM-1 in the pathogenesis of dengue infection.
Introduction
Dengue is a viral disease caused by four antigenically related serotypes (Alejandria, 2009; Altay et al, 2016; Amrun et al, 2020; Banerjee et al, 2017) of the dengue virus (DENV). The incidence of DENV has recently increased by the mosquito vector spreading (Guzman et al, 2016). DENV is a positive-sense single-stranded RNA virus that belongs to the Flaviviridae family and is transmitted to humans by mosquitoes of the genus Aedes (Ferreira-de-Lima and Lima-Camara, 2018). The genomic RNA (11,000 nt) is translated to give rise to a large and single polyprotein precursor of structural protein C, PrM, and E and nonstructural proteins NS1, NS2a, NS2b, NS3, NS4a, NS4b, and NS5. It is estimated that 400 million dengue infections occur annually worldwide and the mortality rate surpasses 5–20% in some areas (Gubler, 2002; Murray et al, 2013).
Dengue infection can be asymptomatic; however, also can induce dengue fever (DF), dengue hemorrhagic fever (DHF), or dengue shock syndrome (DSS). DF is a severe flu-like infection where normally the patients usually recover without sequelae (World Health Organization, 2009). Moreover, DHF and DSS are associated with severe manifestations and death (Alejandra, 2009). Generally, severe forms of dengue occur when circulating levels of DENV decrease, and many authors suggest a correlation with high inflammation (Bozza et al, 2008; Espada-Murao and Morita, 2011). However, how this dysregulation happens is not fully understood.
Neutrophils are cells that normally can express CD15, CD16, CD66b, CD11b, and CD10, and fulfill diverse functions in the immune system (Blanco-Camarillo et al, 2021). During viral infection, the neutrophils restrict viral replication; however, its excessive activation could have a deleterious effect (Rawat et al, 2021). Importantly, during DENV infection, an increase in CD66b, neutrophils extracellular traps formation, interleukin (IL)-8, and tumor necrosis factor-alpha (TNF-α) have been reported (Banerjee et al, 2017; Opasawatchai et al, 2018). In the regulation of neutrophil activation, several cell surface molecules have been implied like TREM-1 (Triggering Receptor Expressed in Myeloid cells-1) and CD10 (Brandau and Hartl, 2017; Colonna and Facchetti, 2003).
Activation of TREM-1 on neutrophils amplifies the production of proinflammatory mediators (Bouchon et al, 2000). Soluble TREM-1 (sTREM-1) is produced in plasma as a result of inflammatory processes due to proteolytic cleavage of the TREM-1 membrane (Gomez-Pina et al, 2007). In turn, TREM-1 expression can be upregulated by invading pathogens or self-molecules, and differentially affected by cytokines (Bleharski et al, 2003; Bouchon et al, 2000; Wu et al, 2012). Nonetheless, there is still scarce knowledge about the participation of TREM-1 during viral infection (da Silva-Neto et al, 2021; Denner et al, 2013; Mohamadzadeh et al, 2006; Suthar et al, 2013).
CD10 is a 100-kDa transmembrane glycoprotein expressed on human mature neutrophils capable of regulating many biological functions, such as transmigration by lowering their extracellular concentration available for receptor binding (Hofman et al, 1998; Malfroy et al, 1988; Mishra et al, 2016; Roques et al, 1993; Turner and Tanzawa, 1997). In addition, an upregulation of CD10 was observed in neutrophils during Staphylococcus aureus and mononucleosis infection; however, septic patients show a decreased CD10 expression (Martens et al, 1999). Despite these results, the role of CD10 as CD10-expressing neutrophils during bacterial or viral infections is unknown. However, CD10+ neutrophils can inhibit T cell activation, proliferation, and interferon-gamma production (Marini et al, 2017).
In another study, the percentage of CD10+ neutrophils correlated with infection severity (Morisaki et al, 1992). Taking this background into consideration, it is possible to suggest the active participation of these cells during DENV infection and clinical complications. Previous work by our laboratory group demonstrated a significant reduction of TREM-1 on neutrophils of dengue-infected individuals, as well as high levels of circulating sTREM-1 compared to healthy individuals (Ruiz-Pacheco et al, 2014). These results show the involvement of TREM-1 during DENV infection; however, it is poorly known whether DENV induces TREM-1 on neutrophils or whether these cells are a source of sTREM-1 during infection. The aim of this study was to investigate whether DENV could induce the expression of TREM-1, CD10, and sTREM-1 in human neutrophils.
Methods
Dengue virus
As previously reported, DENV-2 was obtained and expanded from a blood sample taken from a DENV-infected patient (Limon-Flores et al, 2005). Serotyping of this new isolate, DENV-2, is routinely performed in the laboratories of the Institute of Epidemiological Diagnosis and Reference (InDRE), Mexico City. The virus was then titrated by the standard plaque-forming assay using the BHK-21 cells. In brief, a 10-fold serial dilution of the virus was added to BHK-21 monolayers cultured in 24-well plates at 2.5 × 105 cells/mL, and these were incubated at 37°C for 4 h (Talavera et al, 2004). Subsequently, 0.5 mL minimum essential medium (MEM) containing 10% fetal bovine serum (FBS) and 3% (w/v) carboxymethylcellulose was added to each well.
After 5–6 days at 37°C, visualization of the plaques was done by staining with naphthol blue-black. Virus titers are expressed as plaque-forming units (pfu)/mL. Afterward, the virus was aliquoted and stored at −70°C until use. In some experiments, the virus was inactivated (heat-inactivated dengue virus [iDENV]) using a high temperature, 56°C for 30 min (Libraty et al, 2001).
Neutrophils
For the isolation and enrichment of neutrophils, buffy coats obtained from healthy donors (n = 9) from the Blood Bank of the Hospital General Rubén Leñero, Mexico City, were used. Ethical approval was obtained from the Institutional Review Committee at the Instituto de Diagnóstico y Referencia Epidemiológicos, Secretaría de Salud, Mexico City, Mexico. Twenty milliliters of blood were placed in a 50 mL tube with an equal volume of 3% dextran solution (Sigma-Aldrich, Poole, UK) and allowed to settle for 20–30 min as previously described (Nauseef, 2007). The leukocyte-rich plasma was then transferred to a new tube previously loaded with Ficoll-Hypaque (GE Healthcare, Uppsala, Sweden).
Immediately, the cells were centrifuged at 400 g for 30 min. The neutrophil-rich pellet was resuspended in sterile water and an equal volume of 1.8% saline solution was immediately added and gently homogenized, before, the sample was centrifuged at 400 g for 5 min and after centrifugation, the cells were resuspended in 5 mL of Roswell Park Memorial Institute (RPMI) 1640 (Gibco). The percentage of purity according to the expression of CD10 by cytometric analysis was 97.90 ± 1.15, mean ± standard deviation (SD) (Fig. 1). In addition, the viability of neutrophils in culture was analyzed through the incorporation of 7-Aminoactinimycin D (7AAD) (BD Biosciences, San Jose, CA) (Fig. 2).
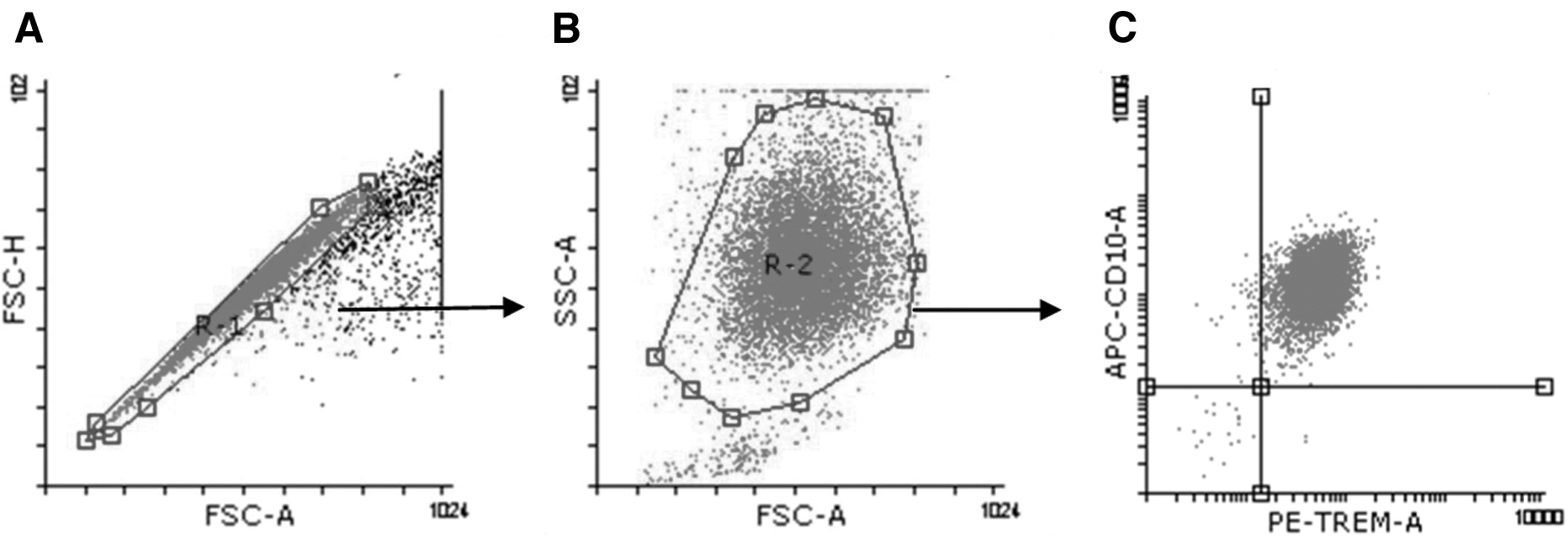
Representative of the analysis of cell-surface TREM-1 and CD10 expression.
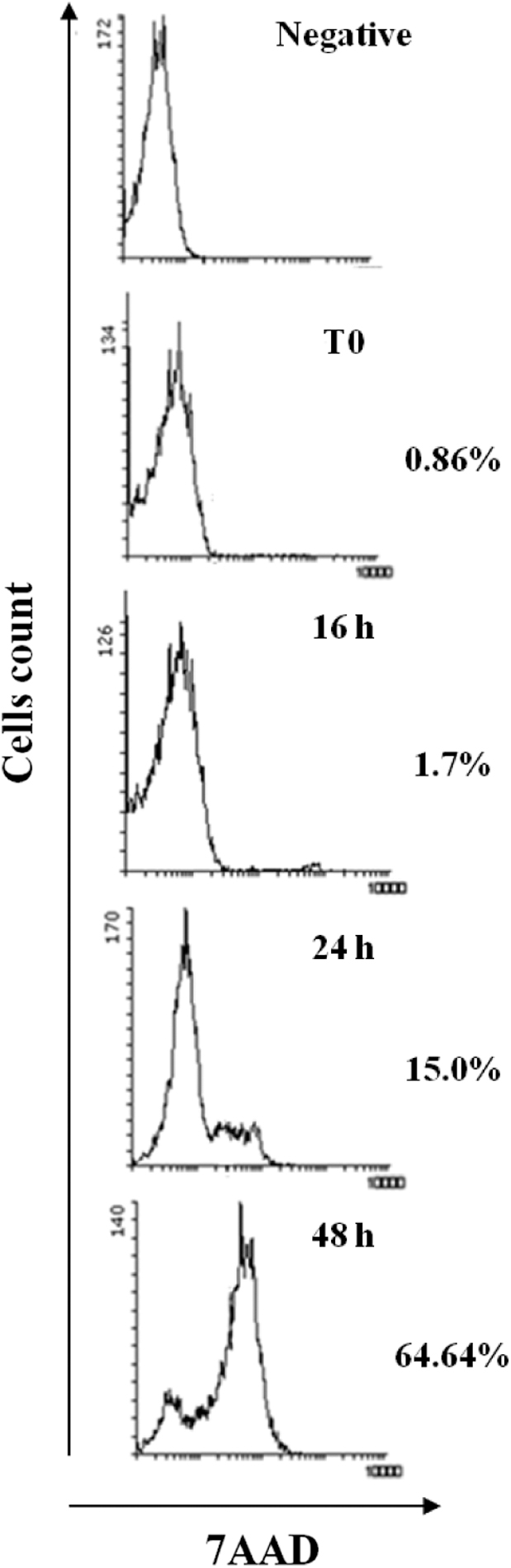
Percentages of viable peripheral neutrophils after isolation. Purified neutrophils were obtained from healthy donors (n = 9), which were cultured (1 × 106 cells/mL) for 16, 24, and 48 h. After the incubation time, the cells were collected and stained with anti-CD10 (neutrophil marker) and 7-AAD (viable dye). At least 10,000 CD10+ cell events were collected and the percentage of 7AAD-positive cells was quantified by flow cytometry. 7AAD, 7-Aminoactinimycin D.
TREM-1 and CD10 expression
To determine the effect of DENV-2 on TREM-1 expression on the neutrophils, in 24-well plates (Costar, NY) 1 × 106 cells were seeded and infected with DENV-2 at 1, 4, and 10 multiplicities on the infection (MOI). After 16 h of incubation, the expression of TREM-1 and CD10 were done by cytometric analysis. In some experiments, 2.5 × 105 neutrophils were cultured in 96-well plates (Costar) containing 0.01, 0.1, 1, and 10 ng/mL human recombinant granulocyte-macrophage colony stimulating factor (GM-CSF) (Peprotech, London, UK) in presence or absence of DENV-2 (m.o.i. = 4) during 1, 5, and 16 h.
Flow cytometry
At the end of each incubation, 1 × 106 cell/mL of human neutrophils were stained with CD10-fluorescein isothiocyanate (APC) (Biolegend, San Diego, CA), and TREM-1 phycoerythrin (PE) (Biolegend). After 30 min of incubation, cells were washed and analyzed using a FACS Calibur Flow Cytometer (BD, La Jolla, CA). At least 10,000 events corresponding to CD10+ were captured. Flow data analysis for frequencies and median fluorescence intensity (MFI) values were calculated using Flowing Software V 2.5.1 (Turku Centre for Biotechnology, Finland).
Statistical analysis
Experiments were performed in triplicate and values are presented as mean ± SD. One-way analysis of variance followed by Dunnett's post-test multiple comparison tests was used to compare responses between controls and treatment groups. Mann–Whitney test was used depending on data characteristics. A p-value of <0.05 was considered significant. Analysis of all data was performed using GraphPad Prism (version 5; GraphPad Software).
Results
DENV-2 upregulates TREM-1 expression in purified human neutrophils
To understand the effects of DENV-2 on TREM-1 expression, we first determined the viability of neutrophils in culture. Our results showed the viability of 98%, 85%, and 35% at 16, 24, and 48 h of culture, respectively (Fig. 2). According to the above, all our experiments were performed at 16 h. Subsequently, neutrophils from a healthy donor (n = 9) were cultured for 16 h with three different concentrations of DENV-2 (multiplicity of infection [MOI]: 1, 4, and 10).
There was a significant increase of TREM-1 in cultured neutrophils with MOI = 4 and 10 compared with control (Ctrl 41.91 ± 13.80 vs. m.o.i. = 4: 57.52 ± 6.56**; MOI = 10: 53.95 ± 7.12*, mean ± SD). In addition, there was no significant difference in TREM-1 expression between neutrophils cultured with m.o.i. of 4 compared to m.o.i. = 10. This is the reason why all our subsequent experiments were carried out with m.o.i. = 4 of DENV-2, (Fig. 3).

TREM-1 increases on neutrophils exposed to DENV-2. Neutrophils previously purified (1 × 106/mL) from blood obtained from healthy donors (n = 9) were cultured for 16 h with iDENV (MOI 10) and different concentrations of DENV-2 (MOI 1, 4 and 10). After the incubation time, TREM-1 expression was quantified by cytometry in at least 100,000 granulocyte events. Significant differences between groups are shown (One-way ANOVA followed by Dunnett's post-test multiple comparison tests was used to compare responses between controls and treatment groups). *p = 0.05, and **p < 0.05. ANOVA, analysis of variance; Ctrl, control group; DENV, dengue virus; iDENV, heat-inactivated dengue virus; MOI, multiplicity of infection; MFI, median fluorescence intensity.
GM-CSF does not affect the expression of TREM-1 in the presence of DENV
GM-CSF has been shown to induce an increase in TREM-1 in monocytes and importantly severe dengue patients produce higher levels of GM-CSF compared with low dengue manifestation (Banerjee et al, 2017; Bozza et al, 2008). To evaluate the influence of GM-CSF on TREM-1 in neutrophils, 1 × 106 neutrophils were cultured at different times (1, 5, and 16 h) with different concentrations of GM-CSF (0.01, 0.1, 1.0, and 10 ng/mL) in the presence or absence of DENV-2 (m.o.i. = 4). TREM-1 expression increased significantly up to 16 h of culture with 0.1, 1.0, and 10 ng/mL of GM-CSF (Ctrl: 74.55 ± 6.72 vs. 0.1 ng: 96.53 ± 7.62**; 1.0 ng: 101.1 ± 5.67** and 10 ng: 98.52 ± 8.01**, mean ± SD).
TREM-1 expression was not significantly modified when 0.01 ng/mL GM-CSF was used compared to control (Ctrl: 74.55 ± 6.72 vs. 0.01 ng: 72.01 ± 5.21, mean ± SD), (Fig. 4A–C). Similar results were obtained when 1 × 106 neutrophils were simultaneously stimulated with GM-CSF/DENV-2. TREM-1 expression significantly increased at 16 h with 0.1, 1.0, and 10 ng/mL of GM-CSF and simultaneously cultured with DENV-2 m.o.i. 4 (Ctrl: 72.23 ± 5.44 vs. 0.1 ng/DENV-2: 73.73 ± 26.75*; 1.0 ng DENV-2: 86.12 ± 14.26** and 10 ng/DENV-2: 88.02 ± 13.12**, mean ± SD, respectively) compared 0.01 ng/mL of GM-CSF/DENV-2 and control, (Ctrl: 72.23 ± 5.44; 0.01 ng/DENV-2 36.82 ± 2.75, mean ± SD), (Fig. 4D–F).

GM-CSF increases TREM-1 expression on neutrophils.
DENV-2 upregulates CD10 expression in purified human neutrophils
Herein, CD10 was used for the phenotyping of human neutrophils; however, it can regulate neutrophil function. When 1 × 106 neutrophils were cultured with heat-inactivated DENV-2 (m.o.i. = 10) and different concentrations of live DENV-2 (m.o.i. of 1, 4, and 10) for 16 h, there was a significant increase in CD10 compared with control (Ctrl: 32.85 ± 3.15 vs. iDENV: 51.45 ± 17.44**; MOI = 1: 60.57 ± 14.87****; MOI = 4: 66.34 ± 9.23**** and MOI = 10: 60.47 ± 8.97****, mean ± SD). Like TREM-1, CD10 expression increased similarly regardless of the DENV-2 concentrations used (Fig. 5).

CD10 increases on neutrophils exposed to DENV-2. Neutrophils previously purified (1 × 106/mL) from blood obtained from healthy donors (n = 9) were cultured for 16 h with iDENV (MOI 10) and different concentrations of DENV-2 (MOI 1, 4 and 10). After the incubation time, CD10 expression was quantified by cytometry in at least 10,000 granulocyte events. Significant differences between groups are shown (One-way ANOVA followed by Dunnett's post-test multiple comparison tests was used to compare responses between controls and treatment groups). ***p < 0.05, and ****p ≤ 0.001.
GM-CSF upregulates CD10 expression in purified human neutrophils
Similar to TREM-1, we decided to evaluate the role of GM-CSF on CD10 expression. CD10 expression was significantly increased in neutrophils only during the first hour of culture in the presence of 0.1, 1.0, and 10 ng/mL GM-CSF (Ctrl: 243.0 ± 12.11 vs. 0.01 ng: 249.3 ± 8.79; 0.1 ng: 388.7 ± 21.83***; 1.0 ng: 401.9 ± 38.09*** and 10 ng: 373.4 ± 37.88***, mean ± SD), (Fig. 6A–C). However, when DENV was added in the presence of GM-CSF, the expression of CD10 was elevated during 1 and 16 h.
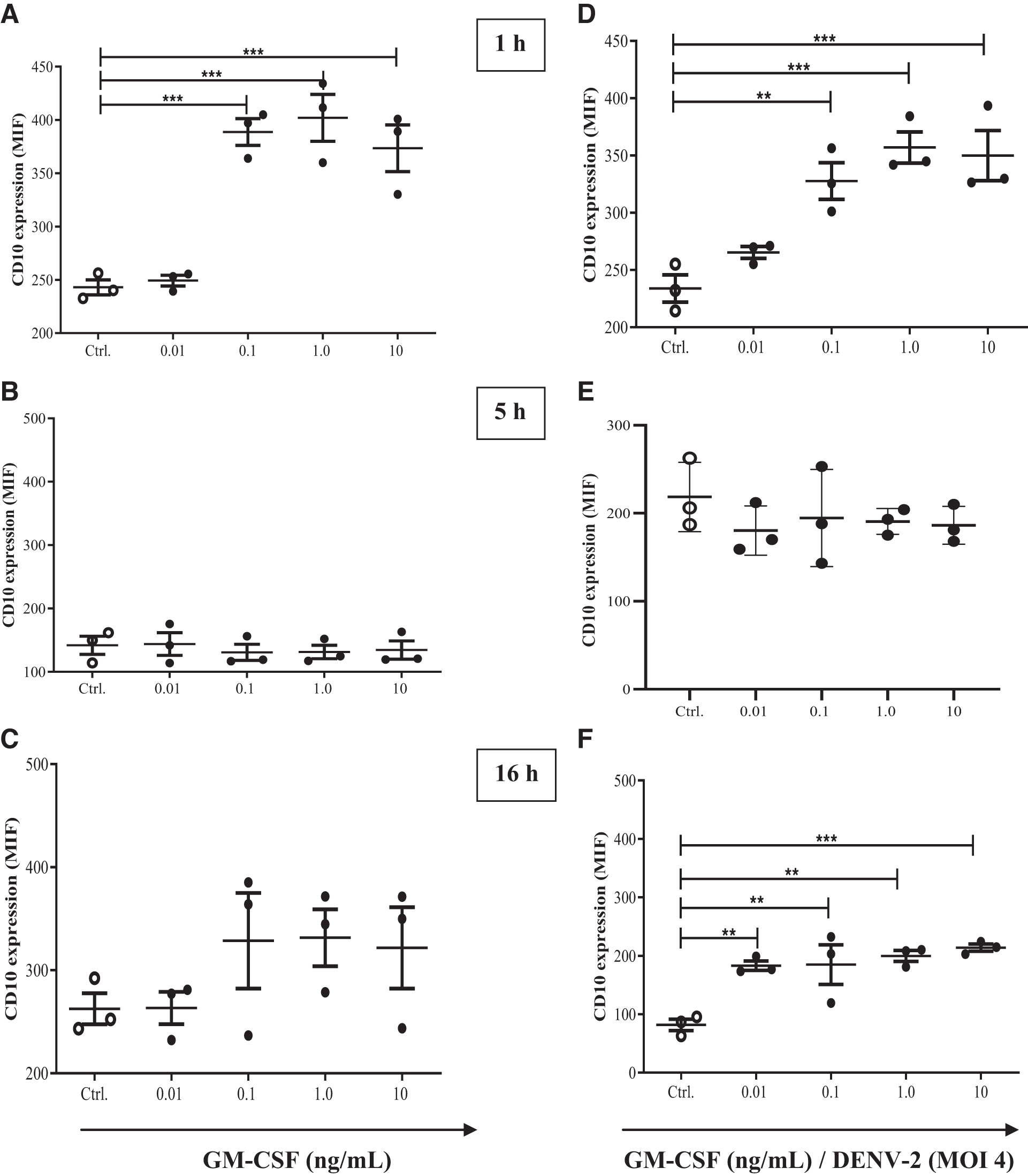
GM-CSF increases CD10 expression on DENV-exposed neutrophils.
Interestingly, when DENV-2 was added, a significant increase in CD10 expression was observed in cells cultured with 0.01 ng of GM-CSF (1 h Ctrl: 233.9 ± 20.48 vs. 1 h 0.1 ng/DENV-2: 327.7 ± 27.69**; 1 h 1.0 ng/DENV-2: 357.0 ± 23.60***; 1 h 10 ng/DENV-2: 349.9 ± 37.83***, and 16 h Ctrl: 81.70 ± 16.71 vs. 16 h 0.01 ng/DENV-2: 183.0 ± 13.90**; 16 h 0.1 ng/DENV-2: 184.9 ± 58.72**; 16 h 1.0 ng/DENV-2: 199.7 ± 16.32** and 16 h 10 ng/DENV-2: 213.9 ± 10.58***, mean ± SD), (Fig. 6D–F).
Neutrophils produce sTREM-1 in the presence of DENV-2
sTREM-1 production is considered a marker of membrane-anchored TREM-1 activation and the determination of its levels could help with the diagnosis and severity of infection (Cao et al, 2017). DENV-2 is capable of increasing sTREM-1 levels in cultured neutrophils, (Fig. 7).
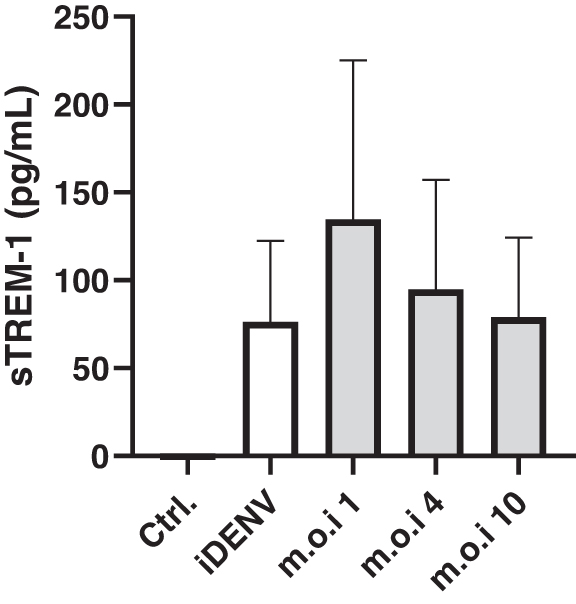
Soluble TREM-1. Neutrophils were incubated with live or inactivated DENV-2 for 16 h. Afterward, cell supernatants were harvested and sTREM-1 was detected by ELISA as described in Materials and Methods.
Discussion
TREM-1 is mainly expressed in neutrophils and its activation increases the inflammatory response (Bouchon et al, 2000). Unfortunately, the function of TREM-1 during viral infections remains poorly understood. During human immunodeficiency virus (HIV), Marburg virus (MARV), Ebola virus, and West Nile virus infection the phosphorylation of DAP12, TREM-1 shedding, and inflammatory genes have been reported (Denner et al, 2013; Mohamadzadeh et al, 2006; Suthar et al, 2013). In addition, we show for the first time changes in TREM-1 on neutrophils and increased sTREM-1 in dengue patients (Ruiz-Pacheco et al, 2014). However, we do not know whether neutrophils could be a source of sTREM-1 during DENV-2 infection.
We show that at least in vitro, DENV-2 is a sufficient stimulus to significantly increase the expression of TREM-1. Results that contrast with our previous publication where we demonstrated a reduction of TREM-1 on neutrophils obtained from patients with dengue (Ruiz-Pacheco et al, 2014). One possible explanation for this is that dengue patients experience aberrant immune activation and granulocytopenia where TREM-1-high neutrophils are possibly sequestered to target organs, while TREM-1-low neutrophils are released from the bone marrow, raising the impression of downregulated TREM-1 expression on circulating neutrophils (Butthep et al, 1993; Knapp et al, 2004; Liu et al, 2002).
However, the exact role of TREM-1 during virus infection, especially during dengue disease remains uncertain. Macrophages stimulated with double-stranded RNA showed an increase in TREM-1 in a dose- and time-dependent manner that was related to type I interferon, RIG-I-like receptors, and toll-like receptors expression (Zhu et al, 2016). Similarly, after internalization of Ebola and MARVs by neutrophils, there was transient phosphorylation of DAP12, shedding of TREM-1, and production of IL-1, TNF-α, IL-6, IL- 8, MIP-1, and MPO (Mohamadzadeh et al, 2006). Hepatitis B virus infection is also associated with the upregulation of TREM-1 on monocytes and high levels of TNF-α, IL-1β, and IL-6 (Wu et al, 2021).
During HIV infection, the expression TREM-1 on macrophages was correlated with antiapoptotic protein expression and high survival of cells (Campbell et al, 2019; Yuan et al, 2017), revealing that TREM-1 might have a critical role in establishing latent reservoirs. In addition, activation of TREM-1 and proinflammatory genes have also been associated with enterovirus A71 (EV-A71) infection (Amrun et al, 2020).
Some soluble molecules, such as GM-CSF, have been shown to enhance TREM-1 expression in human monocytes in response to LPS and 19 kDa lipoprotein (Bleharski et al, 2003). Considering that GM-CSF is widely produced during DENV-2 infection and its presence has correlated with protein dysregulation in neutrophils (Bozza et al, 2008; Wittmann et al, 2004), we decided to assess the effects of different concentrations of GM-CSF at different times when DENV-2 was added or not. The results showed that concentrations of 0.1, 1.0, and 10 ng/mL of GM-CSF were sufficient to significantly induce TREM-1 expression in the neutrophil membrane at 16 h of culture. Similar results were observed when GM-CSF and DENV-2 were added, demonstrating that the expression of TREM-1 does not differ significantly between neutrophils cultured with GM-CSF or GM-CSF/DENV-2.
The upregulation and aggregation of TREM-1 is a step that precedes its own solubilization in an MMP9-dependent manner (Jolly et al, 2021). Dengue and Crimean Congo hemorrhagic fever patients have high levels of sTREM-1 than the control group (Altay et al, 2016; Ruiz-Pacheco et al, 2014). Importantly, the levels of sTREM-1 have correlated with the C reactive protein, white blood cell, and platelet count (Altay et al, 2016). Recently, sTREM-1 has been proposed as a new ligand for the membrane receptor Robo2, a receptor related to liver damage (Liu et al, 2021). Hepatic dysfunction is also a severe complication of dengue infection (Samanta and Sharma, 2015). Therefore, sTREM-1 could be used as a marker of membrane-anchored TREM-1 activation and a predictor of the severity of viral infections (de Silva-Neto et al, 2021).
Here, sTREM-1 was elevated in the supernatants of neutrophils incubated with DENV-2, suggesting the activation of TREM-1 on neutrophils in the presence of DENV-2 and also demonstrating that neutrophils may contribute to elevated levels of sTREM-1 during dengue infection. In this sense, there are two hypotheses regarding the source of sTREM-1, translation of an alternative TREM-1 mRNA splice variant, or proteolytic membrane shedding of surface-expressed TREM-1 (Gingras et al, 2002).
In the case of neutrophils, both processes have been reported (Baruah et al, 2015; Gomez-Pina et al, 2007). Since we did not observe a reduction of TREM-1 in the neutrophil membrane, we suggest that the production of sTREM-1 can be done through to alternative TREM-1 mRNA splice variant (Gomez-Pina et al, 2007). However, sTREM-1 role during dengue infection is uncertain. Nevertheless, it is possible that sTREM-1 could function as a decoy that competes with the TREM-1 ligand to inhibit its activation and prevent activation and damage caused by infection (Roe et al, 2014).
In addition, CD10 expression was used for neutrophil phenotyping (McCormack et al, 1986). Due to the enzymatic activity linked to CD10 and the lack of knowledge about its participation in viral infection (Morisaki et al, 1992; Roques et al, 1993; Turner and Tanzawa, 1997), we decided to evaluate the expression of CD10 in cultured neutrophils in the presence of DENV-2. Surprisingly, neutrophils purified and exposed to DENV-2 for 16 h significantly increased CD10 levels regardless of the concentration of DENV-2 used. This seems in marked contrast to the literature suggesting reduced levels of CD10 expression on neutrophils during human cytomegalovirus and SARS-CoV-2 infection (Phillips et al, 1998; Spijkerman et al, 2021).
The role of CD10 in viral infection is unknown; however, the overexpression of CD10 in melanoma cells was associated with the expression of genes related to apoptosis, WNT signaling, MAP kinase pathway, and antigen processing and presentation (Velazquez et al, 2007). Meanwhile, the expression of CD10 in T cells and neutrophils was associated with apoptosis and suppression of Th1 lymphocyte proliferation via CD18 contact-dependent production of arginase-1, respectively (Cutrona et al, 1999; Marini et al, 2017). In addition, the abrogation of CD10/NEP enzymatic activity is associated with a reduced amount of Met-enkephalin required for hemocytes activation (Shipp et al, 1990). In this case, the expression of CD10 could work as a virus-associated escape from T cell recognition and/or modulator of the activity of inflammatory mediators.
Finally, we also observed that GM-CSF treatment significantly increased CD10 expression during the first hour of culture compared to cultures at 5 and 16 h. Results are supported by other studies (Connelly et al, 1993). However, when DENV-2 was present, CD10 expression significantly increased after 1 and 16 h of culture, as well as using 0.01 ng/mL of GM-CSF, demonstrating that the molecules induced by dengue infection can also regulate the expression of CD10. In conclusion, we have shown that DENV-2 induces TREM-1 expression and sTREM-1 production in cultured human neutrophils.
In addition, we are the first to demonstrate an increase in CD10 on the membrane of neutrophils when they are cultured with DENV-2, a finding which can contribute to the understanding of pathogenic mechanisms of DENV-2; however, despite the statistical significance shown in our results, it is important to mention the size of the sample used in this work. Therefore, more investigations both in vitro and in vivo are required to establish the precise role of these molecules during DENV infection.
Footnotes
Authors' Contributions
J.R.P. and A.E.G. contributed to conception and design. E.M.M., and L.C.D. performed the collection and assembly of data. J.R.P., and R.C.S performed data analysis and interpretation. All authors contributed to the article writing and final approval of the article.
Acknowledgments
The authors thank Hospital General Ruben Leñero, Mexico City, for donating the buffy coats and for supporting this study.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This research did not receive any specific grant from funding agencies in the public, commercial, or not-for-profit sectors.