
Abstract
We report an asymptomatic child with heterotaxy syndrome who had recurrent positive SARS-CoV-2 polymerase chain reaction testing. An aberrant lymphocyte population expressing CD19, CD16, and CD56 was identified; its phenotyping revealing atypical NK cells. This subset's role in protection from severe disease or in reinfection cannot be ascertained.
Introduction/Background
As the COVID-19 pandemic is well into the third year, the role of reinfections has been studied, both in vaccinated and unvaccinated individuals (Abu-Raddad and Bertollini, 2021; Mensah et al, 2022; Wang et al, 2021). The phenomenon of reinfection is closely tied to the durability of the immune response and protection after exposure and/or first infection, and vaccination. Several studies have assessed the dynamics of the immune response, and its role in predicting reinfection (Cromer et al, 2021; Khoshkam et al, 2021; Mensah et al, 2022; Rahman et al, 2022; Sette and Crotty, 2021). Both humoral and cellular immunity are important in the control of SARS-CoV-2 infection, and the efficacy of the immune response likely plays a large role in determining the potential for reinfection.
However, the longevity and the robustness of the immune response to this virus may be of shorter duration than with certain other pathogens, due to the documented reinfection in vaccinated individuals as well as those who have been previously infected. Reinfection is dependent on several factors, including whether the original infection was caused by a different strain, resulting in less cross-reactive immune coverage and greater susceptibility to infections with newer strains (Cromer et al, 2021; Khoshkam et al, 2021; Mensah et al, 2022; Rahman et al, 2022; Sette and Crotty, 2021).
In addition, the duration of viral shedding may be a contributing factor as some individuals continue to carry the virus without new clinical symptoms. The role of T cells (CD4+ and CD8+) and B cells in the immune response and outcomes of SARS-CoV-2 infection have been explored in numerous studies (Cox and Brokstad, 2020; Khoshkam et al, 2021). However, innate lymphocytes, such as NK cells, have also been suggested to play a key role in the immune response to COVID-19, which would not be unexpected given the role that NK cells play in protecting against viral infections. NK cell activation may modulate the magnitude of the immune response to SARS-CoV-2 infection, with a specific threshold that facilitates protection, and a separate threshold that raises the risk for aberrant immune activation and associated pathological consequences.
We report in this study a pediatric patient with heterotaxy and polysplenia who had “recurrent” SARS-CoV-2 infection with no evidence of clinical disease, and the presence of an atypical NK cell subset expressing low levels of CD19, regarded as a primary B cell marker.
Methods
The patient was enrolled in a research protocol with written informed consent in accordance with Institutional Review Board (IRB) guidelines. A single sample of blood was collected on two separate occasions for detailed immunophenotyping under the research protocol.
Multiparametric immunophenotyping was performed in addition to routine clinically obtained lymphocyte subset (T, B, and NK cells) quantitation to further characterize the atypical lymphocytes, which expressed multilineage markers. Flow cytometry was performed on a whole blood (EDTA) sample along with a healthy experimental control using a panel of antibodies, which included CD19, CD16, CD56, NKp46 (CD335), CD21, CD24, CD38, CD10, CD27, IgM, and IgD. The samples were analyzed on a CytoFlex LX® flow cytometer (Beckman Coulter, La Brea, CA) and data analysis was performed with FCS Express v.7 software (De Novo). Other studies, including SARS-CoV-2 polymerase chain reaction (PCR), were obtained as part of routine clinical care.
Case Report and Results
The patient was a 7-year-old boy who presented with heterotaxy syndrome and complex congenital heart disease consisting of dextrocardia, pulmonary atresia, right-dominant unbalanced atrioventricular septal defect, interrupted inferior vena cava with hepatic veins draining to the atrium, and a malposed aorta off the right ventricle and extracardiac findings of polysplenia, midline liver, and intestinal malrotation. He has had multiple cardiac and intestinal procedures. The patient received prophylactic dosing of amoxicillin during the first 5 years of life and had no history of recurrent infections. He had an elective cardiac catheterization arranged to evaluate his hemodynamics before his planned Fontan surgery. As per institutional protocol for patients undergoing surgery, a nasopharyngeal swab was obtained for SARS-CoV-2 PCR testing, which was reported as positive.
The cycle threshold (Ct) value was not available with this particular assay. The patient had no symptoms indicative of SARS-CoV-2 infection at the time of the positive test and remained asymptomatic throughout the course of follow-up testing. After his positive SARS-CoV-2 result, both parents reported feeling fatigued with fevers but were not tested for SARS-CoV-2. His repeat SARS CoV-2 qualitative PCR using a laboratory-developed assay targeting nucleocapsid (N) targets a month later was reported as negative with N1 = 0 and N2 = 39.02 (Supplementary Fig. S1). He underwent cardiac catheterization after another negative PCR with no virus detected.
Three months later, he underwent cardiac surgery without complications and was discharged in 6 days. He was discharged with a PleurX chest tube as per our institutional postoperative management. He had multiplex PCR panel done before discharge. That test was negative for SARS-CoV-2 but positive for rhinovirus/enterovirus. He returned for elective PleurX chest tube removal and another preoperative SARS-CoV-2 PCR test 2 months later, which revealed a positive result. The Ct value was reported to be 33.10 by the Cepheid assay.
The sample was sent for gene sequencing but could not be sequenced. His SARS-CoV-2 IgG nucleocapsid antibody index 26 days after his last positive PCR test was negative at 0.9 (negative <1.40 index S/C). His quantitative immunoglobulins were normal. The patient had pneumococcal antibodies tested about 7 years after receiving four doses of pneumococcal conjugate PCV13 vaccine showing adequate response to six of the serotypes included in the vaccine. Adequate response for test purposes was 1–1.3 μg/mL.
A basic lymphocyte subset quantitation for T, B, and NK cells was performed on blood, which revealed moderate CD4+ T cell lymphopenia (346 cells/μL; range 468–2,416 cells/μL) with normal CD3+ and CD8+ T cell counts, CD19+CD20+ B cell counts, and total CD16+CD56+ NK cell counts. However, he was noted to have an unusual lymphocyte population, identified by CD19+CD16+ and CD56+/− (a little more than half these cells expressed CD56 and the remainder expressed CD16 without CD56) (Fig. 1). These cells were 17% of CD3-negative lymphocytes and 7% of total CD45+ lymphocytes.

Detection of the atypical CD19+CD16+CD56+/− population in patient whole blood immunophenotyping. Immunophenotyping of EDTA whole blood from
Additional immunophenotyping of T cells for naive and memory markers (data not shown) revealed decreased naive CD4+CD45RA+ T cells for age at 8% with the vast majority of CD4+ T cells being of the memory phenotype (CD4+CD45RO+). Recent thymic emigrants identified as CD4+CD45RA+CD31+ were also mildly decreased for age at 65% of CD4+CD45RA+ T cells or 21% of total CD4+ T cells. There was also an increase in terminally differentiated, senescent TEMRA CD8+ T cells (CD8+CD45RA+CCR7−CD62L−) for age at 11% of total CD8+ T cells (in contrast to 0.34% in the experimental healthy control).
B cell subset phenotyping (data not shown) revealed a population of dim CD19+ B cells. Analysis of the conventional CD19bright B cell subset revealed normal total memory B cells and plasmablasts expressing CD27, as well as normal switched memory B cells. However, analysis of the dim CD19+ B cells revealed that 71% expressed surface IgM, and 10% were positive for both IgM and IgD expression. About 25% of these CD19dimCD27− B cells expressed low levels of IgG, and the vast majority (89%) were immature B cells negative for CD21 (data not shown). Other B cell markers were negligible in this subset. The expression of surface IgM and/or IgD suggested that this dimCD19 population identified on the B cell subset analysis are indeed B cells, and not the CD19+CD16+CD56+/− subset, identified on the TBNK flow cytometry analysis, which was evaluated separately, as described hereunder.
The atypical population identified in the patient (CD19+CD16+CD56+/−) was further analyzed for several B cell markers as well as CD335 (NKp46), an NK cell marker (Fig. 2), which revealed that this subset was only present in the patient but not the healthy control (an analytically negligible population). These cells were positive for NKp46 but negative for CD21 and CD27 (Fig. 2). Interestingly, these cells expressed CD24 at very low levels similar to NK cells from the patient and essentially no CD38 (in contrast to the healthy control, which expressed CD38 and no CD24). The pattern of expression of these markers also did not follow the expression of CD24 and CD38 seen on transitional B cells (Fig. 2). These NKp46+ cells were negative for IgG, similar to classical NK cells, and unlike B cells, which express IgG (data not shown).
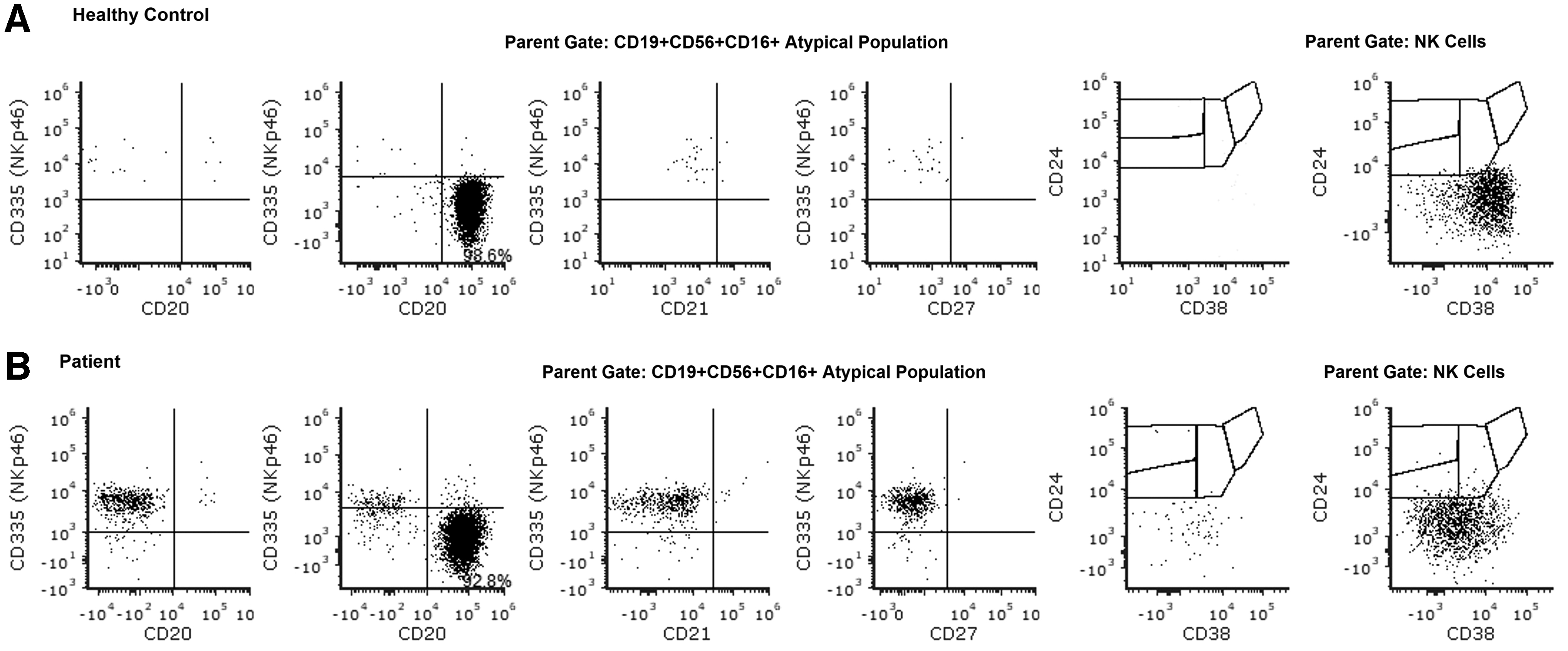
Detailed immunophenotyping of the atypical population. Analysis of CD335 (NKp46) expression in the atypical CD19+CD56+CD16+ population showed presence of CD335+ cells only in the patient, but not in the healthy control. No CD21 and CD27 expression were detected on these cells. These atypical cells resembled NK cells from the patient, in terms of dim CD24 expression and no CD38 staining, whereas atypical population and NK cells from the healthy control were CD38+CD24−. Row
Therefore, these atypical cells expressing CD19 appear to be NK cells expressing CD16 with two subsets, one, positive for CD56, and the other, negative.
Discussion
A considerable body of literature is available on SARS-CoV-2 infection, and overall, the relative frequency of infection with severe clinical manifestations remains rare in children (Brodin, 2022). Most infected children appear to have asymptomatic disease, although this may be related to the prevalent strain dominant at the time in the population. Although this is good news for pediatric population, it makes the identification of true reinfection challenging, when coupled with the potential for prolonged viral shedding, and is further complicated by variability in analytical testing. Reinfections with repeat positive SARS-CoV-2 PCR results have been shown to be extremely uncommon (Peltan et al, 2021). Our patient had a relatively higher risk of developing severe disease, yet he remained asymptomatic. His SARS-CoV-2 reinfection was identified while undergoing presurgery screening.
It is generally considered unlikely to have SARS-CoV-2 recurrence in the absence of concurrent clinical syndromes, and also exhibiting a downward trajectory of Ct values with a very low Ct value at recurrence. Whether these patients have asymptomatic recurrence, reinfection, or persistent shedding of viral RNA is unknown. In some cases, the interval of testing has been less than a month in samples with a repeat positive result (Lu et al, 2020). Although the unavailability of viral genome sequencing and serial antibody titers along with use of a variety of different SARS-CoV-2 detection tests are limitations of this study, the prolonged period of negativity in between positive tests with highly sensitive testing platforms are convincing for reinfection. We were unable to assess whether the patient had a different virus variant with his second infection further limiting the assessment of his immune response.
The asymptomatic nature of SARS-CoV-2 reinfections in a patient with multiple risk factors associated with heterotaxy syndrome, congenital heart disease with single ventricle physiology, and multiple prior surgeries makes this case particularly intriguing. It is notable that he had a PleurX chest tube in place without significant increase in drainage or other clinical changes before retesting positive. There is no known association with the observed clonal change and protection from symptomatic illness.
Atypical NK cells expressing CD19 have been previously reported, where it has been shown to confound the analysis for minimal residual disease for B-acute lymphoblastic leukemia (Soma et al, 2015). Another study observed this subset in several pediatric patients being evaluated for a suspected immunodeficiency, although it was not specified as to which genetic immunodeficiencies it was associated with, and it was suggested to be present at a frequency of 4.4% in this cohort (Korol et al, 2015). We have identified several patients (with malignancies, autoimmunity, hyperinflammation, and infection) who have been identified with this atypical lymphocyte subset, suggesting that this population is present more frequently than previously thought, and can only be identified by laboratories, which use the right combination of markers and gating strategy.
Korol et al (2015) observed that CD19 expression on these cells is dimmer than normally observed in typical B cells. The mechanism of the acquisition of CD19 by these NK cells is unclear, although it is possible it could be by trogocytosis, as has been reported in patients treated with epratuzumab (anti-CD22) who acquire several B cells markers on FcγR-expressing cells, including monocytes, NK cells, and neutrophils (Rossi et al, 2013). Some studies suggest that cytoplasmic expression of the CD3 epsilon-chain by the CD19+ NK cells indicate a specific activated state of this subset (Korol et al, 2015; Lanier et al, 1992).
We did not observe CD3-epsilon expression (data not shown) in this atypical NK cell subset; however, that does not mean this population is not activated. It remains unclear as to what immunological role this population might play in modulation of the immune response in various contexts. Although we have observed the presence of this subset in several pediatric patients with a variety of clinical diagnoses, it is unclear whether this population of atypical NK cells is actually active in the immune response in those contexts. In some patients with dim CD19+ B cells, this subset is negative for CD16 and CD56 but may express NKp46.
It remains unclear as to how these cells acquire CD19, and if by trogocytosis, does it represent an epi-phenomenon or does it identify a subset of activated NK cells participating in the immune response against viral or other infections. It is unlikely that the presence of this population is related to the heterotaxy in this patient, as there is no objective evidence or data from the literature to support this. This finding, although anecdotal in the context of asymptomatic SARS-CoV-2 infection, may represent yet another facet of the complex immune response to this pathogen, and may warrant scrutiny in larger populations.
Through this report, we attempt to provide insight into the clinical and biological significance of unusual immune cell subsets, which may identify novel pathways of immune activation and immune responsiveness, potentially in the context of SARS-CoV-2 infection, although we currently do not have any evidence of what role these cells may play in this patient's phenotype or in the other contexts where it has been observed. There are very little data, if any, on the impact of heterotaxy on quantitative distribution of lymphocyte subsets. Patients with heterotaxy have been reported to have an increased risk of infections due to the functional asplenia (Kothari, 2014). Thus, it is unclear if the patient's immunological phenotype has any relationship with his heterotaxy. Nonetheless, individual case reports, such as this, can contribute meaningfully to the literature, especially in the context of rare observations.
Footnotes
Authors' Contributions
Conceptualizing (lead), writing (lead), and review and editing (equal) by G.E. Conceptualization (supporting) and writing—review and editing (equal) by C.L.C. and S.L. Methodology (supporting) and writing—review and editing (equal) by A.B. and A.L. Formal analysis (lead), methodology (lead), writing (lead), and review and editing (equal) by R.S.A.
Author Disclosure Statement
The authors have no conflicts of interest relevant to this article to disclose. The information in this article has not been previously presented.
Funding Information
No funding was received for this article.
Supplementary Material
Supplementary Figure S1
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.