
Abstract
Human T lymphotropic virus 1 (HTLV-1) is a retrovirus associated with inflammatory diseases, including HTLV-1-associated myelopathy (HAM), and host genetic factors may be involved in disease evolution. The forkhead Box P3 (FOXP3) transcription factor is linked to homeostasis of the immune system, and the presence of polymorphisms in the promoter region of the FOXP3 gene should reflect its expression levels and consequent activation of regulatory T cells, which may contribute to severe inflammatory disorders, such as HAM. This study evaluated the rs2232365 polymorphism (−924 A/G) located in the promoter region of the FOXP3 gene and its association with HAM. Forty DNA samples from asymptomatic carriers and 25 samples from HAM patients were used, in addition to 130 control samples. The polymorphism was genotyped by conducting real-time polymerase chain reaction (PCR) (quantitative PCR [qPCR]) on extracted DNA. The proviral loads (PVLs) and CD4+ and CD8+ T lymphocyte counts were determined by qPCR and FACSCalibur flow cytometry, respectively. The PVLs, CD4+ T lymphocyte concentrations, and tumor necrosis factor-α dosages were considered predictive factors of the clinical profiles of HTLV-1 infection, all of which had higher levels in the HAM group. Carriers of the GG genotype for the polymorphism rs2232365 had high PVLs and CD4+ T lymphocyte concentrations.
Introduction
Human T lymphotropic virus type 1 (HTLV-1) is a retrovirus associated with persistent infection of human T cells, preferentially CD4+ T lymphocytes (Kannian et al, 2012). The virus infects up to 10 million people worldwide and is endemic mainly in southwestern Japan, the Caribbean, South America (Brazil and Peru), and West Africa (Malpica et al, 2018). HTLV-1 is associated with adult T cell leukemia/lymphoma (ATL), HTLV-1-associated myelopathy (HAM), and other diseases, such as uveitis, rheumatoid arthritis, Sjögren's syndrome, skin, and lung diseases (Dias et al, 2018; Pinheiro et al, 1995; Proietti et al, 2005).
HAM, which is characterized as an inflammatory disease of the central nervous system in which patients present spastic paraparesis and lower limb sensory disturbances, is the most common clinical manifestation of the virus. The disease develops in only 5% of infected individuals, and the majority of patients are female. An increased proviral load (PVL) of HTLV-1 is a strong predictor for the disease, but the exact mechanisms by which HTLV-1 induces these clinical conditions remain unclear (Araújo and Silva, 2006; Fuzii et al, 2014; Osame, 2002).
It is proposed that the expression of a viral transcriptional activator known as Tax may alter the regulation of cellular genes, which leads to the processes of T cell activation and proliferation (Casseb and Penalva-de-Oliveira, 2000). In addition, it is suggested that HAM development seems to be regulated by the interaction between the host immune response and virus-infected cells, where cellular immune responses may have an important role in disease evolution (Nozuma et al, 2020).
It is already known that the numbers of HTLV-1-infected T cells that circulate in the peripheral blood are higher in patients with HAM than in asymptomatic carriers (Pinto et al, 2015). Another hypothesis is that the genetic background of the host plays a role in the etiology of HAM. From this perspective, several studies have been carried out on host genetic factors that can lead to exacerbated immune responses against HTLV-1 carriers, including HLA haplotypes (Benencio et al, 2020) and polymorphisms in genes such as FAS (cell surface receptor Fas), interleukin (IL)10 and Mannose-binding lectin (MBL) (Alves et al, 2007; Braz et al, 2019; Vallinoto et al, 2018).
The Forkhead box protein P3 (FOXP3) gene is located on the short arm of the sexual human chromosome, Xp 11.23-Xql (3.3 locus; 21 kb), and consists of 11 exons, which encode 431 amino acid proteins (Bennett et al, 2001; Jiang et al, 2014). FOXP3 is an important transcription factor that is causally linked to the establishment and maintenance of regulatory T cells (Tregs) (Lu et al, 2017; Pererira et al, 2017); therefore, studies have been performed to understand the biological properties of FOXP3 and its impact on the immune response to infectious diseases (Beiranvand et al, 2017; Chen et al, 2013; Mercer and Unutmaz, 2009; Pereira et al, 2018; Satou et al, 2012).
In HAM patients, Foxp3 mRNA levels seem to be lower than those in asymptomatic carriers, suggesting that virus infection can alter the expressions of functional transcription factors, resulting in immune dysfunction and evolution to HAM (Ghezeldasht et al, 2017; Miyazato et al, 2014). The presence of polymorphisms in the FOXP3 gene has also been investigated (Oda et al, 2013), and it has been reported that T cell activation may be altered by the presence of single nucleotide polymorphisms (SNPs) in the promoter region of the gene, which modifies its transcriptional activation, interactions with regulatory elements for gene expression, and, as a consequence, expression levels of the gene (Hoogendoorn et al, 2003).
Thus, this study aimed to investigate whether the rs2232365 polymorphism located at the promoter region of the FOXP3 gene is related to functional changes that may predispose individuals to HAM.
Materials and Methods
Study population
A total of 65 HTLV-1-infected individuals were evaluated: 40 from asymptomatic carriers and 25 from patients with confirmed HAM diagnoses, who were of both sexes (44 females and 21 males), older than 40 years old (2 HAM individuals were 28 and 37 years old), not being treated with glucocorticoids, and without dermatological and rheumatological diseases associated with HTLV-1. HTLV-1-positive patients were classified as asymptomatic or with HAM according to pre-established criteria (Castro-Costa et al, 2006). Individuals were seen at the outpatient clinic of the Tropical Medicine Nucleus of the Federal University of Pará.
The project was approved by the Ethics Committee for Research with Human Beings of the Institute of Health Sciences of the Federal University of Pará (CAAE: 73782017.8.0000.0018). All participants were informed about the objectives of the study and signed a Free and Informed Consent Form.
Sample collection and storage
Blood samples (10 mL) were collected from all participants by intravenous puncture using a vacuum collection system containing ethylenediaminetetraacetic acid as an anticoagulant. Samples were centrifuged and separated into plasma and leukocyte fractions. The leukocyte samples were used for genomic DNA extraction for polymorphism analysis and PVL quantification and were stored at −70°C until use.
DNA extraction
DNA was extracted from peripheral blood leukocytes using the Puregene kit (Puregene, Gentra Systems, Inc.) according to the manufacturer's protocol, which included cell lysis, protein precipitation, and DNA precipitation and hydration steps.
Genotyping of FOXP3-924 A/G (rs2232365)
The SNP, rs2232365 (−924A/G), of FOXP3 was investigated by real-time quantitative polymerase chain reaction (qPCR) using the StepOnePLUS™ Real-Time PCR system (Thermo Fisher, Carlsbad, CA). The reaction used commercial TaqMan® SNP Genotyping Assays (C_15942641_10) containing primers and probes specific to the target sequence (Thermo Fisher). To amplify and detect alleles, 50 ng of DNA was used in a final reaction volume of 10 μL under the following conditions: 60°C for 30 sec and 95°C for 10 min, followed by 40 cycles of 95°C for 15 sec, 60°C for 1 min, and 60°C for 30 sec.
Quantification of HTLV-1 PVL and CD4+/CD8+ T lymphocytes
PVL quantification was based on previously described protocols (Tamegão-Lopes et al, 2006). Quantifications of CD4+ T lymphocytes (LTCD4+) and CD8+ T lymphocytes (LTCD8+) were performed by immunophenotyping by flow cytometry (FACSCount™ Reagent-TriTEST™/TruCount; BD Biosciences, San Jose, CA).
Measurement of cytokines
Plasma concentrations of IL-10, IL-6, IL-8, interferon-γ, tumor necrosis factor (TNF)-α, and TNF-β (cytokine Th1/Th2 profile) were measured by an enzyme-linked immunosorbent assay (Human ELISAReady-SET-Go; EBioscience, Inc., San Diego, CA). Kits were used to detect each cytokine with a specific monoclonal antibody. The procedure was performed according to the manufacturer's instructions.
Statistical analysis
To assess the homologies and differences among the clinical classes of HTLV-1 infection, a discriminant multivariate analysis was performed using data that quantified the PVLs, T lymphocyte concentrations and cytokine dosages. Then, a principal component analysis was performed to indicate the relative significances of the predictive variables related to the generated clusters. The suggested predictive variables were confirmed by comparing the data between the asymptomatic groups and those with HAM using Student's t-tests or Mann–Whitney tests, according to the normality of the data previously calculated by the D'Agostino test.
The associations of the predictive variables with sex and with genotypes of the rs2232365 polymorphism were evaluated with Student's t-tests or Mann–Whitney tests, according to the normality of the data previously calculated by the D'Agostino test. The associations of the frequencies of sex and rs2232365 polymorphism genotypes and alleles with the clinical profile of HTLV-1 infection were evaluated by the G test. FOXP3 is located on the sexual chromosome, “X,” and the adherence of rs2232365 to Hardy–Weinberg equilibrium was calculated only in women to evaluate whether the distributions of genotype frequencies were consistent with the expected results. Statistical analysis was performed using BioEstat 5.0 (Ayres et al, 2008) and GraphPad Prism 6.1, with a significance level of 95% (p < 0.05).
Results
Predictive variables of clinical profiles of HTLV-1 infection
A discriminant scatter diagram showing the multivariate analysis of asymptomatic and HAM carriers was created. These two groups tended to cluster at different graph coordinates without significant overlaps (Fig. 1A). We observed that PVLs, CD4+ T lymphocyte concentrations, and TNF-α dosages were the main components related to the clinical profiles of HTLV-1 infection, which account for ∼62% of the accumulated variance among the evaluated groups (Fig. 1B).
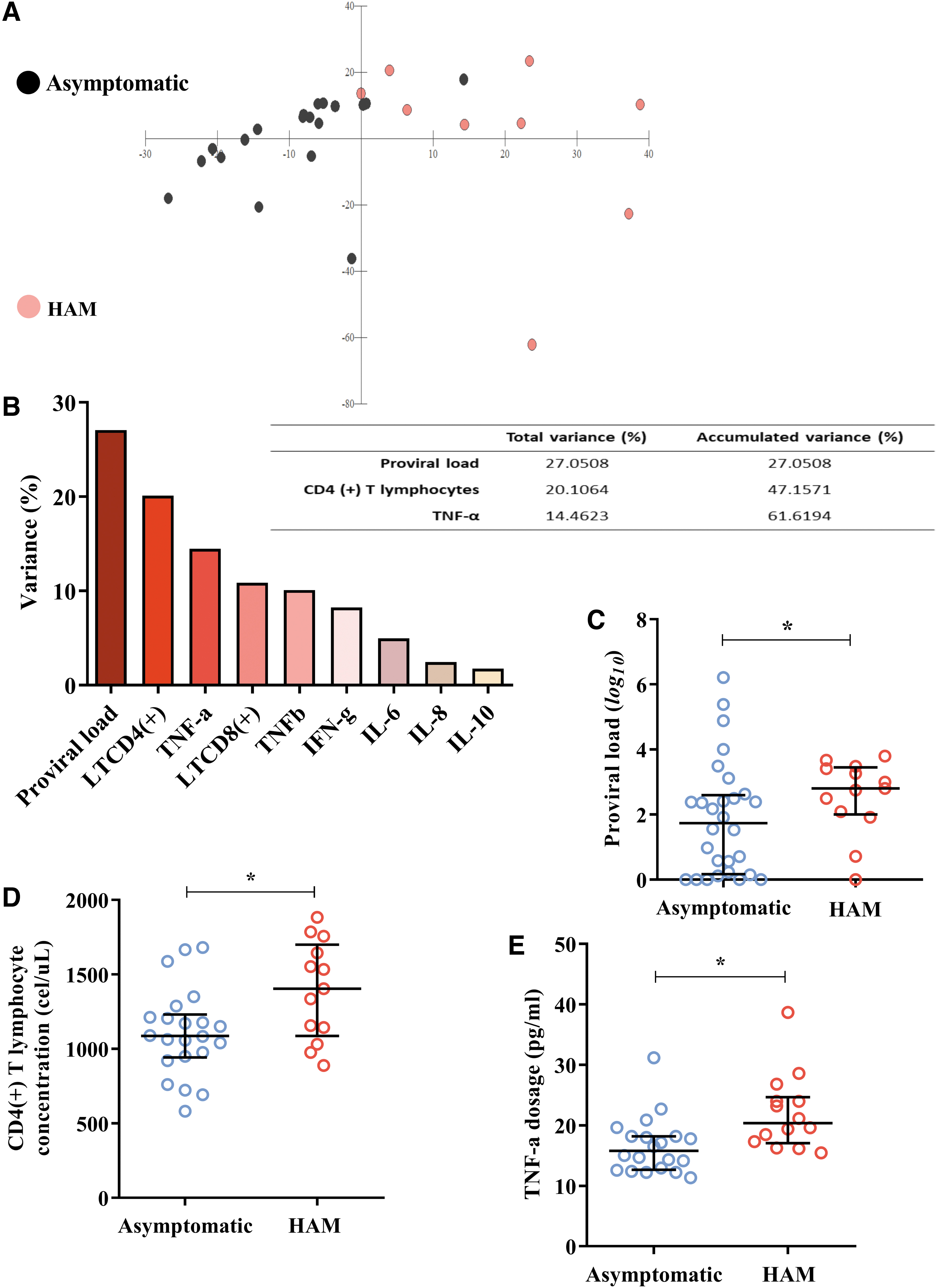
Predictive variables of clinical profiles of HTLV-1 infection:
We confirmed these findings when comparing the three variables among the clinical groups. The median log 10 PVL was higher (p: 0.0411) in the group with HAM [median (IQR 25%–75%): 2.80 (2.09–3.41)] than in the asymptomatic group [median (IQR 25%–75%): 1.74 (0.21–2.53)] (Fig. 1C). The mean concentration of CD4(+) T lymphocytes was also higher (p: 0.0132) in the HAM group [ (σ): 1391.62 (331.13)] than in the asymptomatic group [ (σ): 1110.09 (292.54)] (Fig. 1D). For example, the median TNF-α dosage was higher (p = 0.0029) in HAM subjects [median (IQR 25%–75%): 20.36 (17.60–23.96)] than in asymptomatic subjects [median (IQR 25%–75%): 15.76 (12.83–18.86)] (Fig. 1E).
Sex and the rs2232365 polymorphism were not associated with the clinical profiles of HTLV-1 infection
Regarding gender, we observed that there were more female individuals in both the general infection and clinical groups, with the subtle differences in frequency not being significant between the analyzed groups (Table 1).
Sex and Frequencies of Polymorphism rs2232365 in Individuals Infected with Human T Lymphotropic Virus 1
HAM, HTLV-1-associated myelopathy; HTLV-1, human T lymphotropic virus 1.
The frequency of the rs2232365 polymorphism was in Hardy–Weinberg equilibrium in both the general infection (p: 0.1001), asymptomatic (p: 0.2261), and HAM (p: 0.2631) groups. In females, the AA genotype was the most frequent, both in the general infection and different clinical groups, and this was reflected in the allele frequency of the A allele as well (Table 1). However, for males, the G allele was the most frequent in both the general infection and different clinical groups (Table 1). In the polymorphism frequency analysis, none of the subtle differences were significant (Table 1).
The rs2232365 polymorphism, but not sex, was associated with the predictive variables of clinical profiles of HTLV-1 infection
We evaluated the likely associations of sex and the rs2232365 polymorphism with the predictive factors of different clinical profiles of HTLV-1 infection. Regarding sex, we did not observe significant differences between the means or medians of the predictive factors when compared between the sexes (Fig. 2A, C, E). Our analyses showed that both the log 10 values of the PVL (p: 0.7903; AA: median: 2.80; 95% CI: 0.63–2.99); (AG: median: 1.6; 95% CI: 0.41–3.08); (GG: median: 2.24; 95% CI: 1.62–2.40) and concentrations of CD4+ T lymphocytes [p: 0.0712; (AA: median: 1666; 95% CI: 1404–1756); (AG: median: 10526; 95% CI: 943–1147); (GG: median:1177; 95% CI: 1051–1562)] exhibited no significant differences when comparing the genotypes only among females.

Association of predictive factors for sex and polymorphism rs2232365:
However, regarding polymorphism, we observed that the median log 10 value of the PVL was higher (p: 0.0416) in carriers of the GG genotype [median (IQR 25%–75%): 3.12 (2.36–3.49)] than in carriers of the AA genotype [median (IQR 25%–75%): 1.92 (0.09–2.39)] and heterozygotes [median (IQR 25%–75%): 1.56 (0.41–3.08)] (Fig. 2B).
Similarly, carriers of the GG genotype had a higher mean concentration of CD4+ T lymphocytes than carriers of the AA genotype and heterozygotes [p: 0.0232; AA: (σ): 1146.50 (323.24); AG: (σ): 1058.38 (209.39); GG: (σ): 1384.23 (348.94)] (Fig. 2D).
Only the TNF-α dosage was not associated with rs2232365 polymorphism genotypes (Fig. 2F). It is worth noting that in the analysis of the association of polymorphisms with predictive factors, data from hemizygous men were pooled with data from homozygous women. We do not consider that this may have biased our analysis, since sex was not a determining factor in this study.
In addition, we evaluated whether the associations observed between PVL, CD4+ T lymphocyte count, and polymorphism were maintained when we separated patients according to clinical aspects of HTLV-1 infection. The log 10 PVL remained elevated in carriers of the GG genotype regardless of the clinical status of the infection [asymptomatic: 2.87 (2.22–5.53); p: 0.0375], [HAM: 3.26 (3.00–3.45); p: 0.0384] (Fig. 3A, B).
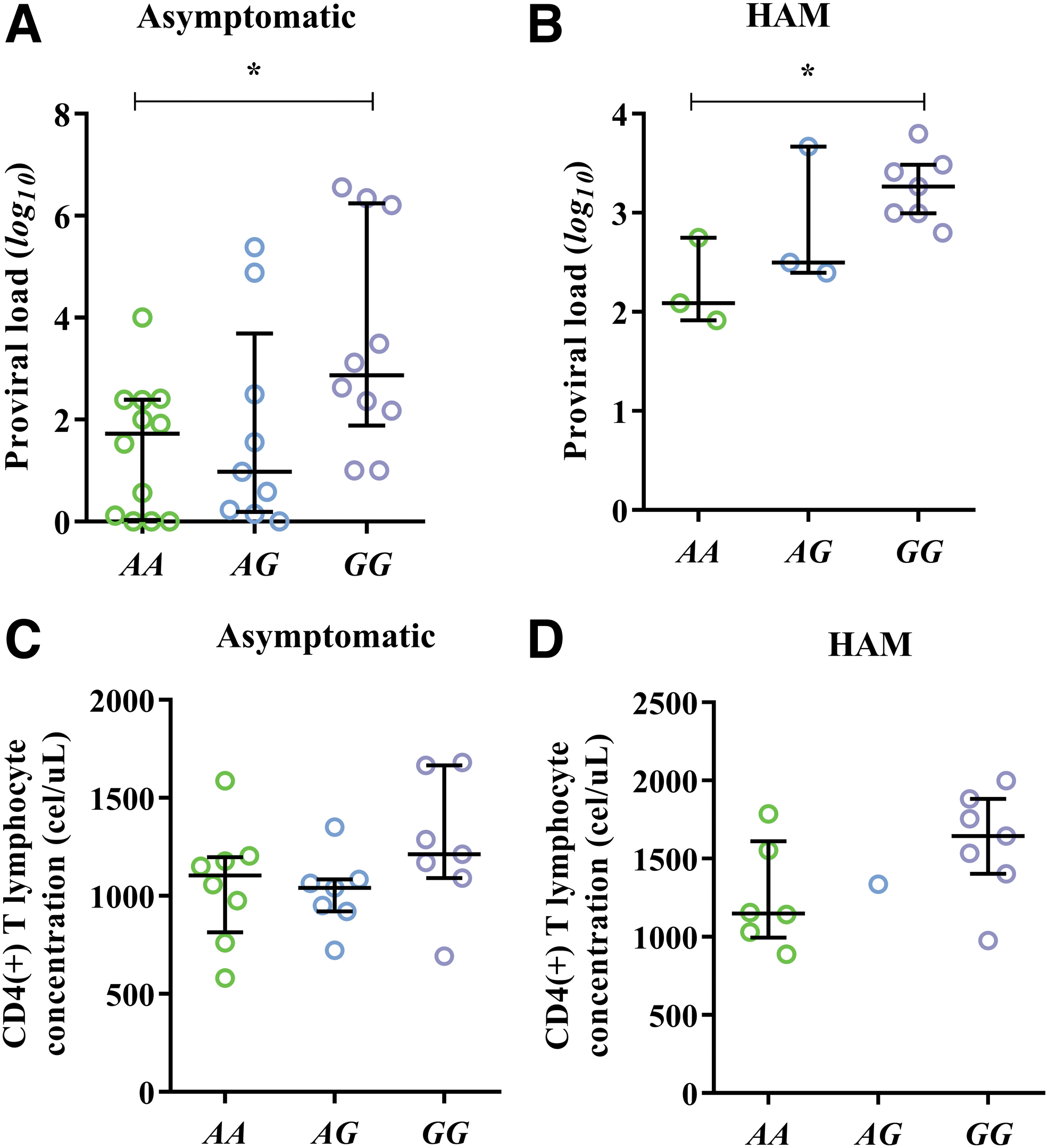
Association of PVL and CD4+ T lymphocyte count to rs2232365 polymorphisms according to clinical status of HTLV-1 infection:
The CD4+ T lymphocyte count did not remain associated with the polymorphism; however, we observed a tendency for carriers of the GG genotype to have an increase in cell counts regardless of the clinical status of the infection [asymptomatic: 1599.57 (341.22); p: 0.2657], [HAM: 1256.87 (342.45); p: 0.1003] (Fig. 3C, D).
Discussion
HTLV-1 is endemic worldwide and is associated with diseases such as HAM. Transmission occurs through sexual intercourse or from mother to child, and sexual transmission occurs mainly from males to females (Martel and Gotuzzo, 2022; Paiva and Casseb, 2014). Considering the gender analysis presented in this study, we did not notice any significant differences among the means or medians of the predictive factors when comparing both sexes, although previous data from our group have suggested that polymorphism rs2232365 might be a protective factor against infection, especially in males (Pereira et al, 2020).
This study demonstrated that the PVLs were higher in the group with HAM, which corroborates the findings from other groups (Nagai et al, 1998). The pathophysiology of HAM is not well understood; nonetheless, previous studies reported that host genetic and immunological factors can be correlated with the disease. PVLs are commonly higher among HAM patients than in asymptomatic individuals, and a high PVL suggests a higher risk of inflammatory diseases (Brunetto et al, 2014; Nagai et al, 2001; Primo et al, 2009). Regarding the presence of the polymorphism in the promoter region of FOXP3, in this study, carriers of the GG genotype had higher PVLs.
Studies suggest that the GG genotype of the rs2232365 polymorphism decreases expression of the FOXP3 gene and causes an immune imbalance, predisposing the individual to develop inflammatory diseases (Song et al, 2012). This proinflammatory imbalance is characteristic of the worsening clinical conditions of HTLV-1 infections, which favor viral proliferation and expressions of genes linked to pathogenesis, which are similarly associated with a failure of immune regulation (Oh et al, 2006). We suggest that the genetic predisposition of the host could thus provide a favorable environment for replication and consequent viral pathogenesis.
Therefore, a series of immunological dysfunctions include the proliferation of CD4+ T lymphocytes and increased migratory capacity of circulating leukocytes. Proliferation of infected T lymphocytes is induced by accessory and regulatory proteins, such as the transcriptional cofactor, Tax, which alters the signaling pathway related to antiapoptosis and cell proliferation (Mohanty and Harhaj, 2020). Regarding the LTCD4+ analysis, the levels of CD4+ T lymphocytes in patients diagnosed with HAM were statistically higher than those found in asymptomatic individuals, which supports the evidence that these cells are related to development of the disease (Goon et al, 2004).
In addition, it has also been reported that HTLV-1 carriers have higher frequencies of CD4+FOXP3+ Tregs (Pinto et al, 2014). Beyond this, this study also demonstrated that carriers of the GG genotype had a higher mean concentration of CD4+ T lymphocytes, as observed in previous studies (Pereira et al, 2017). Again, with a decrease in FOXP3 expression, immunological dysregulation may occur, which would facilitate maintenance of the inflammatory profile that is typically observed in HTLV-1 infections. It is worth noting that the polymorphism has already been associated with other inflammatory conditions, such as psoriasis (Song et al, 2012) and vitiligo (Song et al, 2013).
For TNF-α, our results showed high serum levels of these cytokines. As reported previously, individuals with HAM present high levels of proinflammatory cytokines, such as TNF-α, which allows infiltration of T cells infected with HTLV-1 in neural tissues, contributing to inflammation and tissue damage (Futsch et al, 2018).
Conclusions
In summary, the results suggest that PVLs, lymphocyte concentrations, and TNF-α dosages are predictive factors for the clinical picture of HAM. Allelic variants of the rs2232365 polymorphism of the FOXP3 gene may contribute to pathogenesis of the disease due to its association with some predictive factors. Considering the small sample size, we believe that multicentric studies involving larger groups of persons infected with HTLV-1 from different populations and ethnicities deserve special attention, given the importance of the FOXP3 gene in the homeostasis of the immune response as well as the immunopathological nature of HAM.
Footnotes
Acknowledgment
We thank all patients who agreed to participate in this study.
Authors' Contributions
M.W.S.M., M.A.F.Q., R.N.M.F., and L.M.S.P. performed DNA extraction, molecular analysis, cell quantification and cytokine dosage. C.A.C. and M.S.S. recruited patients and collected samples. M.A.F.Q. performed PVL quantifications. M.W.S.M., A.N.M.R.S., and L.M.S.P. performed the statistical analysis of the data. J.C.M., R.I., A.C.R.V., and A.N.M.R.S. carried out a general review of the article. R.I., A.C.R.V., and A.N.M.R.S. were the creators of the research project.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
National Council for Scientific and Technological Development Grants no. 312979/2018-5 (R.I.), no. 301869/2017-0 (A.C.R.V.), and no. 442522/2019-3 (ACRV Project Grant) and Federal University of Pará (PAPQ/2022).