
Abstract
The kynurenine pathway of tryptophan catabolism can modulate inflammatory responses inducing immunotolerance or immunosuppressive effects. Indoleamine 2,3-dioxygenase (IDO) is the rate-limiting enzyme in this pathway. Early aberrant inflammation is implicated in severe dengue, and herein we investigate and characterize the expression of IDO pathway genes in severe dengue patients. We use a SyBR green-based qPCR to evaluate the leukocyte expression levels of IDO1, IDO2, AhR, TGF-β, ARG1, IFNγ, and IFNα in a dengue patient cohort (n = 51). Twenty-two cases were identified as severe dengue using the WHO case classification (2009) criteria. Principal component analysis (PCA) was employed to examine the relationships of gene expression profiles with disease severity and laboratory markers of clinical severity. We find that two principal components describe most of the variance (65.3%) in the expression patterns of the cohort. Reduced expression of IDO1, TGF-β, and AhR, represented by low Component 2 scores, was significantly associated with disease severity, thrombocytopenia, and leukopenia. Higher expression levels of IDO2, IFNγ, and IFNα positively correlated with Component 1 scores, and were significantly associated with elevated ALT (p = 0.018) and AST (p = 0.017) enzymes. Our results suggest that profiling the baseline expression patterns of the IDO pathway genes may aid in the identification of dengue patients most at risk of severe disease.
Introduction
Dengue fever remains a public health concern in regions where frequent dengue outbreaks cause significant mortality and morbidity. The European Center for Disease Prevention and Control (ECDC) reported 3766153 cases of dengue globally until September 2021 (Communicable Disease, 2022). The mortality rate of dengue infections varies regionwise and remains at its highest (2%) in DENV2 endemic regions. Frequently, coinfections with multiple serotypes account for half of the reported hospitalized severe dengue cases (Guo et al, 2017; Gutierrez-Barbosa et al, 2020).
India recorded a sharp upsurge in dengue incidence from January to December 2022, with 86 reported deaths and 110473 infections (Communicable Disease, 2022). While dengue usually presents as a mild self-limiting illness, in some cases, dysregulated virus-induced inflammation occurs, which results in severe immunopathology leading to fatality. The infection-activated immune response induces cytokines and chemokines that mediate transient functional alterations in the vasculature integrity. This results in bleeding, coagulation disturbances, and thrombocytopenia—the hallmark features of severe disease (Guzman et al, 2016; Pirofski and Casadevall, 2008; WHO, 2011).
The catabolic pathways of amino acids tryptophan and arginine can modulate cell signaling and proliferation during infections. The IDO/KYN pathway consists of two related enzymes, indoleamine 2,3 dioxygenase 1 and 2 (IDO1 and IDO2), that mediate oxidative cleavage of Trp to N-formyl kynurenine and other kynurenines (KYNs) (Ayres, 2020; Jung et al, 2019; Sáez-Cirión and Sereti, 2021; Schmidt and Schultze, 2014).
In contrast, enzymes arginase1 and arginase2 (ARG1 and ARG2) catabolize arginine to L-ornithine, an essential precursor in the biosynthesis of glutamic acid, proline, and polyamines (Cha et al, 2018; Mondanelli et al, 2017). Recent evidence indicates that the elevated catalytical activity of these enzymes skews immune homeostasis to immunosuppression and tolerance. The by-and-end products of these catabolic pathways can also act as immunomodulators (Jones et al, 2015; Kang and Kumanogoh, 2020; Mondanelli et al, 2017).
Among the immune cells, expression levels of IDO1 and ARG1 are highest (∼70%) within the myeloid population, viz. dendritic cells (DCs), macrophages, monocytes, and neutrophils. Lymphocytes account for the rest, with B cells reporting the lowest expression levels at ∼5% (Wirthgen et al, 2018). Type I and II interferons induced during inflammatory conditions are recognized as potent activators of IDO1 expression (Fallarino et al, 2012; Grohmann et al, 2003; Pallotta et al, 2011).
The upregulation of IDO expression in DCs affects a tolerogenic state that mediates a Treg phenotype and limits the effector T cell population (Grohmann and Bronte, 2010; Gutiérrez-Vázquez and Quintana, 2018). Depletion of Trp and generation of KYNs also enable immunosuppression through inhibition of mTOR activity and attenuation of the proinflammatory Th17 signaling (Fallarino et al, 2012; Grohmann et al, 2003). The proinflammatory cytokine TNFα regulates ARG1 expression through IL-4, resulting in increased NO levels essential for pathogen clearance (Cha et al, 2018; Schleicher et al, 2016).
The IDO pathway activity has demonstrated a significant growth inhibitory effect on various pathogens, including Toxoplasma spp., Mycobacterium tuberculosis, and Chlamydia pneumoniae (Blumenthal et al, 2012; Murakami et al, 2013; Ufermann et al, 2019); however, its exact role in viral infections is complex and context dependent, not always correlating to pathogen clearance. In HSV and CMV infections, an upregulation of IDO1 expression correlates with improved viral clearance (Newman et al, 2021; Reinhart et al, 2012; Spekker-Bosker et al, 2019). Adam et al (2004) observed an IFNγ-induced increase in IDO1 activity corresponding to reduced HSV replication in human astrocytoma cells. However, in HIV infections, Li et al. showed that depletion of IDO-expressing pDCs restored the cytolytic T cell activity.
Furthermore, upregulation of IDO1 induced by chronic activation of pDCs led to T cell exhaustion and persistence of HIV DNA reservoirs, despite combination ART therapy (Li et al, 2019). A similar effect of diminished effector T cell function was also observed in EBV infection of monocyte-derived macrophages, but in these infections IL6 and TNFα induced IDO expression instead of IFNγ (Liu et al, 2014).
The precise role of IDO1 and ARG1 in dengue or other flavivirus infections is not as well studied. Few studies have reported an upregulation of IDO1 expression and activity in dengue infections (Becerra et al, 2009; Geisler et al, 2020). A significant increase in KYNs and a decrease in Trp levels were noted in dengue illness, especially in the early phase of infection. A similar observation was also noted in a PBMC culture model of DENV infection, confirming increased IDO activity in dengue infections (Bonezi et al, 2020).
Interestingly, a study by Azevedo et al (2019) demonstrated an association between G2431A IDO1 gene single nucleotide polymorphism (rs3739319) and clinical severity in dengue fever, although the exact mechanisms underlying this association are still unknown. A role for the IDO pathway was also described in Zika infections, using IDO1 −/− mice and 1-MT, a known IDO1 inhibitor. Marim et al (2021) showed that IDO1 blockade mediates neuroprotection in virus-induced pathology.
Independent of the IDO1 coregulation, Type I and II interferons are known to mediate an antiviral effect in dengue infections, particularly when dengue-infected cells are pretreated with these cytokines. Several studies have highlighted the critical role of interferons, especially IFNγ, in dengue infections both directly and indirectly (Diamond et al, 2000; Fonseka et al, 2022; Iyer and Sucila Thangam, 2022; Lee et al, 2022).
Results have revealed a possible association of the levels of this cytokine with disease severity, although a positive correlation is far from established. Virus genotype, phase of infection, and infection status have been shown to differentially regulate the levels of IFNγ in dengue patients. IFNγ is also shown to be a strong inducer of IDO1 expression through the JAK/STAT pathway, and its role in tumor tissues is well demonstrated (Watcharanurak et al, 2014). Nevertheless, the regulatory role of the IDO pathway in severe dengue infections remains unclear.
In this study, we attempt to examine whether pathways regulating Trp and Arg activity modulate the immune responses to dengue infections at an increased risk of clinical severity by evaluating the expression patterns of IDO1 and its coregulators in immune cells.
Materials and Methods
Study design and patients
This retrospective study comprises 51 patients who presented with a history of febrile illnesses to the General Medicine Department of Amrita Institute of Medical Sciences, Kochi, from June 2017 to June 2021. Dengue diagnosis was confirmed either serologically using the dengue NS1 antigen (Panbio Dengue Early ELISA) or anti-Dengue IgM (Panbio Dengue IgM Capture ELISA) or viral RNA PCR test using the Tropical Fever Panel (FTD Diagnostics).
This study was conducted in accordance with the Declaration of Helsinki, 1975, and approved by the Institutional Ethics Committee of Amrita Institute of Medical Sciences, Kochi, Kerala (ECASM-AIMS-2021-381). A study waiver was obtained to investigate the unused deidentified clinical samples and collect the corresponding clinical data from the hospital's electronic medical records (EMRs).
Study procedures
Clinical and laboratory data collection
Detailed demographic, clinical, biochemical, and microbiological data and outcomes of the study subjects were collected from the hospital's EMRs. All confirmed dengue cases were classified according to The WHO Dengue Case Classification 2009 scheme—Dengue without warning sign (DwoWS), dengue with warning sign (DwWS), and Severe Dengue (SD)(WHO, 2011). EDTA and clotted blood samples from study patients and age- and gender-matched healthy subjects (n = 4) were collected, processed, and stored at −80°C immediately upon receipt of the samples.
Serum was investigated for the presence of nonstructural protein 1 (NS1) and dengue-specific IgM and IgG using the ELISA method. Samples that tested positive for NS1 and IgM were considered “acute phase” dengue.
Quantitative real-time polymerase chain reaction
Total RNA was extracted from the buffy coat using the TRI Reagent® (Sigma) per the manufacturer's instructions. The RNA purity and concentration were determined, and cDNA was synthesized from the DNase I (Sigma)-treated total RNA (1 μg) using the Verso cDNA synthesis kit (Thermo Fisher Scientific; AB1453/A). Real-time quantitative gene expression PCR was performed on ABI 7500 fast (Thermo Fisher Scientific) instrument using the Maxima SYBR Green/ROX qPCR Master Mix kit (Thermo Fisher Scientific).
A melt curve analysis was performed after amplification to confirm the presence of a single product peak. Table 1 lists all the gene-specific primers used in this assay. Each target gene expression value was normalized against the housekeeping gene, GAPDH, and relative gene expression (ΔCt) was estimated as the difference between the target gene's cycle threshold (Ct) and the reference gene. All results were analyzed as normalized values of 2−ΔCt (Bustin, 2010).
List of Primers Used in This Study
Statistical data analysis
Principal component analysis (PCA) was performed using the relative gene expression values to determine the noncorrelated “principal components.” Before PCA, variables were log-transformed and normalized to the mean to account for distribution skewness and range of each variable and differences between the clinical phenotypes. PCA results in principal components (PCs) that explain the maximum variability in the data, with the first component accounting for the highest variance and subsequent components explaining the variance, given that it is uncorrelated to the preceding components.
Logistic regression models evaluated the association between PC scores and clinical severity. Pearson's correlation coefficient was determined to evaluate correlations of individual gene expression with continuous clinical laboratory variables. Either one-way ANOVA or the Wilcoxon rank-sum test was applied on continuous variables of normal distribution. Statistical significance was defined as p ≤ 0.05, and all data analysis was performed in the R 4.2.0 (R Core Team, 2022) and RStudio 2022.07.1 using packages prin.comp, biplot, ggplot2, heatmap, ggpubr, and dplyr.
Results
Baseline characteristics of the study cohort
The study cohort included 51 dengue-positive patients admitted to the hospital with fever (≥38°C), myalgia or arthralgia, rash, thrombocytopenia, or hepatomegaly, and a serological or molecular confirmation of dengue diagnosis. The baseline demographics and clinical and laboratory features of the patients in this study are detailed in Table 2.
Baseline Details of the Dengue Study Cohort
The continuous variables are represented as mean (±SD) values, and the categorical variables are reported as numbers (%). The WHO (2009) case classification scheme was employed to categorize the dengue cases based on the “on admission” presentation.
DwoWS, dengue without warning signs; DwWS, dengue with warning signs; SD, severe dengue.
The dengue severity status of the patients was determined using the WHO 2009 case classification criteria. Of the total 51 patients, 14 (27.4%) patients had “dengue without warning signs” (DwoWS), 15 (29.4%) had “dengue with warning signs” (DwWS), and 22 (43.1%) were classified as “severe dengue” (SD). A male preponderance (60.8%) was noted in all three subgroups.
The median age was 54.5 years (IQR 38–78) among the DwoWS group compared with 47 years (IQR 28–72) and 45 years (IQR 32–70) among the DwWS and SD patients, respectively. There were no significant differences among the three clinical case classification groups for most of the evaluated variables in Table 1.
Gene expression in dengue patients and association with severity
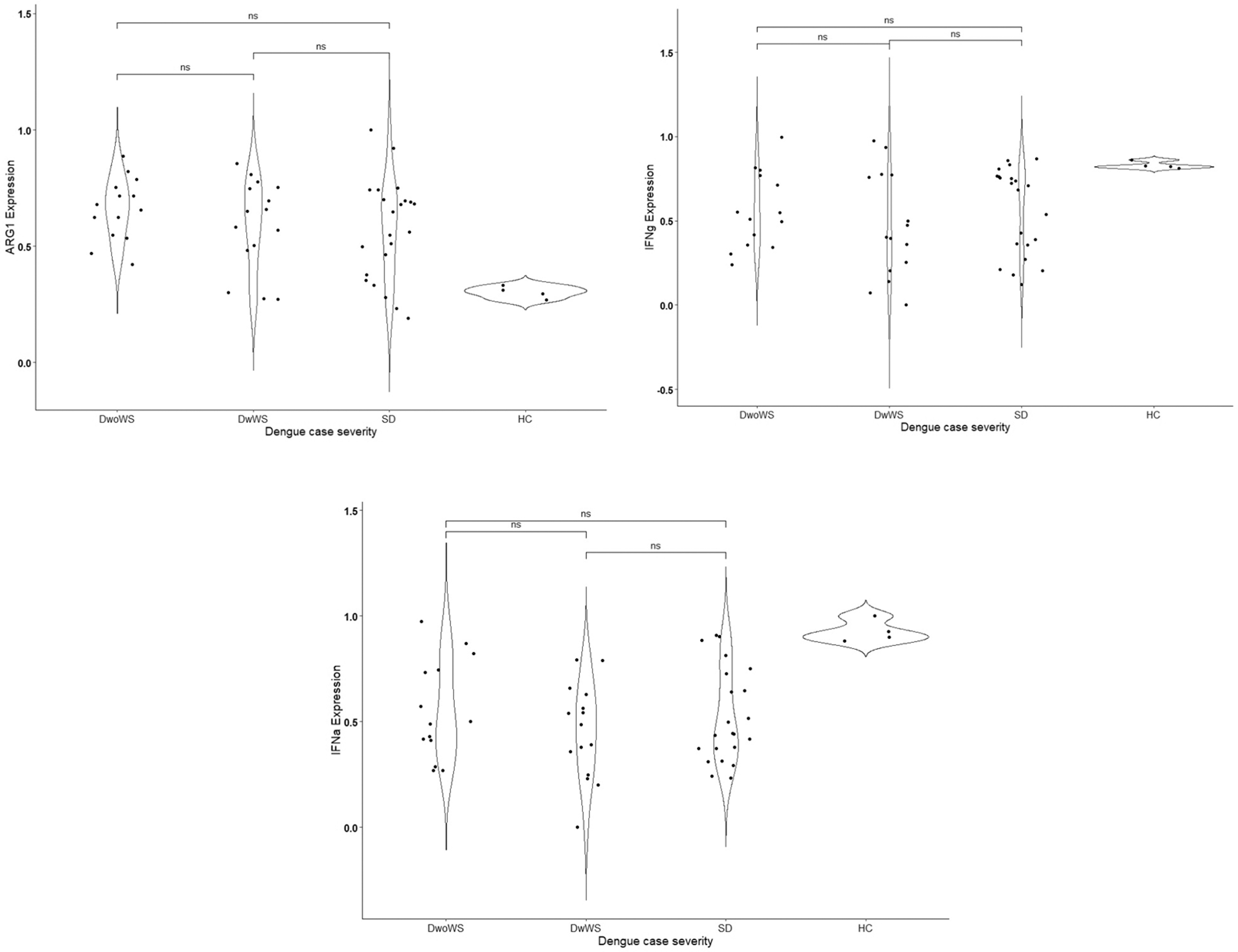
We compared the expression levels of IDO1, IDO2, IFNγ, TGF-β, AhR, ARG1, and IFNα in leukocytes of dengue patients (n = 51) and healthy controls (n = 4). As shown in Figure 1, we observed a significant decrease in the IDO1 expression levels in the leukocytes of SD patients compared with dengue patients with and without warning signs (DwoWS and DwWS).
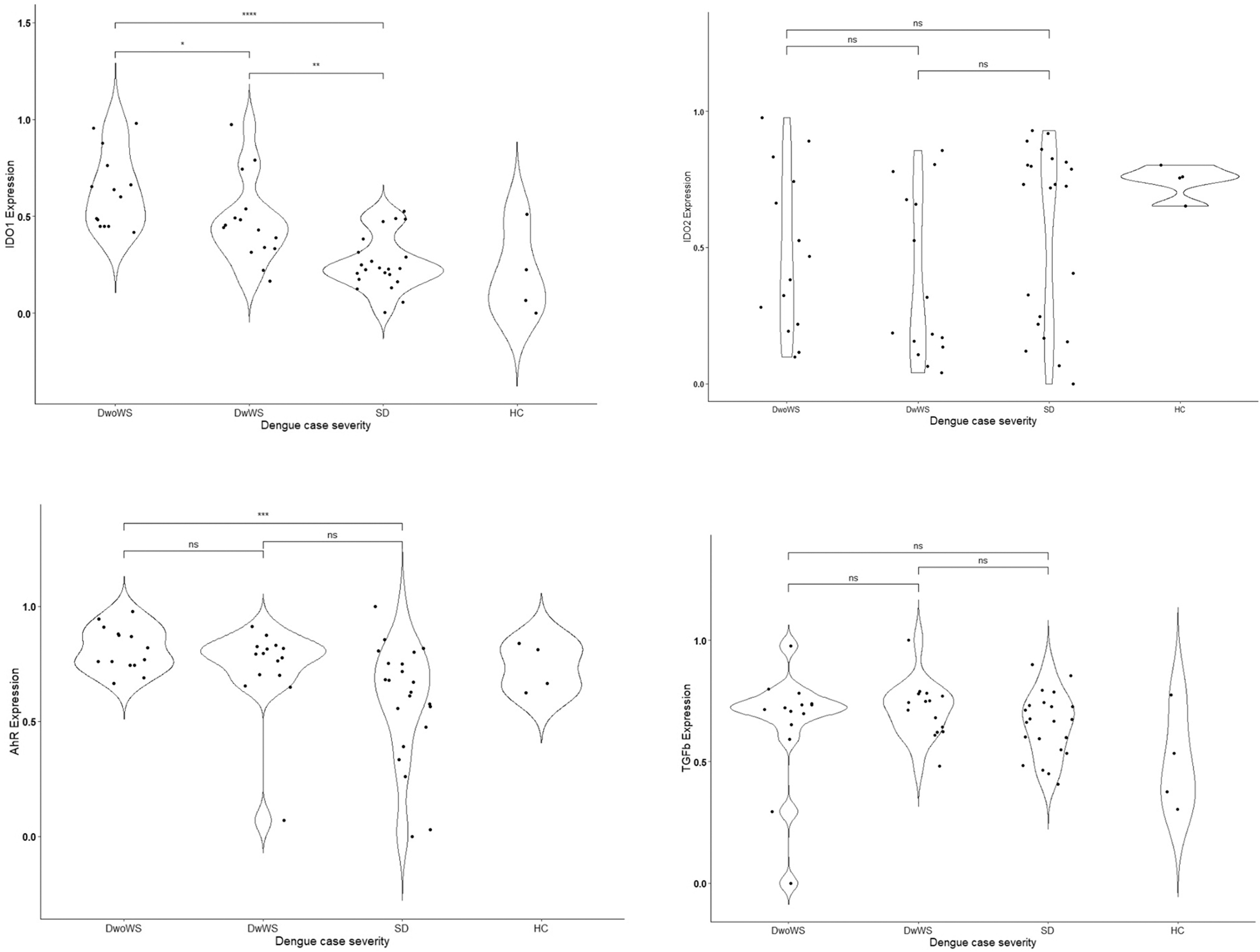
Baseline mean (±SEM) log-transformed and normalized relative gene expression values (2-ΔCt) of IDO1, IDO2, AhR, TGFβ, ARG1, IFNγ, and IFNα in a cohort of 51 dengue cases categorized as DwoWS, DwWS, and SD. One-way ANOVA was performed to evaluate for statistically significant associations between gene expression values and severe dengue cases, while the Kruskal test was performed for multiple comparisons. DwoWS, dengue without warning signs; DwWS, dengue with warning signs; SD, severe dengue.
The mean relative normalized gene expression values of IDO1 observed in the leukocytes of patients without warning signs were 0.62 ± 0.2 compared with 0.25 ± 0.1 in severe dengue, a decrease of 2.5-fold (p < 0.05). A 1.6-fold decrease was also noted in DwWS compared with DwoWS patients (0.38 ± 0.24; p < 0.05).
The expression levels of AhR and TGF-β were also lower in SD patients compared with the other groups. AhR expression decreased ∼1.4-fold in both DwWS and SD patients compared with the DwoWS group (0.57 ± 0.26 or 0.58 ± 0.21 vs. 0.78 ± 0.09; p < 0.05). However, we did not observe statistically significant differences in the expression levels of IDO2, IFNγ, IFNα, and ARG1 between the dengue groups. Nevertheless, IDO2, IFNγ, and IFNα expression levels were upregulated, while ARG1 and TGF-β expression levels were lower in severe dengue.
Next, we used PCA to identify correlated gene expression values across the sample dataset of 51 patients and 4 healthy controls. Most of the variance was accounted for by two components, viz. Comp.1 and Comp.2; Comp.1 was responsible for 41.6% and Comp.2 for 23.7% of the dataset variances—Figure 2A.

PC analysis scores and weightings.
The gene expression loadings within each PC are depicted as a heatmap where IDO2, IFNγ, and IFNα gene expression positively correlated with Comp.1. Comp.2 scores were driven primarily by positive loadings of AhR1, IDO1, TGF-β, and ARG1 and by negative correlation with IFNγ (Fig. 2B). Figure 2C depicts the relationship of the top loadings within each component. As indicated in Figure 2C, Comp.1 domain (top panel) significantly correlated with IDO2 (R = 0.95; p < 0.0001), IFNγ (R = 0.89; p < 0.0001), and IFNα (R = 0.75; p < 0.0001), whereas Comp.2 (bottom panel) correlated with AhR (R = 0.78; p < 0.0001), TGF-β (R = 0.69; p < 0.0001), and IDO1 (R = 0.48; p < 0.002).
PCA and its correlation with dengue severity and clinical variables and outcomes
Next, we evaluated the association of Comp.1 and Comp.2 scores as independent variables with clinical severity. We observed that the Comp.2 score was significantly associated with clinical severity (p < 0.0001). The Comp.2 scores of severe dengue cases were significantly lower (−0.64 ± 0.88) compared with DwWS (−0.22 ± 0.92) and DwoWS (0.66 ± 0.71) (Fig. 3A), indicating that reduced expression of IDO1, AhR1, and TGF-β coupled with increased expression of IFNγ associates with clinical severity. Although not significant, we observed differences in Comp.1 scores among the three groups of dengue cases. While the Comp.1 scores of SD (0.15 ± 0.97) and DwoWS (0.12 ± 0.99) were similar, lower scores were attributed to the DwWS cases (−0.42 ± 0.99).
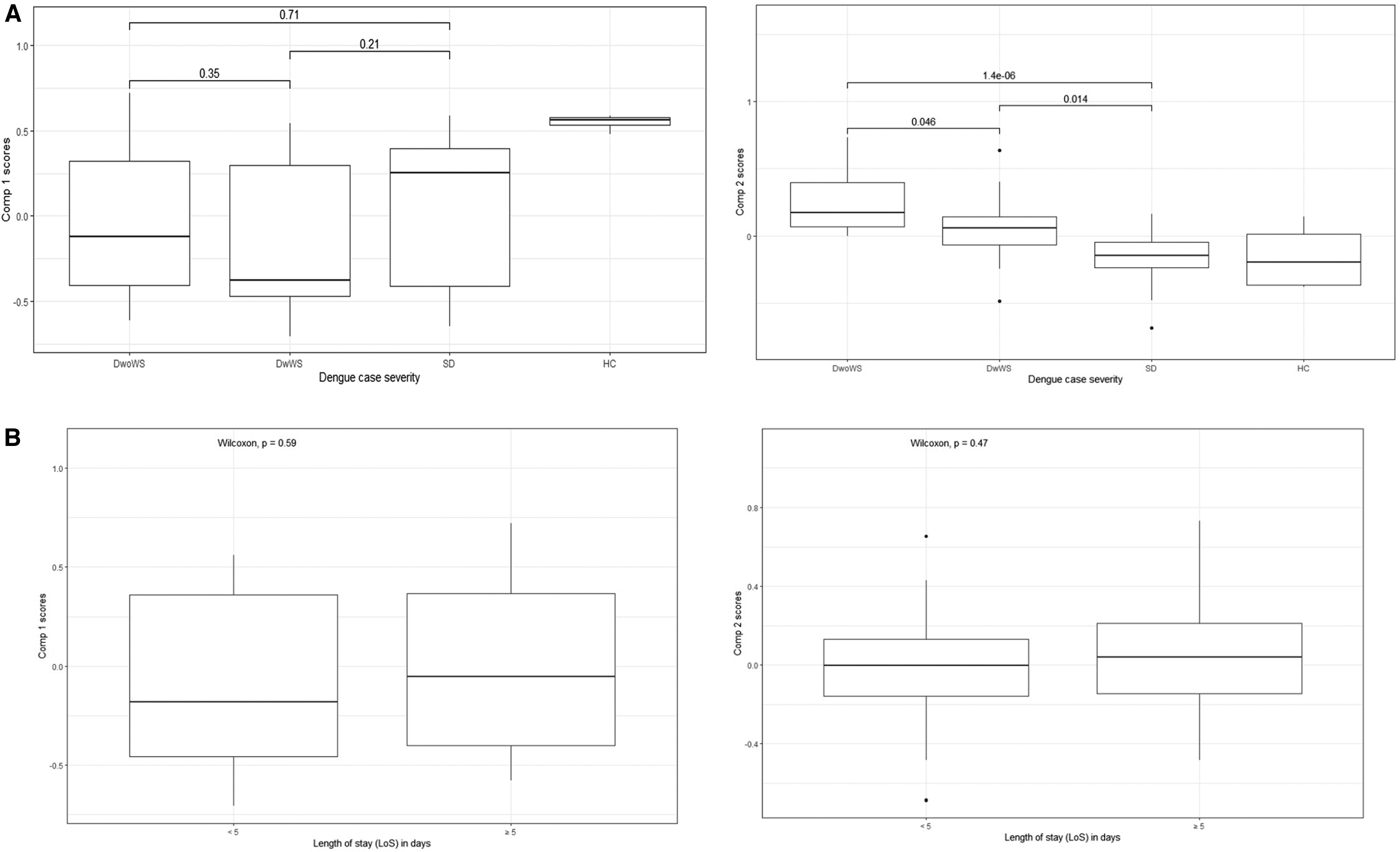
Associations between PC scores and dengue severity and hospital stay.
We also evaluated an association of the PC scores with disease morbidity using hospitalization duration as a surrogate. As shown in Figure 3B, there was no significant association of the hospitalization period with either Comp.1 or Comp.2 scores. Although patients with ≥5 days had higher Comp.1 scores, the average length of hospital stay is a multifactorial outcome that can confound any observation of a direct correlation with gene expression patterns that may not be reflected.
On the contrary, we found that cases with moderate-to-severe thrombocytopenia (platelet counts <50 K/μL) had significantly lower Comp.2 scores than those with >50 K/μL counts (p = 0.038) (Fig. 4B). Similarly, cases with plasma leakage had lower mean ± SD Comp.2 scores than those without any plasma leakage (−0.24 ± 1.2 vs. 0.27 ± 0.65; p = 0.34) (Figure 4A). Unlike Comp.2 scores, Comp.1 scores of the dengue cohort did not differentiate dengue patients with thrombocytopenia and plasma leakage.

Associations of the PC scores with clinical laboratory parameters of

Further exploration of the relationship of Comp.1 and Comp.2 with liver enzyme (AST and ALT) levels, WBC, monocytes, eosinophils, and platelet lymphocyte ratio (PLR)—a known marker of inflammation—is tabulated in Table 3. While the liver enzymes—AST and ALT—positively correlated with Comp.1 scores, Comp.2 scores for these variables showed an inverse relationship.
Correlations Between the Principal Components Scores and Baseline Clinical Laboratory Values
Elevated liver enzymes correlated with increased expression of IDO2, IFNγ, and IFNα and reduced ARG1, while lower expression levels of IDO1, AhR1, and TGF-β of Comp.2 correlated with increased WBCs. Although not significant, a weak positive association was noted between Comp.2 scores and PLR, and an inverse relationship between monocytes and Comp.1 scores.
Baseline gene expression values and dengue illness risk stratification
We next evaluated if the gene expression patterns defined by the PC loadings can predict dengue severity. As shown in Table 4, the association between Comp.2 scores and dengue severity remained significant in gender- and age-adjusted logistic regression analysis; log odds −9.22 ± 2.84 p < 0.05 and log odds −13.89 ± 4.47 p < 0.05, respectively. Similarly, Comp.2 scores were a predictable risk factor of DwWS (log odds −9.63 ± 4.21, p = 0.02), indicating that an increase in Comp.2 scores reduces the risk of severe dengue compared with the nonsevere cases.
Logistic Regression Analysis Adjusting for Age and Gender for Predicting Log Odds of Increased Dengue Severity with Principal Component Scores
A p-value of <0.05 was considered statistically significant.
Discussion
In this study, we investigated the role of the catabolic pathways of Trp and Arg in the pathogenesis of severe dengue. Numerous studies have highlighted the importance of tryptophan catabolism as a critical regulator of peripheral immune responses. Earlier, Becerra et al (2009) reported an upregulation of IDO1 activity, a corresponding decrease in serum Trp levels, and increased serum kynurenines in dengue infections.
Likewise, we also observed a similar significant activation of IDO1 in dengue patients, although these were primarily in nonsevere cases. We report here, probably for the first time, that there is a significant impairment in the expression of IDO1 and AhR1 in severe dengue. Although not statistically significant, IDO2, IFNγ, ARG1, and TGF-β also exhibited differential expression patterns among dengue patients based on their clinical severity.
Further, PCA of the gene expression profiles identified two clusters or “components,” viz. Comp.1 and Comp.2, functionally representing a proinflammatory and immunotolerogenic phenotype, respectively. A careful evaluation of the component loadings revealed a higher Comp.1 score and lower Comp.2 score correlated with severe dengue features such as thrombocytopenia, liver dysfunction, and leukopenia, suggesting a significant impairment of immunotolerance.
Overall, these observations suggest that the decreased expression of IDO1, AhR1, TGF-β, and ARG1, coupled with increased expression levels of IDO2 and IFNγ, correlates with immunopathological features of dengue. These results are consistent with previous findings demonstrating a similar downregulated expression pattern of IDO and ARG pathway genes in infections and other conditions such as cancer and autoimmune disorders associated with excessive inflammation or lack of immunosuppression (Chapman et al, 2020; de Araújo et al, 2020; Dougnac et al, 2007).
The emerging paradigm in immune tolerance emphasizes a central role for the IDO1 signaling and enzymatic mechanisms, AhR1-regulated gene expression, and TGF-β–induced T-regulatory activity. IDO1 influences immune responses by causing “amino acid” deprivation, inhibiting cell and pathogen proliferation, or modulating T cell activation and Treg differentiation by the Trp breakdown metabolites (Chapman et al, 2020; Dougnac et al, 2007; Gutiérrez-Vázquez and Quintana, 2018
Kynurenine, a metabolite of the IDO pathway, acts as a transcription activator of AhR1. Several studies have reported cooperative regulation of IDO1 and AhR1 in the context of fungal infections (de Araújo et al, 2020; de Araújo et al, 2017). In our study, an association of the expression patterns of these three genes in severe dengue patients suggests an underlying attenuation of immunosuppressive or Treg activity, which may explain the severe immunopathology of severe dengue.
A recent study by Alves de Souza et al (2022) suggests a possible regulatory role of the IDO/kynurenine pathway in controlling the inflammation-driven pathology in chronic chikungunya infections. IDO2 expression, on the contrary, is less studied in the context of dengue infections. Available evidence from studies evaluating its role in autoimmune conditions and cancer has shown that IDO2 mediates proinflammatory response in autoimmune conditions distinct from IDO1 (Merlo and Mandik-Nayak, 2016
Not surprisingly, in our cohort, IDO2 expression positively correlated with a higher Comp.1 score associated with dengue severity. Arginase, on the contrary, is known to exert immunoregulatory effects in myeloid cells (Grzywa et al, 2020). Upregulation of ARG1 is associated with TGF-β production, and macrophages that are Arg+ are shown to suppress antigen-specific T cell proliferation and limit Th17 differentiation (de Araújo et al, 2017).
Although the role of ARG1 in dengue infection is not well characterized, results from our pilot study suggest that reduced expression of ARG1 is likely associated with increased immune activation and clinical severity. This seems to support a similar finding by Cortes-Selva et al (2018), wherein they have shown that ArG1 suppressed intestinal inflammation in acute schistosomiasis.
Overall, these findings of the gene expression signatures may explain, to some extent, the underlying molecular pathology associated with dengue disease severity. Nevertheless, this study did have a few salient limitations. The small study sample size significantly limited the power of the study findings. Demonstration of the correlation of gene expression with peripheral cytokine levels would have validated the study findings.
Alternatively, characterization of the immune cell-specific gene expression patterns would have facilitated a better understanding of virus-induced inflammation that leads to immunopathogenesis. Finally, as a single-center tertiary care hospital, the possibility of a selection bias in the cohort under investigation remains.
Conclusions
In this study, we noted the dysregulation in Trp and Arg catabolic pathways in severe dengue that may mediate a phenotype of reduced Treg activity and excessive inflammation. These findings highlight the importance of amino acid catabolic pathways as potential immunomodulatory targets and provide new opportunities for therapeutic interventions for severe dengue.
Footnotes
Acknowledgments
The authors thank Merlin Moni MD, Dipu T.S., MD, and Vidya Menon of Dept of Gen Medicine, Amrita Institute of Medical Sciences, who helped and guided them with dengue clinical case classification.
Authors' Contributions
V.M., D.L., and S.J. contributed to conceptualization and writing—review & editing; S.J., A.K., R.J., and J.O. assisted with methodology and investigation; V.M. and S.J. performed formal analysis; S.J. contributed to writing—original draft preparation; V.M. and D.L performed supervision. All authors have read and agreed to the published version of the article.
Informed Consent Statement
Patient consent was waived as the study involved unused, deidentified retrospective clinical samples.
Author Disclosure Statement
The authors declare no conflict of interest.
Funding Information
This research received no external funding.