
Abstract
Toll-like receptors (TLRs) play a crucial role in the innate immune response to pathogens, and TLR3 could recognize and control the herpesvirus. We studied the effect of TLR3 polymorphisms on the risk of Kaposi's sarcoma-associated herpesvirus (KSHV) infection. A cross-sectional study was performed among human immunodeficiency virus (HIV)-infected individuals in Xinjiang, a KSHV-endemic region of China. The frequencies of nine single-nucleotide polymorphisms (SNPs) in TLR3 in 370 KSHV-infected patients and 558 controls, and their impact on plasma IFN-γ levels, were compared. The effect of TLR3 SNPs on the KSHV viral load in KSHV-infected subjects was also assessed. The minor allelic variant at rs13126816 was more common among KSHV-seronegative than KSHV-infected individuals. Two TLR3 SNPs (rs13126816 and rs3775291) showed a protective effect against KSHV infection (rs13126816: odds ratio [OR]dominant = 0.66, 95% confidence interval [CI]: 0.50–0.87; ORoverdominant = 0.65, 95% CI: 0.49–0.87; rs3775291: ORdominant = 0.76, 95% CI: 0.58–0.99; ORoverdominant = 0.75, 95% CI: 0.57–0.98). These associations were stronger in the Uyghur compared with the Han population. The haplotype, CGAC, significantly correlated with the risk of KSHV infection (OR = 0.72, p = 0.029). KSHV-infected individuals with homozygous rs13126816 AA genotypes had a lower KSHV viral load (aOR = 0.14; p = 0.038). However, no association was observed between TLR3 SNPs and plasma levels of IFN-γ. Genetic variants in TLR3 reduce the risk of KSHV infection and affect KSHV reactivation among HIV-infected individuals, especially in the Uyghur population.
Introduction
Kaposi's sarcoma-associated herpesvirus (KSHV), also known as human herpesvirus 8 (HHV-8), is the causative agent of Kaposi's sarcoma (KS), the most common neoplasm in human immunodeficiency virus (HIV)-infected patients (Chang et al., 1994). Of note, KS was reported to be 20,000 times more frequent in patients with acquired immunodeficiency syndrome (AIDS) than in the general population in Western countries (Cesarman et al., 2019), with AIDS-KS being the major cause of death in ∼50% of patients with AIDS (Coghill et al., 2015; Engels et al., 2008).
Regardless of the wide application of combined antiretroviral therapy (cART), AIDS-KS continues to afflict patients infected with HIV (Robey and Bower, 2015; Silverberg et al., 2011), posing a great challenge to the 2022–2030 global health sector strategy of the World Health Organization (WHO) on HIV. Epidemiological data have indicated considerable variations in KSHV infections across populations and geographic regions (Cesarman et al., 2019).
The Xinjiang province in China is an endemic area for KSHV infection, with a prevalence of 48.9% among HIV-infected Uyghurs (Liu et al., 2017). Currently, susceptibility to KSHV infection remains poorly understood; however, the disproportionately higher incidence of KSHV infections in immunodeficient patients than in individuals with an uncompromised immune system points to the crucial role of the immune system.
KSHV can establish a delicate balance between activation and suppression of the immune response to establish its lifelong latency in the host (Damania, 2004; Rezaee et al., 2006). Following viral entry or reactivation and recognition through pattern recognition receptors (PRRs), the host builds an immune response to KSHV (Uppal et al., 2018).
Toll-like receptors (TLRs) are a family of conserved receptors belonging to PRRs (Akira and Sato, 2003). Activated TLRs generally lead to the induction of production of interferon, which is considered to protect against KSHV infection (Kawai and Akira, 2010). Among the 10 known TLRs, TLR3 is unique, in that the initial signaling cascade begins with a noncanonical pathway, with binding of TIR domain-containing adapter-inducing interferon-β (TRIF; Patra et al., 2020) to the TIR binding domains within the intracellular domains of TLR3 (Yamamoto et al., 2003).
TLR3 mediates various immune inflammatory responses through production of TRIF-activated nuclear factor kappa B (NF-κB) and regulation of the release of various inflammatory factors, such as cytokines (Akira and Takeda, 2004; Alexopoulou et al., 2001). TLR3 recognizes poly (I: C), a synthetic double-stranded RNA analog, and viral dsRNA, which is presumably formed during viral infection. In the presence of inflammatory cytokines, poly I: C has been reported to induce the production of natural killer (NK) cell-secreted interferon-γ (IFN-γ), which is essential for intrinsic immunity against viral infections.
Polymorphisms in TLR3 might alter its expression, thus affecting the function and efficacy of signal transduction, leading to an altered immune response (Talaat et al., 2022). Recent studies have identified the role of TLR3 and its polymorphism in the recognition and control of herpesviruses (Skevaki et al., 2015; Svensson et al., 2012; Thompson et al., 2011; Zhang et al., 2007). For instance, TLR3 deficiency in humans has been associated with the uncontrolled spread of herpes simplex virus 1 (HSV-1) and HSV-1 encephalitis (Zhang et al., 2007).
Genetic variations in TLR3 might affect susceptibility to HSV-2 infection (Svensson et al., 2012). However, the mechanism through which TLR3 modulates KSHV infection remains unclear. Most relevant studies have been conducted at the cellular and in vivo levels (Gargano et al., 2009; Gregory et al., 2009; West and Damania, 2008), whereas studies at the population level are lacking.
In this study, we investigated the relationship between TLR3 genetic variants and KSHV infection among HIV-infected individuals in Xinjiang, China, and evaluated the effect of single-nucleotide polymorphisms (SNPs) of TLR3 on the plasma level of IFN-γ. Our findings will contribute to an enhanced awareness of the factors affecting KSHV infection among high-risk populations and will benefit prevention of KSHV in countries facing similar epidemiologic situations.
Materials and Methods
Study design and sample collection
This cross-sectional study was conducted in the Kazakh Autonomous Prefecture of Ili, Xinjiang, China. From May to December 2019, HIV-infected individuals over 18 years of age who received regular antiviral treatment were consecutively recruited in our study. The HIV-positive status of all participants was confirmed by the Centers for Disease Control and Prevention (CDC). Participants were interviewed in person by trained staff from local health facilities.
A standard questionnaire was used to collect information on demographic characteristics and information related to HIV infection. All recruited individuals agreed to their participation in the study and provided blood samples. This study was approved by the research ethics review committee of Fudan University (Shanghai, China; Approval No. IRB#03-0506).
Blood collection
Professional nurses collected blood samples using sterile disposable needles and ethylenediaminetetraacetic acid (EDTA) tubes. Plasma was separated by centrifugation and stored at −80°C for serological testing. All specimens were anonymized and coded using unique identification numbers.
Laboratory testing
KSHV serology
Antibodies against KSHV were detected using an enzyme-linked immunosorbent assay (ELISA) that employs the most immunogenic lytic antigens, ORF65 and ORF73, and latent antigen, K8.1. When used together, serological assays against KSHV lytic and latent antigens show the best combination of sensitivity (89.1%) and specificity (94.9%; Tedeschi et al., 2002). The ORF65, K8.1, and ORF73 coding sequences were recombined into the pQE-80L vector to express the respective proteins in competent Escherichia coli cells, followed by purification of the three proteins using a nickel column, as antigens.
Purified recombinant proteins were diluted to a concentration of 1 ng/μL in 0.05 M carbonate (pH 9), plated with 50 μL of antigenic protein dilution onto microtiter plates, and incubated at 4°C overnight. Plates were then blocked with 250 μL of phosphate-buffered saline (PBS; containing 1% goat serum and 5% powdered milk) at 37°C for 1 h. Plasma samples were diluted 1:100 in the above PBS and incubated at 37℃ for 1 h, followed by incubation with a secondary antibody (1:2000, alkaline phosphatase-labeled sheep anti-human IgG) at 37°C for 1 h.
Subsequently, 1 mg/mL dilute P-NPP with chromogenic buffer was added and incubated for 30 min at 37°C, and 3 M NaOH was used to stop the chromogenic reaction (blue color changed to yellow). The absorbance value of each plasma sample was measured at 405 nm using a microplate reader, and the average optical density (OD) value of the negative control sample plus twice the standard deviation was used as the cutoff value. Participants who were positive for any of the three KSHV antibodies were considered KSHV seropositive.
HIV and KSHV viral load
The HIV viral load was quantified using the Cobas AmpliPrep Cobas TaqMan HIV-1 Test, v2.0 (Roche). HIV-I RNA was extracted from plasma and reverse transcribed into cDNA for detection. The real-time polymerase chain reaction (PCR) was used to amplify the gag region of HIV-1, and samples were quantified using the internal standard method, with a linear range of 20–107 copies/mL.
KSHV viral loads were detected using real-time fluorescence quantitative PCR (qPCR). Briefly, DNA was extracted from leukocytes, according to the manufacturer's instructions. Primer pairs used were KS-1 and -2 (Fwd, 5′-AGCCGAAAGGATTCCACCAT-3′; Rev, 5′- TCCGTGTTGTCTACGTCCAG-3′), amplifying the KS330Bam233 region of ORF26 (a viral capsid protein).
Each 20-μL PCR contained 10 μL of SYBR 2 × mix (NO ROX), 2 μL of DNA template, 7 μL of ddH2O, and 0.5 μL of each primer. The PCR conditions were 95°C for 3 min, followed by 40 cycles at 95°C for 5 s, 55°C for 30 s, 72°C for 30 s, and a final step at 72°C for 5 min. A cutoff value of ≥1000 copies/μL was considered positive for KSHV DNA (Supplementary Fig. S1).
SNP genotyping
DNA was extracted using the TIANGEN® blood genomic DNA extraction kit DP318–03 (centrifugal column type). DNA samples were stored at −80°C before genotyping. Of note, eligible TLR3 SNPs were selected based on studies suggesting a close correlation of these SNPs with viral infections. Eventually, nine TLR3 SNPs (rs1879026, rs3775290, rs3775291, rs3775296, rs5743312, rs5743313, rs5743314, rs13126816, and rs76713360) were included. Information on these SNPs is provided in Supplementary Table S1.
SNPs were detected through multiplex PCR amplification using the FastTarget™ system. Sequencing libraries were generated using the FastTarget® Custom Panel, following the manufacturer's recommendations, with index codes being added to each sample. The fragment length distribution of the library was verified using the Agilent 2100 bioanalyzer. Following accurate quantification of the molar concentration of the library, FastQ data were obtained using the Illumina HiSeq platform for high-throughput sequencing using the 2 × 150-bp double-terminal sequencing mode.
Primer sequences used are listed in Supplementary Table S2.
Detection of levels of IFN-γ
The levels of plasma IFN-γ were determined using a commercially available ELISA kit (ml077386, Mlbio). Absorbance was measured using a microplate reader at a wavelength of 450 nm, followed by calculation of the concentration of each sample. The standard provided by the manufacturer was plotted with a six-parameter fit, and the concentration of each sample was determined based on this standard curve. Each reaction plate contained one negative, one positive, and one blank control.
Statistical analysis
Both the laboratory results and questionnaires were double entered and managed using EpiData 3.1. All data were subsequently transferred to R 3.5.2 and Haploview 4.2 for further statistical analysis. Comparisons of the distribution of characteristics between different groups were made using the Mann–Whitney U test for continuous variables (each when adequate) or Pearson's χ 2 test and Fisher's exact test for categorical variables. Allele frequencies in different populations were compared using the χ 2 test.
Genotype frequencies in different genetic patterns (codominant, dominant, recessive, and overdominant models) were compared using the SNPassoc package. The Hardy–Weinberg (HWE) test, minimum allele frequency (MAF) test, linkage disequilibrium (LD), and haplotype reconstruction with association tests were performed using Haploview 4.2.
Logistic regression was used for correlation analysis to explore the relationship between TLR3 polymorphisms and KSHV infection. General linear regression analysis was used to explore the relationship between TLR3 polymorphism and levels of IFN-γ. These models were adjusted using important covariables. Both odds ratios (ORs) and 95% confidence intervals (CIs) were calculated for all regression coefficients. Differences were considered statistically significant at p < 0.05.
Results
Characteristics of study participants
We recruited a total of 928 HIV-positive individuals for the present study. The demographic and HIV infection-related characteristics of participants are described in Table 1. Briefly, 370 individuals were KSHV seropositive, yielding a seroprevalence of 39.87%. We found that compared with KSHV-seronegative participants, individuals infected with KSHV had significantly lower CD4+ T cell counts (582.46 ± 304.19 vs. 523.18 ± 249.40, p = 0.002).
Characteristics of All Study Participants
AIDS, acquired immunodeficiency syndrome; HIV, human immunodeficiency virus; KSHV, Kaposi's sarcoma-associated herpesvirus.
However, we did not detect any significant differences between the two groups in terms of HIV stage, HIV viral load, and duration of HIV infection.
Rs13126816 and rs3775291 were associated with a reduced risk of KSHV infection
To explore the association between TLR3 polymorphisms and the risk of KSHV infection, we compared the frequency of nine different TLR3 SNPs among 370 KSHV-infected and 558 KSHV-seronegative individuals. The HWE test showed that our recruited participants were a representative population sample (PHWE > 0.05; MAF >0.05). We found that two of the nine SNPs correlated with the risk of KSHV infection among participants in Xinjiang.
We further observed that allele substitution with adenine (A) instead of guanine (G) at rs13126816 in an intron region of TLR3 was correlated with a reduced risk of KSHV infection. We detected the A allele variant in 21.21% of KSHV-seronegative individuals, whereas its frequency in KSHV-infected individuals was 13.45% (p ˂ 0.001; Table 2). Moreover, we identified that 3.43% and 35.56% of KSHV-seronegative individuals were homozygous and heterozygous for the A allele, respectively, whereas only 3.26% and 26.36% of KSHV-infected individuals had homozygous and heterozygous genotypes, respectively (p = 0.011; Table 3).
Allele Frequencies in Kaposi's Sarcoma-Associated Herpesvirus-Infected and -Seronegative Individuals for Nine Single-Nucleotide Polymorphisms in the TLR3 Gene
Bold type indicates SNPs that are associated with a reduced incidence of KSHV infection.
SNP, single-nucleotide polymorphism.
Genotype Frequencies in Kaposi's Sarcoma-Associated Herpesvirus-Infected and Kaposi's Sarcoma-Associated Herpesvirus-Seronegative Individuals of Different Populations for rs13126816 and rs3775291
Interestingly, we observed that rs13126816 significantly decreased the risk of KSHV infection in the dominant (GA + AA vs. GG genotype) and overdominant (GA vs. GG + AA genotype) models (ORdominant = 0.66, 95% CI: 0.50–0.87; ORoverdominant = 0.65, 95% CI: 0.49–0.87; Table 3).
Similar to rs13126816, the homozygous variant of the minor allele at rs3775291 was correlated with a reduced risk of KSHV infection. The SNP, rs3775291, is a missense mutation that leads to the replacement of leucine with phenylalanine at amino acid position 412. We accordingly found that participants with rs3775291 under the dominant and overdominant models were at a lower risk of infection (ORdominant = 0.76, 95% CI: 0.58–0.99; ORoverdominant = 0.75, 95% CI: 0.57–0.98).
Our previous study showed that compared with other ethnicities, KSHV/KS was more frequently detected in the Uyghur ethnic group (Liu et al., 2017). Thus, we further compared the association of genotype frequencies and KSHV infection across different ethnicities. We found that both SNPs, rs13126816 and rs3775291, were associated with KSHV infection in the Uyghur population.
The frequencies of the GG, GA, and AA genotypes of rs13126816 were 59.12%, 37.29%, and 3.59% in KSHV-uninfected individuals, compared with 69.80%, 26.27%, and 3.92% in KSHV-infected patients, respectively. We also noticed that compared with KSHV-uninfected individuals, the rs13126816 genotype significantly decreased the risk of KSHV infection under the dominant (OR = 0.63, 95% CI: 0.45–0.88, p = 0.006) and overdominant (OR = 0.60, 95% CI: 0.42–0.85, p = 0.004) models.
Similarly, we detected the rs3775291 genotype CT more frequently among KSHV-seronegative individuals (41.67% vs. 32.16%). These results showed that the CT and CC-TT genotypes also decreased the risk of KSHV infection in the overdominant model (OR = 0.66, 95% CI: 0.47–0.93, p = 0.016; Table 3). Although we did not find any significant association among the Han population, the genotypes, especially the heterozygous genotypes, of these two TLR3 SNPs showed a protective effect against KSHV infection among the Uyghur population.
We further performed linkage disequilibrium and haplotype analysis among the Uyghur population, with the nine selected TLR3 SNPs that reflected close linkage (Supplementary Fig. S2) being divided into two haplotype blocks (Supplementary Fig. S3). Among them, block2 consisted of rs5743313, rs5743314, rs3775291, and rs3775290.
We observed that the CGAC combination significantly correlated with KSHV infection in this group (OR = 0.72, p = 0.029). This combination was found in 22.0% of KSHV-seronegative individuals compared with 17.0% of KSHV-infected individuals (Table 4).
Haplotype Frequencies in Kaposi's Sarcoma-Associated Herpesvirus-Infected and -Seronegative Individuals for Nine Single-Nucleotide Polymorphisms in the TLR3 Gene Among the Uyghur Population
Bold type indicates haplotypes that are associated with KSHV infection.
Association between TLR3 haplotypes and KSHV viral load
We also assessed the impact of rs13126816 and rs3775291 on the viral load in the KSHV-seropositive population. After adjusting for sex, age, and HIV stage, we determined that the AA genotype of rs13126816 in the infected population correlated with a low KSHV viral load.
In addition, we observed that KSHV-infected individuals with all unfavorable alleles (AA haplotype) had increased odds of a low KSHV viral load (aOR = 0.14; p = 0.038). Strangely, this association disappeared when we divided the population according to the region (Fig. 1).
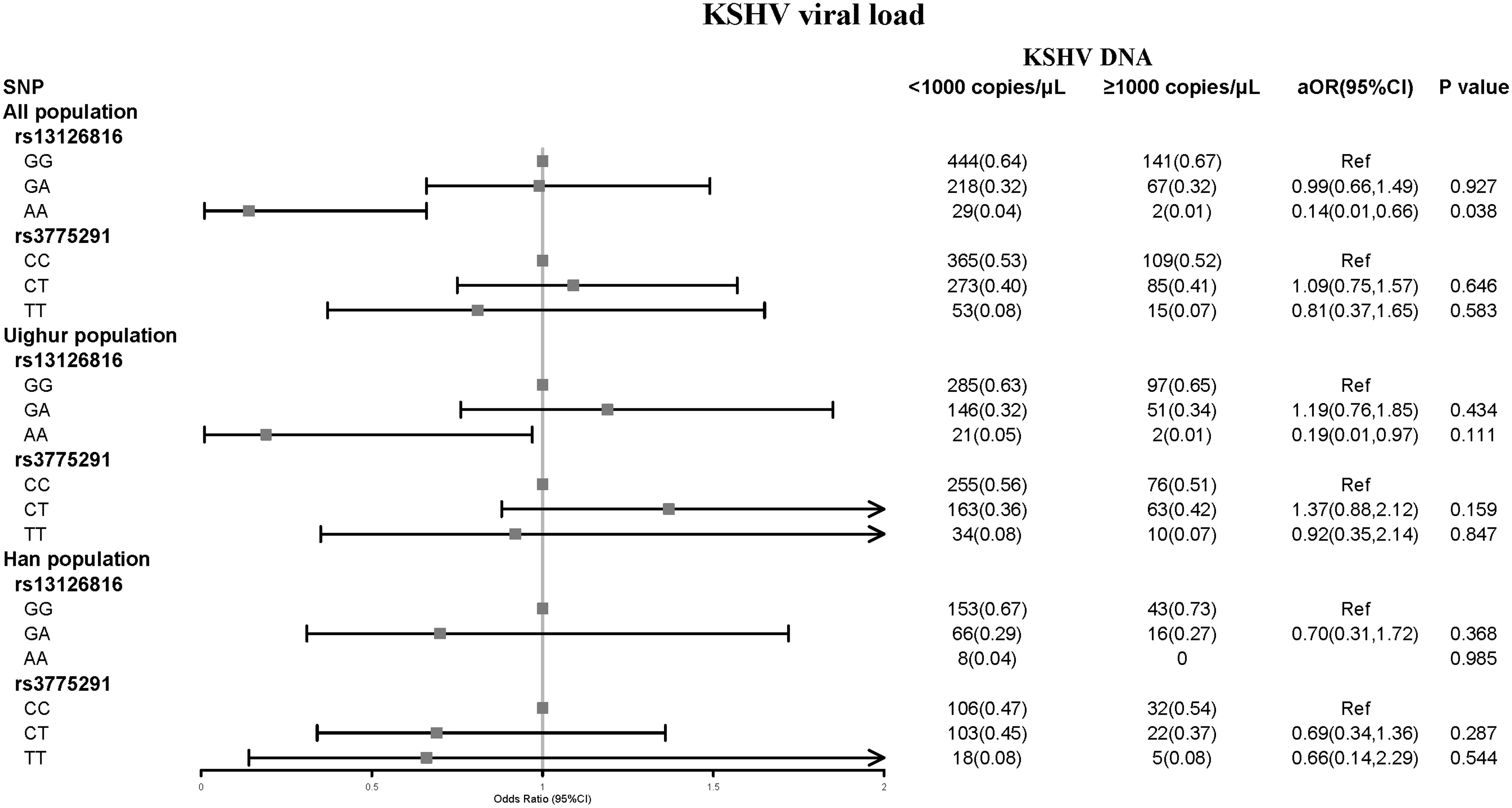
Relationships between two SNP genotypes of TLR3 (rs13126816 and rs3775291) and KSHV viral load in KSHV/HIV patients. p-Values were calculated by logistic regression analysis adjusted for the most important characteristics. 95% CI, 95% confidence interval; AIDS, acquired immunodeficiency syndrome; aOR, adjusted odds ratio; HIV, human immunodeficiency virus; KSHV, Kaposi's sarcoma-associated herpesvirus; SNP, single-nucleotide polymorphism.
Correlation between TLR3 polymorphisms and levels of IFN-γ
We measured the levels of IFN-γ in KSHV-infected individuals and assessed their association with rs13126816 and rs3775291 in TLR3 to explore the possible function of these SNPs in immune responses. We found that the mean plasma level of IFN-γ in infected individuals was 551.89 pg/mL. However, we did not detect any notable differences in levels of IFN-γ between participants (data not shown).
Discussion
It has been well established that susceptibility to KSHV infection is caused by virus–host–environment interactions (Sallah et al., 2020). Despite the chance of exposure, the heterogeneity of KSHV prevalence has suggested the important impact of host genetics, including factors such as HLA (Zhang et al., 2022), MBL2 (de Morais et al., 2018), and EPHA2 (Blumenthal et al., 2018). Nevertheless, research on the impact of genetic predisposition on KSHV infection is lacking, especially at the population level.
TLRs, one of the most important families of PRRs, can rapidly recognize evolutionarily conserved structures on invading microorganisms. By binding to these patterns, TLRs trigger several antiviral responses, playing a key role in the first line of defense against KSHV and in promoting adaptive immune responses (Skevaki et al., 2015). TLR3 recognizes poly (I: C), a synthetic double-stranded RNA analog, and viral dsRNA, which is presumably formed during viral infection.
Therefore, delineating the role and effects of TLR3 variants on KSHV susceptibility is essential. We conducted this study to elucidate the association between TLR3 polymorphisms and the risk of KSHV infection in a Xinjiang HIV-positive population. We found that TLR3 SNPs, rs13126816 and rs3775291, correlated with a reduced risk of KSHV infection, with the homozygous genotype of rs13126816 (AA) potentially affecting the KSHV viral load in infected individuals.
Among all recruited HIV-infected individuals, those coinfected with KSHV had a relatively low CD4+ T cell count, indicating that the risk of KSHV infection in HIV-infected individuals is associated with CD4+ T cell deficiency.
We also observed that SNP variations in TLR3 affect susceptibility to KSHV. For instance, both homozygous and heterozygous genotypes for the A allele at rs13126816 and the CT genotype at rs3775291 showed a protective effect against KSHV infection. These effects became even more evident in the Uyghur population. The rs13126816 polymorphism is located in an intron and might not be a true variant affecting infection; however, we cannot rule out the functional effect of introns that might contain regulatory sequences or even encode RNAs with regulatory functions or a protein.
The SNP, rs3775291, is located in the exon region of the gene. The Leu412Phe (rs3775291) TLR3 mutation involves phenylalanine-to-leucine substitution, affecting a residue in the hydrophobic core of the TLR3 extracellular domain (ectodomain, ECD), which is characteristic of multiple leucine-rich repeats (LRRs). Consequently, the Leu412Phe mutation affects the response of TLR3 to its ligands, presumably by destabilizing the LRR architecture (Ranjith-Kumar et al., 2007).
Skevaki et al. reported that Leu412Phe might have a dominant protective effect on KSHV infection (Skevaki et al., 2015). The ability of minor allele variants at rs13126816 and rs3775291 to confer natural resistance to viral infection has been substantiated by other studies, showing that these protective alleles were less common in patients infected with human herpes simplex virus type 2 (HSV-2) (Svensson et al., 2012) or HIV type 1 (Sironi et al., 2012). However, the pathogenic mechanism by which TLR3 affects the risk of KSHV infection requires further investigation.
Linkage disequilibrium and haplotype analysis showed a strong linkage between the studied SNPs. Interestingly, we found that the CGAC combination, including rs3775291, might decrease the risk of KSHV infection in the Uyghur population. No risk effects were detected for other relevant SNPs within the block of KSHV infection in the analysis of individual sites. Thus, we inferred that the protective effect of this haplotype on KSHV infection might be due to certain combined effects between related sites.
In addition, KSHV-infected individuals with homozygous rs13126816 (AA) genotypes had a lower KSHV viral load, indicating that TLR3 polymorphisms affect the signals of latent KSHV reactivation. Previous studies have indicated that KSHV establishes a lifelong latent infection within host lymphocytes, with viral reactivation being required to maintain reservoirs of latently infected cells in chronically infected hosts (Cesarman, 2002; Grundhoff and Ganem, 2004). Reactivation of KSHV has been extensively characterized in established latently infected cell lines in vitro (West and Wood, 2003) and in the MHV68 model in vivo (Bussey et al., 2014).
Our results were consistent with the increased output viral titer following the stimulation of MHV68 latently infected B cell lines with TLR3 ligands (Gargano et al., 2009). Thus, TLR3 polymorphisms appear to be important in controlling the signal that reactivates KSHV from latency. However, this association disappeared when the population was divided according to ethnicity, potentially owing to the heterogeneity of KSHV infection across ethnic groups (Uyghur, aOR = 0.19, 95% CI: 0.01–0.97, p = 0.111).
To assess the possible functional effects of the two TLR3 SNPs identified in this study, we measured the level of IFN-γ in KSHV-infected individuals. However, we were not able to correlate these TLR3 SNPs with KSHV- or poly (I: C)-induced secretion of cytokine (i.e., IFN-γ), which was inconsistent with previous studies, where poly (I: C) in the presence of inflammatory cytokines induced the production of IFN-γ from NK cells (Tu et al., 2011).
This was probably because we did not consider the contribution of other factors besides KSHV infection in the levels of secreted IFN-γ. Therefore, further experiments that will consider all these factors are required.
Our study had some limitations. First, the study did not cover all TLR-related SNPs; we only focused on nine TLR3 SNPs that highly correlated with the viral infection. Second, due to its cross-sectional design, our study lacked temporal information and was thus insufficient for causal inference. To improve its strength and better understand the underlying mechanisms, the results of this study need to be verified in other KSHV-infected populations.
Conclusions
We found that genetic variations in the TLR3 gene might affect the risk of KSHV infection and its viral load. These small, individual genetic variations in TLR genes appear to compensate for the absence of host genetic factors in KSHV susceptibility and might affect the function of antiviral receptors and, consequently, susceptibility to KSHV infection and KSHV reactivation from latency.
Our study provides empirical data as a basis for further in-depth and extensive research.
Footnotes
Authors' Contributions
All authors contributed to the study conception and design. X.Z., X.W., and T.C. were responsible for data collection; Y.X. and Y.L. were responsible for sorting and cleaning the data; data analysis was performed by X.Z. and H.W.; X.Z. was responsible for interpreting the results and drafting the article; and T.Z. was responsible for revising it critically for important intellectual content and final approval.
Disclaimer
The funders did not play a role in the design, conduct, or analysis of the study, nor in drafting of this article.
Ethics Approval and Consent to Participate
The study was conducted with the understanding and the consent of each participant, and ethics approval was obtained.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by the National Natural Science Foundation of China (Grant No. 81772170) and by the Special Project on National Science and Technology Basic Resources Investigation (Grant No. 2019FY101103).
Supplementary Material
Supplementary Figure S1
Supplementary Figure S2
Supplementary Figure S3
Supplementary Table S1
Supplementary Table S2
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.