
Abstract
Coronavirus disease 2019 (COVID-19), caused by the novel severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2), rapidly resulted in a pandemic constituting a global health emergency. As an indicator of long-term immune protection from reinfection with the SARS-CoV-2 virus, the presence of memory B cells (MBCs) should be evaluated. Since the beginning of COVID-19 pandemic, several variants of concerns have been detected, including Alpha (B.1.1.7), Beta (B.1.351), Gamma (P.1/B.1.1.28.1), Delta (B.1.617.2), and Omicron (BA.1) variants with several different mutations, causing serious concern regarding the increased frequency of reinfection, and limiting the effectiveness of the vaccine response. At this regard, we investigated SARS-CoV-2-specific cellular immune responses in four different cohorts: COVID-19, COVID-19 infected and vaccinated, vaccinated, and negative subjects. We found that MBC response to SARS-CoV-2 at more than 11 months postinfection was higher in the peripheral blood of all COVID-19 infected and vaccinated subjects respect to all the other groups. Moreover, to better characterize the differences of SARS-CoV-2 variants immune responses, we genotyped SARS-CoV-2-positive samples from the patients' cohort. We found a higher level of immunoglobulin M+ (IgM+) and IgG+ spike MBCs in SARS-CoV-2-positive patients (5–8 months after symptoms onset) infected with the SARS-CoV-2-Delta variant compared with the SARS-CoV-2-Omicron variant implying a higher immune memory response. Our findings showed that MBCs persist more than 11 months after primary infection indicating a different involvement of the immune system according to the different SARS-CoV-2 variant that infected the host.
Introduction
Coronavirus disease 2019 (COVID-19), caused by the novel severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2), rapidly resulted in a pandemic constituting a global health emergency. The COVID-19 pathological process exhibits a wide spectrum of clinical manifestations, ranging from asymptomatic to critical disease. Moreover, the ongoing spreading of SARS-CoV-2 variants has increased alarms about the effectiveness of monoclonal antibody therapies and vaccines efficiencies. These variants have been classified as variants of interest or variants of concern (VOCs) by the World Health Organization.
The more recently identified Omicron (BA.1) variant, designated as a new VOC, has led to a surge in COVID-19 cases in South Africa and is now spreading across the world (Karim and Karim, 2021). Antibody responses rapidly increase about 2–3 weeks after infection followed to a slow decline phase until undetectable levels of antibodies are reached (Dan et al., 2021). In most individuals, anti-SARS-CoV-2 serum antibodies persist for more than 6 months after primary infection, but some patients rapidly lose their specific antibodies, especially those who experienced a mild disease course (Zheng et al., 2021).
It has been proposed that, in addition to serum antibody titers, the SARS-CoV-2-specific memory B cells (MBCs) should be evaluated to estimate humoral immunity as an indicator of immune protection (Abayasingam et al., 2021; Baumgarth, 2021; Hartley et al., 2020).
A recent study demonstrated that, unlike antibodies, levels of MBCs remain stable during the first 5 months after the infection (Vaisman-Mentesh et al., 2020). Other studies reported MBC responses up to 8 months after infection (Dan et al., 2021), and protection from reinfection is high in the first 6–9 months after infection (Hall et al., 2021; Vitale et al., 2021). If these findings can be true for a longer period, MBCs might confer a long-lived immunity against SARS-CoV-2. Therefore, it is of great importance to elucidate the factors underlying the formation of a robust MBC response against SARS-CoV-2. In Italy and other European countries, people with previous infection are now being offered a single vaccine dose 6 months after infection, on the assumption that past immunity will protect from reinfection (Margiotti et al., 2022).
Immune memory, from either primary infection or immunization, is the source of protective immunity from a subsequent infection (Orenstein and Ahmed, 2017; Piot et al., 2019; Plotkin et al., 2018) so analysis of the durability of immune responses following initial infection, correlation with protection from reinfection, and the capacity of these responses to neutralize VOC are important questions. In the current study, we characterize immune MBC responses in COVID-19-infected patients, in vaccinated patients, in vaccinated patients and COVID-19-infected patients (Hybrid immunity), and in negative subjects who never encountered the SARS-CoV-2 virus nor vaccination.
Materials and Methods
From July 2020 to April 2022, we enrolled 132 subjects at Altamedica diagnostic center of Rome. The study was conducted with the consent of all the participants and was approved by the internal ethics committee of Altamedica Laboratories, Artemisia S.p.A. Nasopharyngeal swabs were performed in all subjected enrolled in this study. All the symptoms in SARS-CoV-2-positive patients occurred between 5 and 7 days after the infection.
Among the 132 samples, we could distinguish four groups (Table 1): (i) 86 were COVID-19-infected patients (19 patients recruited 4 months postinfection, 32 patients recruited between 5 and 8 months postinfection, 21 patients recruited between 11 and 17 months postinfection, and 14 patients recruited 20 months postinfection); (ii) 9 were fully vaccinated patients (three doses of BNT162b2 mRNA vaccine), and all patients were recruited less than 6 months after last shot vaccination; (iii) 11 were COVID-19-infected patients and received at least one shot of vaccine (BNT162b2 mRNA vaccine), all patients were vaccinated after COVID-19 infection and were recruited less than 6 months after vaccination; and (iv) 26 samples had never encountered COVID-19 infection neither vaccination (named Negative).
Characteristics and Symptoms of the Enrolled Subject
IQR, interquartile range; ND, not detected; SARS-CoV-2, severe acute respiratory syndrome coronavirus 2.
Peripheral blood mononuclear cells were obtained from blood samples, and B cells were enriched using REAlease® CD19 MicroBead Kit according to the manufacturer's instructions (Miltenyi Biotec). To enable characterization of humoral and B cell responses to SARS-CoV-2, spike recombinant protein of the virus was used. For flow cytometric analysis, after magnetic isolation of SARS-CoV-2-specific B cells, antigen-tetramers were prepared by mixing recombinant SARS-CoV-2 spike-protein (HEK)-biotin with fluorescently labeled streptavidin (Phycoerythrin [PE] or PE-Vio770). For the magnetic separation, antigen-specific B cells that bound the antigen-tetramer-PE were subsequently labeled with anti-PE MicroBeads Ultrapure and magnetically separated over one MS column. For the identification and quantification of isotype spike-specific B after CD19+ selection, total MBCs were selected by CD27+, and isotype MBCs were instead stained with immunoglobulin G (IgG)-VioBlue-A, IgM-APC-A, IgA-VioGreen-A.
Afterward, double staining with two spike-tetramer solutions was used. Whenever further characterization of B cells was required, staining of cells using 10, flow cytometric data were analyzed with the MACSQuantify® Software. Furthermore, we genotyped SARS-CoV-2 mutations using TaqMan® Universal PCR Master Mix and specific custom TaqMan probe (Fabiani et al., 2022) and sequenced using next-generation sequencing (NGS) in some of SARS-CoV-2-positive patients. The sequencing analysis of SARS-CoV-2 was performed using AmpliSeq SARS-CoV-2 panel (Thermo Fisher Scientific). The reads from the library were aligned with the Wuhan-Hu-1 NCBI Reference Genome (Accession Number: MN908947.3) in Torrent Suite v. 5.10.1. A multivariate linear regression model using the enter approach was applied for simultaneously assessing the effects of independent variables on a quantitative dependent variable; p < 0.05 was considered statistically significant. Data quantification and statistical analysis were performed using GraphPad Prism.
Results and Discussion
The longevity of cellular and humoral immune responses after SARS-CoV-2 infection is a critical element informing the trajectory of the COVID-19 global pandemic. SARS-CoV-2 seroconversion occurs in >90% of individuals after infection (Gudbjartsson et al., 2020), with neutralizing antibodies and spike-specific IgG antibodies present up to 8 months after infection (Alfego et al., 2021). It has been shown that SARS-CoV-2-specific MBCs represent a potent strategy for additional immune protection (Dugan et al., 2021; Gaebler et al., 2021; Rodda et al., 2021). Current data indicate that antibody responses to SARS-CoV-2 infection decay more rapidly in comparison with other viral diseases (Huang et al., 2020).
In contrast, SARS-CoV-2-specific MBCs appear to be more persistent and may confer longer immunity (Hartley et al., 2020). We investigated SARS-CoV-2-specific immune MBCs in 132 samples. Among the 132 samples, we could distinguish four groups: (i) 86 were COVID-19-infected patients and not vaccinated (named Covid); (ii) 9 were patients fully vaccinated with three doses of BNT162b2 mRNA vaccine (named Vaccinated); (iii) 11 were COVID-19-infected patients who received at least one shot of vaccine after their infection (named Hybrid immunity); and (iv) 26 samples had never encountered COVID-19 infection or vaccination (named Negative) (Table 1). The presence and magnitude of SARS-CoV-2-specific MBCs were evaluated by flow cytometry (MACSQuant® Analyzer 10 Flow Cytometer; Miltenyi Biotec) identifying cellular subsets using a specific sequence of manual gating (Fig. 1).
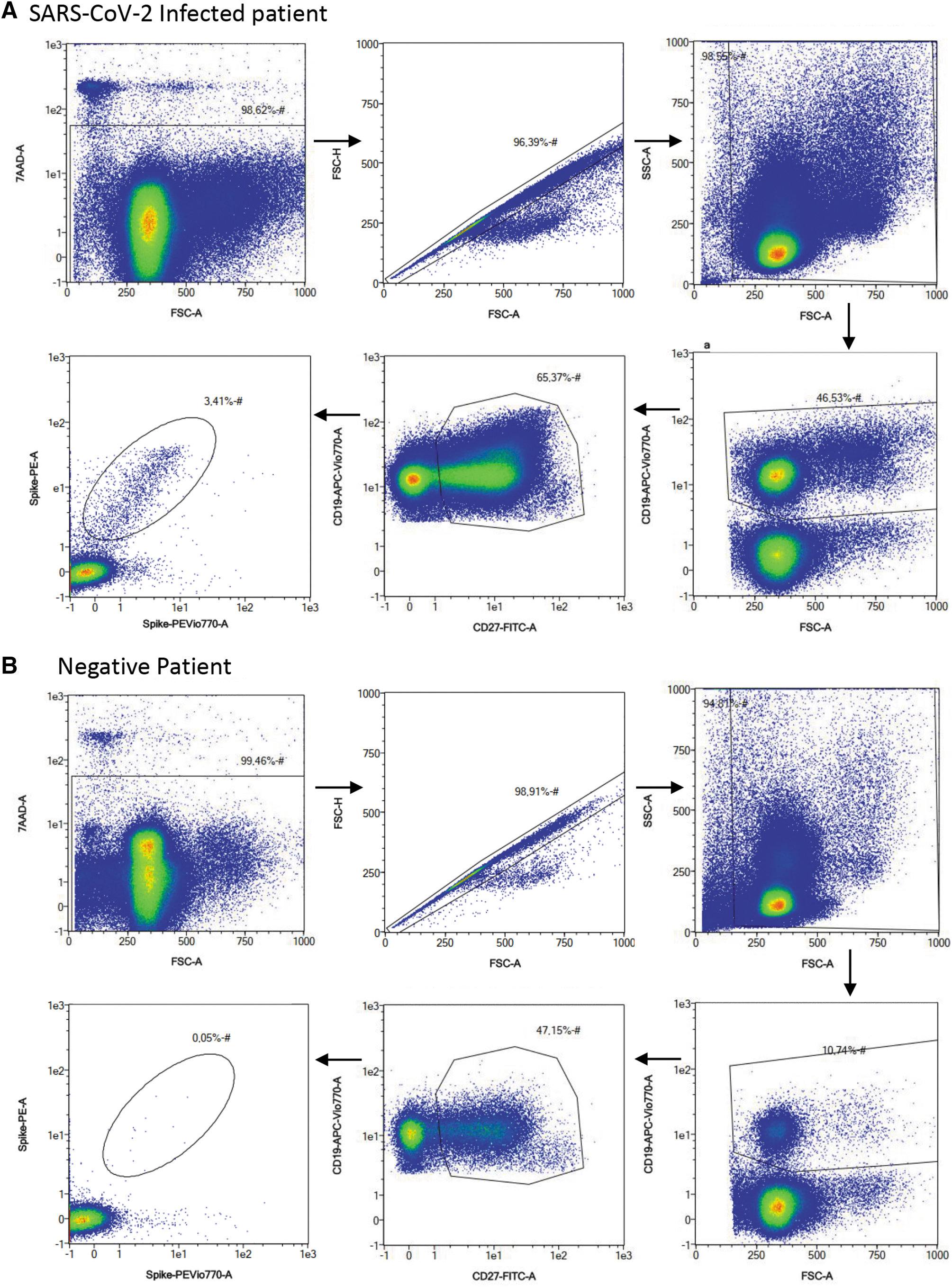
Gating strategy for the identification and quantification of spike-specific B cells. Peripheral blood mononuclear cells of COVID-19 convalescent
COVID-19-infected patients and not vaccinated (n = 86, Covid) were divided into groups based on time postinfection: less than 4 months postinfection (n = 19), between 5 and 8 months postinfection (n = 32), 11 and 17 months postinfection (n = 21), and more than 20 months postinfection (n = 14). We found that MBC frequency decreases with time, but interestingly, immune response results are still detectable in the group of SARS-CoV-2 patients 11–17 months postinfection (median = 0.18, min − max = 0.01–1.4) (Fig. 2A).
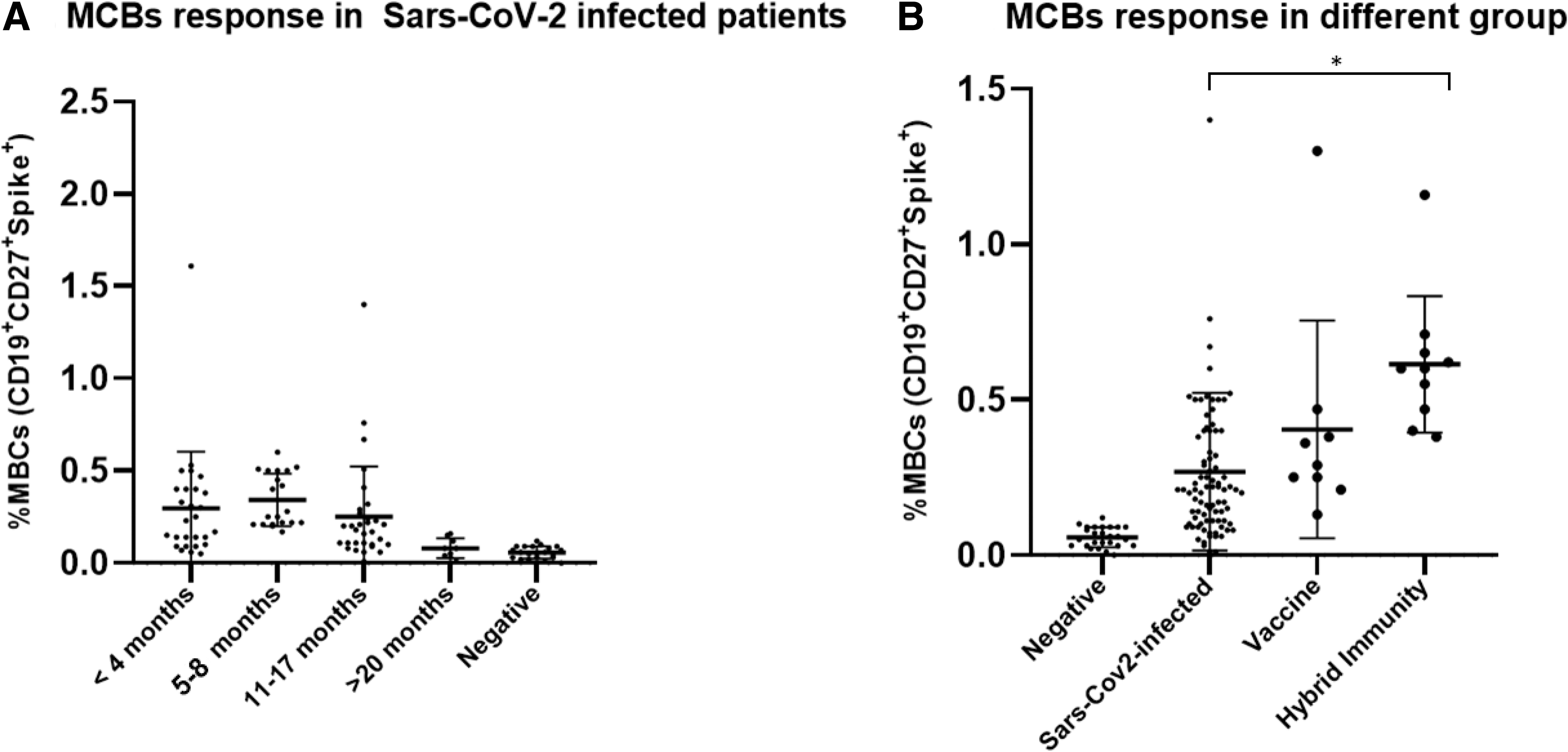
Graph showing frequencies of SARS-CoV-2-specific MBCs.
For patients who contract the virus more than 20 months, the MBC level was only slightly higher to negative subjects, with median value of 0.08 (min − max = 0.03–0.16) respect to the negative patients (median = 0.06, min − max = 0.01–0.12) (Fig. 2A). SARS-CoV-2-specific MBC magnitude response was different among the four groups (Fig. 2B). In particular, the Hybrid immunity patients had higher MBC response respect to the COVID-19-infected patients (p < 0.001), with median values of 0.53% and 0.21%, respectively (Fig. 2B). Whereas the levels of MBCs in SARS-CoV-2-infected patients were only slighter lower than vaccinated people never contracted the virus (Vaccine), with median values of 0.21% and 0.29%, respectively (Fig. 2B). In line with these data, Lederer et al. (2022) showed that vaccines elicit potent immune memory responses with an efficient induction of SARS-CoV-2-specific MBCs. According to these findings, individuals with hybrid immunity exhibit a stronger B cell response than those who exclusively get a viral or vaccine-induced response.
Multivariate linear regression analysis reveals that the measure of SARS-CoV-2 frequency of SARS-CoV-2-specific MBCs was not affected by age, gender, or symptomatology.
An understanding of the immune memory upon SARS-CoV-2 infection or vaccination is still ongoing, including the evaluation of MBCs in patients infected by different VOCs. In this study, to better characterized VOCs, we genotyped 32 SARS-CoV-2-positive nasopharyngeal swabs from patients in the 5 to 8 months postinfection group by quantitative polymerase chain reaction method and confirmed by NGS (Fabiani et al., 2022). Eleven of 32 analyzed samples result positive for Omicron variant and 21 for Delta variant. After genotyping, the correlations between VOCs and frequency of MBCs, together with IgA+, IgM+, and IgG+ spike-specific B cell isotypes, were investigated using the gating strategy presented in Figure 3.
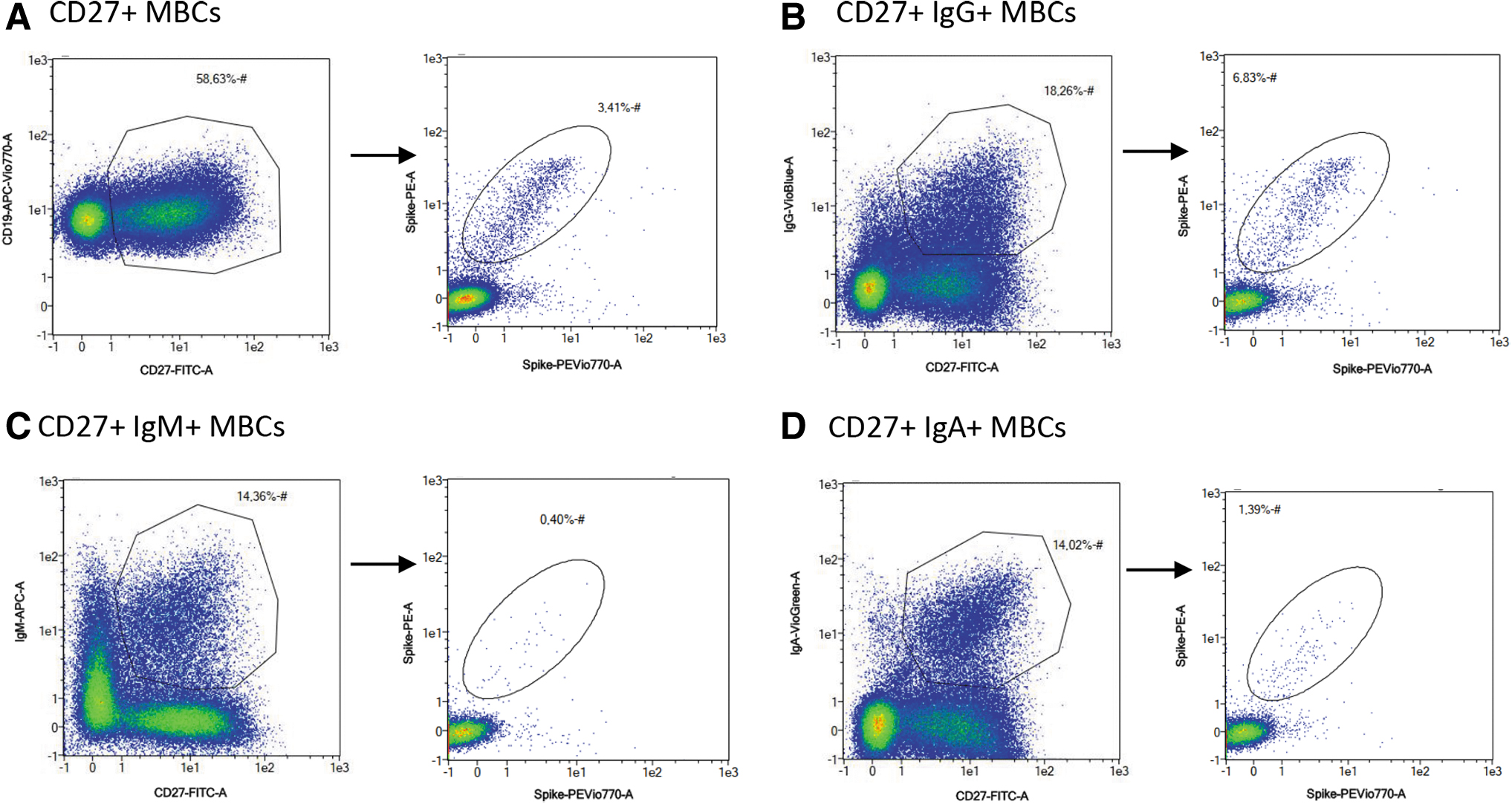
Gating strategy for the identification and quantification of isotype spike-specific B cells. Dead cells, doublets, and cell debris were excluded from the analysis using gating strategy presented in Figure 1. After CD19+ selection, total MBCs were selected by CD27+
Higher levels trend of MBCs was detected in patients affected by the Delta variant respect to patients affected by the Omicron variant, with median values of 0.32% and 0.20%, respectively (Fig. 4A). Differences in levels and duration of specific memory response among the variants may be associated with the capacity of the distinct variants of SARS-CoV-2 to escape of the early innate immune response mechanisms in the mucosal barrier, or the less affinity for some host receptors.

The comparison of SARS-CoV-2-specific MBCs between Omicron and Delta variants in SARS-CoV-2-affected patients.
Higher statistically significant results were obtained for both IgM+ and IgG+ in patients infected by Delta SARS-CoV-2 variant respect to patients infected by Omicron SARS-CoV-2 variant. In particular, the median value of the IgG+ spike-specific B cells for Omicron variant was 0.28% and for Delta variant was 0.75%, with p = 0.018, and the median value of the IgM+ spike-specific B cells for Omicron variant was 0.06% and for Delta variant was 0.12%, with p = 0.002 (Fig. 4C, D). No statistically significant difference was observed for the IgA+ spike-specific B cells, even though a positive trend correlation was detected (Fig. 4B).
Reactivation of antigen-specific MBCs can rapidly control pathogen replication as result of a massive antibodies production (Victora and Nussenzweig, 2012). In this study, we confirm that a predominant pool of SARS-CoV-2-specific IgG+ MBCs (median = 0.75% for Delta and median = 0.28% for Omicron variants) and a smaller population of IgM+ MBCs (median = 0.12% for Delta and median = 0.06% for Omicron variants) are produced following infection, as shown in the study by Lederer et al. (2022). In settings of seasonal re-exposure to a highly variable pathogen, the role of MBCs, IgM+ MBCs, and IgG+ MBCs humoral memory response is very important and must be better understood. Overall, based on the observation of this study, the MBCs, IgM+ MBCs, and IgG+ MBCs humoral memory response was present with higher frequencies in Delta variant than Omicron variant.
Potential limitations of this work include a limited number of subjects enrolled for MBC characterization especially in SARS-CoV-2-infected group after 11-month postinfection.
Unfortunately, in the study, it was not possible to carry out subsequent follow-up for the single patients, thus it was not a longitudinal study; moreover, the patients were recruited at different time points both postinfection and post-vaccination, making the results more inhomogeneous within the groups.
Another limitation of our study was that we were able to investigate only B cells response, while memory T cell responses were not evaluated, which so far appear to be less affected by SARS-CoV-2 variant differences.
Despite the fact that this study employed the same recombinant SARS-CoV-2 spike protein, as other study did to analyze MBC frequencies in patients infected with different VOCs (Goel et al., 2022; Kared et al., 2022; Winklmeier et al., 2022), another potential limitation could be that the recombinant spike protein, derived from the WuhanHu-1 variant, might exhibit different cross-reactivity between VOCs (Cao et al., 2022; Liu et al., 2022).
Another important aspect that must be taken into consideration is that we assess the MBC frequencies in the blood that do not exactly reflect frequencies in mucosal compartment, that is directly involved to block the viral entrance in reinfection cases. Anyway, further research is required to better assess the maximum length of protection from reinfection.
In conclusion, in this study, we observed that MBC frequencies are still detectable in the group of SARS-CoV-2 patients after 11-month postinfection. Additionally, individuals with hybrid immunity show a greater MBC response compared with COVID-19-infected patients demonstrating that vaccinations induce effective MBC induction for SARS-CoV-2 and powerful immunological memory responses. Finally, higher levels trend of MBCs in patients affected with the Delta variant respect to patients affected with the Omicron were detected; as well as higher IgG+ and IgM+ spike MBCs were detected for Delta variant respect to patients affected with the Omicron.
This study suggest that Delta breakthrough infection produces higher response on the overall immune compartment responses, while infection with Omicron elicits more modest effects, which may explain why those who had an infection with the Omicron variant had reduced protection against reinfection and more transmissibility.
Institutional Review Board Statement
The study was approved by the local ethical committee of Artemisia S.p.A.
Informed Consent Statement
Informed consent was obtained from all subjects involved in the study.
Footnotes
Authors' Contributions
Conceptualization: K.M. and M.F. Methodology: F.M., A.C., and A.V. Writing—review and editing: K.M., M.F., A.M., and C.G. All authors have read and agreed to the published version of the article.
Author Disclosure Statement
The authors declare no conflicts of interest.
Funding Information
This research received no external funding.