
Abstract
In this study, we reported the expression and potency of the recombinant H1N1 hemagglutinin (HA) vaccine as our in-house vaccine in a BALB/c mouse model. Recombinant H1N1 HA was produced in SF9 cell line, purified and formulated in MF59 adjuvant. Experimental mice were injected on days 0 and 14 with MF59-formulated vaccine, alum-based vaccine, and phosphate-buffered saline (PBS). Interleukin (IL)-2, IL-4, and interferon (IFN)-γ were assessed with commercial enzyme-linked immunosorbent assay (ELISA). Antibody responses and cytotoxic T lymphocyte (CTL) activity were assessed by hemagglutination inhibition and granzyme B ELISA, respectively. Moreover, the mice were challenged to show the vaccine efficacy. A considerable rise in IFN-γ and IL-4, as well as IFN-γ/IL-4 ratio, was observed in comparison with the alum-based vaccine and PBS group. Furthermore, our candidate vaccine showed superiority in humoral immune responses and CTL activity versus the alum-based vaccine and PBS group. The challenge showed that the survival rate in the vaccinated groups revealed a significant increase as compared with that in the PBS group. In conclusion, our candidate vaccine showed a robust Th1 response and CTL activity the alum-based vaccine. Moreover, a significant humoral immune response and a higher survival rate were detected in our vaccine as compared with the alum-based vaccine. It seems that the superiority of the MF59-based vaccine is due to the type of vaccine formulation in the candidate vaccine.
Introduction
The Orthomyxoviridae family includes influenza viruses and is the causal agent of influenza, a respiratory disorder in humans with a 500,000 mortality rate annually (Nypaver et al., 2021; Te Velthuis and Fodor, 2016). Symptoms of the flu virus disease vary from a mild infection in the upper respiratory to headache, cough, throat pain, fever, rhinorrhea, fatigue, muscular ache, and in some instances, lethal pneumonia (Krammer et al., 2018). The virus is classified into types A, B, C, and D, in which types A and B cause the majority of human infections (Tregoning et al., 2018). Across time, each pandemic was caused by a different type of virus, putting millions of people's health at risk all across the world (Cox, 2003; Kilbourne, 2006).
Types A and B include eight RNA fragments encoding RNA polymerase subgroup, viral glycoproteins such as hemagglutinin (HA) and neuraminidase, nuclear export protein, nucleoprotein, the nonstructural protein (NS1), matrix protein (M1), and membrane protein (M2), whereas types C and D have only seven RNA fragments and do not seem to be an important cause of severe diseases in people (Xie et al., 2011; Zheng et al., 2011).
The previous influenza vaccines, called Trivalent vaccines, contained two inactive types A (H1N1 and H3N2) and one subtype of influenza B, but nowadays a quadrivalent influenza vaccine includes H1N1, H3N2, and both of the influenza B subfamilies, with a higher coverage (Chen et al., 2020; Tisa et al., 2016). One of the commercially available vaccines is the recombinant HA vaccine, which is generated in insect cells. Owing to its importance in the early phases of viral infection, the HA protein is an appealing target for vaccine development (Hashem, 2015). The baculovirus is a strong expression system for recombinant proteins, yielding the same protein modifications as mammalian cells, including phosphorylation, folding, and polymerization (Rashedi et al., 2020; Tripathi and Shrivastava, 2019; Yee et al., 2018).
Adjuvants are one of the most important components in vaccine formulation, which increase the effectiveness and immunogenicity of vaccines (Rashedi et al., 2020). MF-59, as an oil-in-water adjuvant, was demonstrated to stimulate humoral and cellular immune responses in influenza vaccines (Ko and Kang, 2018), making the vaccines more effective (Galli et al., 2009; Ko and Kang, 2018; O'Hagan et al., 2011). To effectively deliver antigens to CD4 T helper cells, fast activation of antigen presenting cells at the injection loci and lymph node evacuation is essential, in turn providing immunologic support for the development of cytotoxic T cell and B lymphocyte cell adaptive immune system. This mechanism is a common feature of MF-59 adjuvant in the influenza vaccine formulation (O'Hagan et al., 2017; O'Hagan et al., 2012).
In this study, recombinant human H1N1 HA was produced in insect cells and the candidate vaccine was then formulated in the MF-59 adjuvant and injected to the experimental mice. Subsequently, cytokine responses, including interferon (IFN)-γ, interleukin (IL)-4, and IL-2 as well as cytotoxic T lymphocyte (CTL) activity, were assessed. Moreover, humoral immune response was assessed through hemagglutination inhibition (HI) and the efficacy of the vaccine candidate was determined by experimental viral challenge. All of these parameters were compared with an alum-based recombinant human H1N1 HA vaccine.
Materials and Methods
H1N1 HA expression vector construction, expression, and purification
In our previous study, the HA cDNA from the H1N1 strain was provided and placed into a pFastBac HT A vector. Afterward, the Escherichia coli TOP10 strain, as a host, was used for the gene transformation using the standard protocol. The pFastBac HT A vector was transferred to DH10Bac to make a recombinant Bacmid. The recombinant protein expression in the SF9 cell line was confirmed by sodium dodecyl sulfate–polyacrylamide gel electrophoresis (SDS-PAGE) and western blotting. T125 flasks were used to culture infected Sf9 cells for production of the recombinant HA in high quantity, and the recombinant molecule was purified by a Nickel-NTA column (Qiagen). The recombinant HA was dialyzed after purification to remove toxic material using phosphate-buffered saline (PBS) buffer and quantified by Bradford and stored at −70°C until use (Rashedi et al., 2020).
Experimental mice
Six- to 8-week female BALB/c mice (N = 45) were purchased from the Pasteur Institute of Iran. The experimental mice were maintained at the Razi Institute of Vaccine and Serum Research Institute's animal house facilities (Karaj, Iran), allowed ad libitum access to food and drink for a week, and maintained in a standard light/dark cycle (12 h/12 h) at 20–22°C. Animal handling was in accordance to the ethical committee's recommendations in the research of AJUMS (Ethics Code
Vaccine formulation
To formulate the recombinant HA protein in MF59 adjuvant (SEPPIC, France), 4 μg of the recombinant protein at the V/V ratio of 50:50 was added to the MF59 adjuvant and mixed for 30 min by vortexing (Vaccine Formulation Laboratory, Razi Vaccine and Serum Research Institute, Karaj, Iran). After this time, a homogen product was developed and used for immunization. Alum-HA was formulated by the standard protocol. In brief, 4 μg of the recombinant HA protein was mixed with 200 μg Alum (Pasteur Institute, Iran) and shacked 100 rpm for 60 min at room temperature. The reaction was conducted at pH 6–7 in double distilled water and, the product was adjusted with PBS after shacking and used for immunization purpose (this vaccine is gifted from Department of Influenza Vaccine of Razi Vaccine and Serum Research Institute of Iran). In the final product of both vaccines, each 100 μL of the vaccines contained 4 μg of the recombinant HA protein used for immunization.
Study planning and immunization
BALB/c mice were divided into three experimental groups, each containing 15 mice. The experimental mice were immunized two times subcutaneously on days 0 and 14 with rHA-MF-59, rHA-Alum vaccine, and PBS as a negative control group. Two weeks after the second injection, the blood samples were obtained and the serum samples were gathered and refrigerated at −70°C until the next experiments.
Enzyme-linked immunosorbent assay of IL-4, IFN-γ, and IL-2 cytokines
To assess the quantity of IL-2, IL-4, and IFN-γ, experimental mice were i.p injected with 500 ng of the HA protein to activate memory T cells and cytokine release. Sera from the experimental mice were collected 16–18 h post i.p injection to assess the cytokines. Commercial enzyme-linked immunosorbent assay (ELISA) kits (Mabtech, Sweden) were used to measure IL-2, IL-4, and IFN-γ in the sera, as per the standard methods. The quantity of the cytokines (pg/mL) was determined using standard curve for each one. To calculate the IFN-γ/IL-4 and IL-2/IL-4 ratios, the amount of IFN-γ and IL-2 produced by every mouse was split by the amount of IL-4 cytokine produced.
CTL assessment
CTL activity was evaluated using granzyme B (Gr-B) production. Sera from the experimental mice injected with 500 ng of the HA protein were collected 16–18 h after i.p injection to assess the serum Gr-B level. Gr-B assessment was performed using the commercial Gr-B ELISA kit (eBioscience) using the company manual. Gr-B concentrations were estimated using the standard curve for every individual mouse and presented as pg/mL.
HI assessment
The presence of nonspecific HA activity in hen serum was checked using 0.25% chicken red blood cells (RBCs) (Razi Serum and Vaccine Research Institute, Alborz, Iran). Hem-adsorption against chicken RBCs (4%) was used to remove HA for 1 h at 4°C. Before the HI experiment, the viral inoculum was back-titrated to ensure that the HA units were accurate. Except for the first well, an initial 25 μL PBS buffer was dispersed into the all wells of 96-well plates.
Thereafter, the sera exposed to receptor destroying enzyme (Accurate) were serially diluted from 1:10 to 1:1,280. Afterward, 25 μL of the diluted sera was added to each well and then 25 μL of the virus (4 HA unit) for 60 min at 25°C room temperature. Then, 50 μL of 0.5% chicken RBCs were added to the solution, and the reaction mixture was kept at 25°C for another 60 min. Then, visual inhibition of HA was examined in pits, with well-defined RBC “buttons” or the teardrop pattern appearing on plate tilting as the indicator. The reciprocal of the greatest serum dilution able to entirely inhibit HA was used to calculate HI titers.
Experimental influenza challenge
Fourteen days after the last vaccination, 10 mice from every 3 group were infected intranasally with 5 × 104 PFU of mouse adapted H1N1 influenza virus available (optimized dose in preliminary study, Razi Vaccine and Serum Research Institute, Karaj, Iran). The mortality of experimental mice was recorded daily up to 10th day of challenge.
Statistical evaluation
The data for every experiment were reported as the mean ± standard deviation. The statistical analysis was performed using Mann–Whitney U test (Graph Pad Prism V 6.01). A p-value <0.05 in every case was considered as significant difference. The Kaplan–Meier log-rank test was used to examine the survival rate of experimental mice.
Results
IFN-γ response
Results from IFN-γ cytokine response showed that mice immunized with HA-MF59 exhibited a significant increase in IFN-γ cytokine secretion as compared with the alum-based vaccine (1033.56 pg/mL vs. 839.74 pg/mL, p = 0.0289) and PBS group (1033.56 pg/mL vs. 586.06 pg/mL, p = 0.0012). Furthermore, mice immunized with the alum-formulated vaccine exhibited a significant increase in IFN-γ secretion as compared with the PBS group (839.74 pg/mL vs. 586.06 pg/mL, p = 0.0012; Fig. 1).
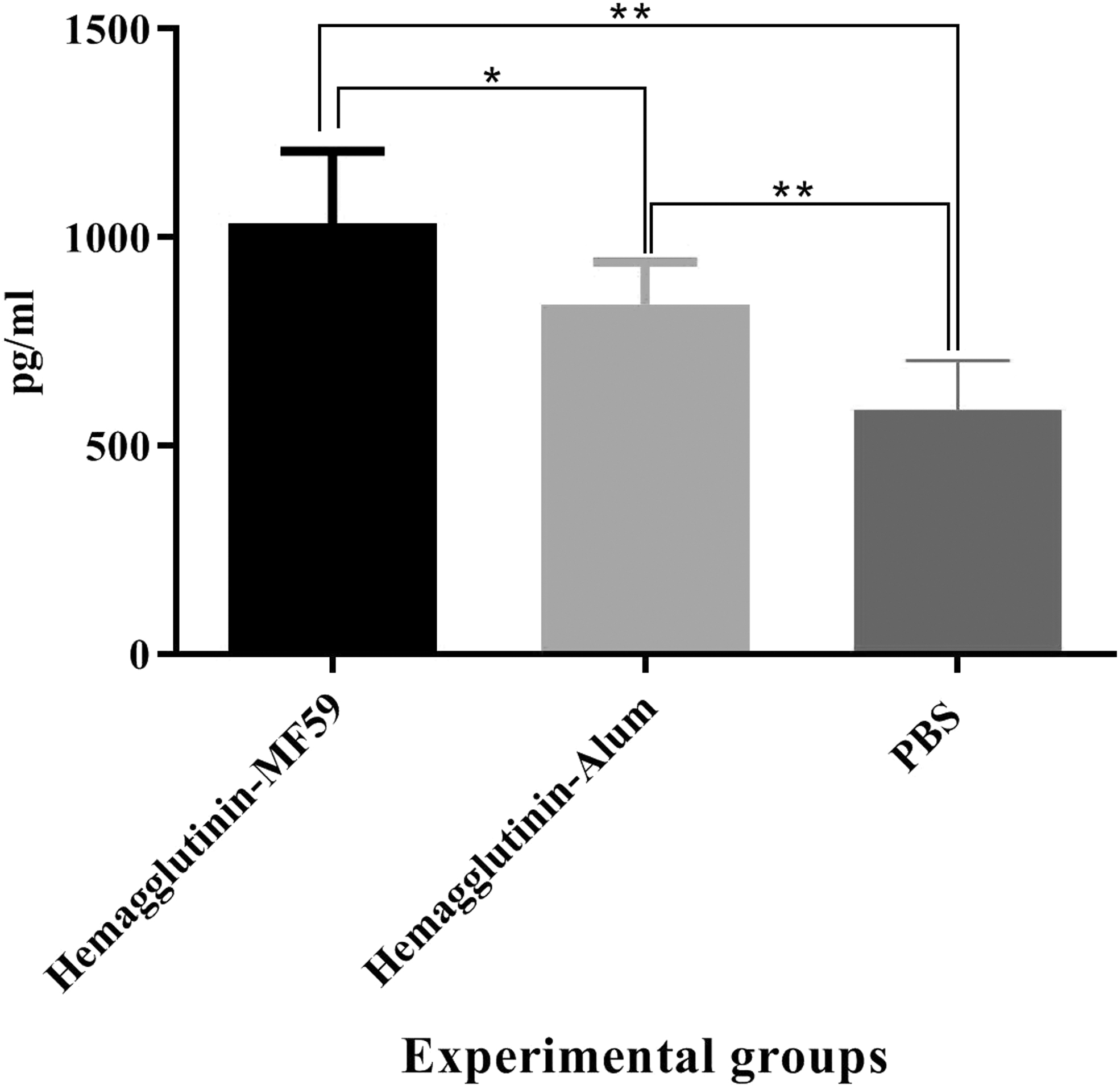
IFN-γ cytokine response in the vaccinated mice (N = 5/group). Immunization with recombinant hemagglutinin formulated in the MF59 adjuvant resulted in a significant increase as compared with the alum-based vaccine and PBS group (*p = 0.0289 and **p = 0.0012, respectively). IFN, interferon; PBS, phosphate-buffered saline.
IL-4 response
Results from the IL-4 cytokine in the MF59-adjuvanted vaccine showed a significant increase in IL-4 secretion as compared with the alum-based vaccine (471.29 pg/mL vs. 429.03 pg/mL, p = 0.0499) and PBS group (471.29 pg/mL vs. 419.26 pg/mL, p = 0.0104). However, IL-4 response in the HA-alum vaccine showed a small increase as compared with the PBS control group but statistically was not significant (429.03 pg/mL vs. 419.26 pg/mL, p = 0.8785; Fig. 2).
IL-4 cytokine response in the experimental groups (N = 5/group). Immunization with recombinant hemagglutinin formulated in the MF59 adjuvant showed a considerable increase as compared with the alum-based influenza vaccine and PBS group (*p < 0.0499). IL, interleukin.
IL-2 cytokine response
Results from IL-2 response showed that mice immunized with HA-MF59 displayed a significant increase in IL-2 secretion as compared with the PBS group (71.35 pg/mL vs. 66.01 pg/mL, p = 0.0205). In addition, mice immunized with the HA-alum vaccine did not show a significant difference as compared with the PBS control group (69.52 pg/mL vs. 66.01 pg/mL, p = 0.8665). Furthermore, mice immunized with HA-MF59 did not show a significant difference as compared with those immunized with the alum-formulated recombinant HA vaccine (71.35 pg/mL vs. 69.52 pg/mL, p = 0.3282; Fig. 3).

IL-2 cytokine response in the vaccinated groups (N = 5/group). Immunization with recombinant hemagglutinin formulated in the MF59 adjuvant showed a considerable increase as compared with the PBS group (*p = 0.0205). Nevertheless, the alum-based influenza vaccine failed to show a significant response versus recombinant hemagglutinin vaccine formulated in the MF59 adjuvant and PBS control group (p > 0.3282).
IFN-γ/IL-4 ratio
Results showed that mice immunized with the HA-MF59 and HA-alum vaccines increased the IFN-γ/IL-4 ratio as compared with the PBS control group (2.291 vs. 1.335, p = 0.0006 and 1.922 vs. 1.335, p = 0.0140, respectively). In addition, mice immunized with the HA-MF59 group increased the IFN-γ/IL-4 ratio as compared with the HA-alum vaccine but at the borderline range (2.291 vs. 1.922, p = 0.0513; Fig. 4).
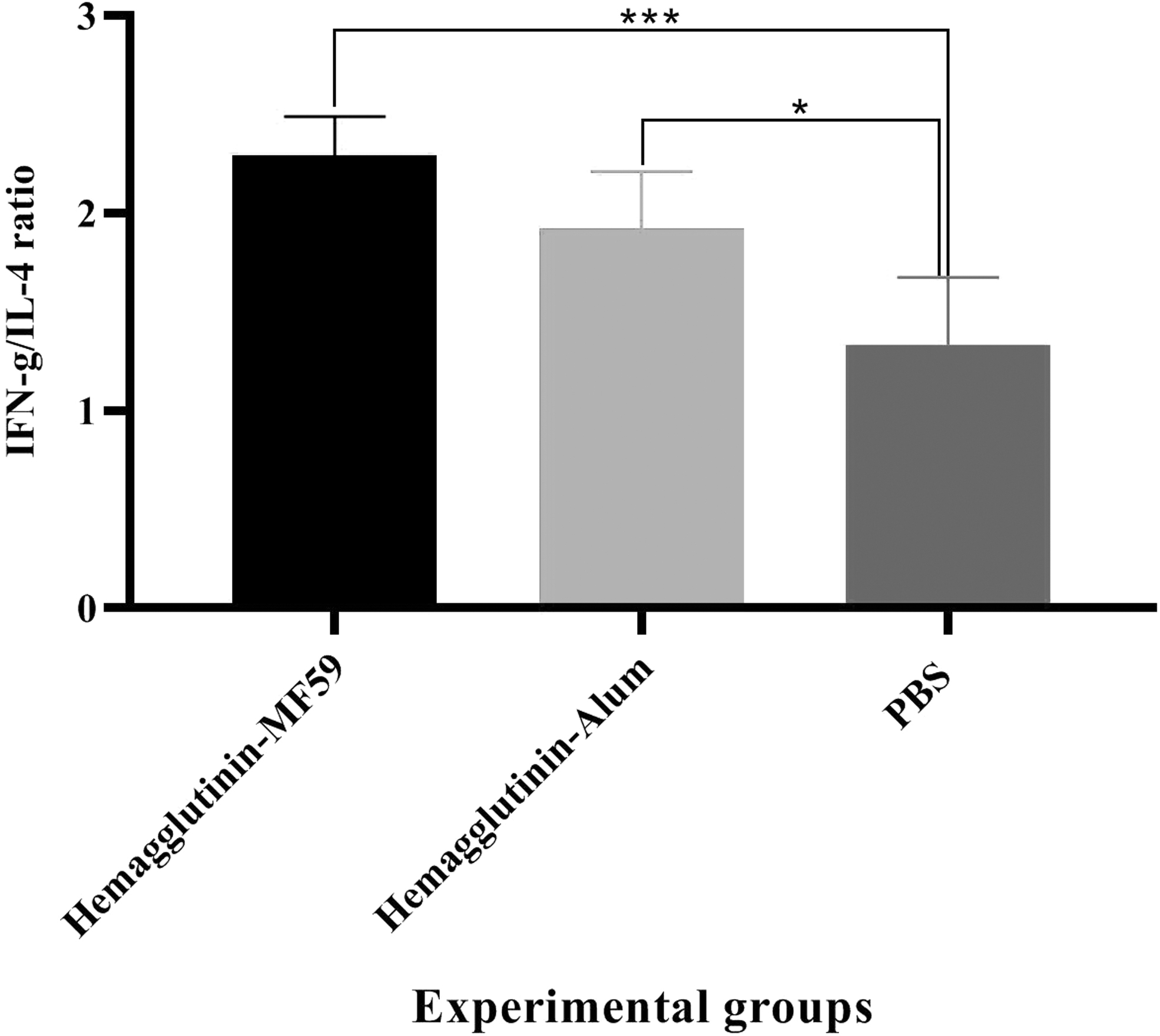
IFN-γ/IL-4 ratio in the mice groups N = 5/group). Recombinant hemagglutinin formulated in the MF59 adjuvant and alum-based influenza vaccine increased the IFN-γ/IL-4 ratio versus PBS control group (***p = 0.0006 and *p = 0.0140, respectively). Furthermore, the MF59-adjuvanted vaccine increased the IFN-γ/IL-4 ratio as compared with the commercial vaccine at the border line (p = 0.0513).
IL-2/IL-4 ratio
Results showed that mice immunized with the HA-MF59 and HA-alum vaccines did not show a significant difference as compared with the PBS group (0.153 vs. 0.149 and 0.162 vs. 0.149, p > 0.4286). Furthermore, the HA-MF59 group did not show a significant response as compared with the HA-alum vaccine (0.153 vs. 0.162, p = 0.4559; Fig. 5).
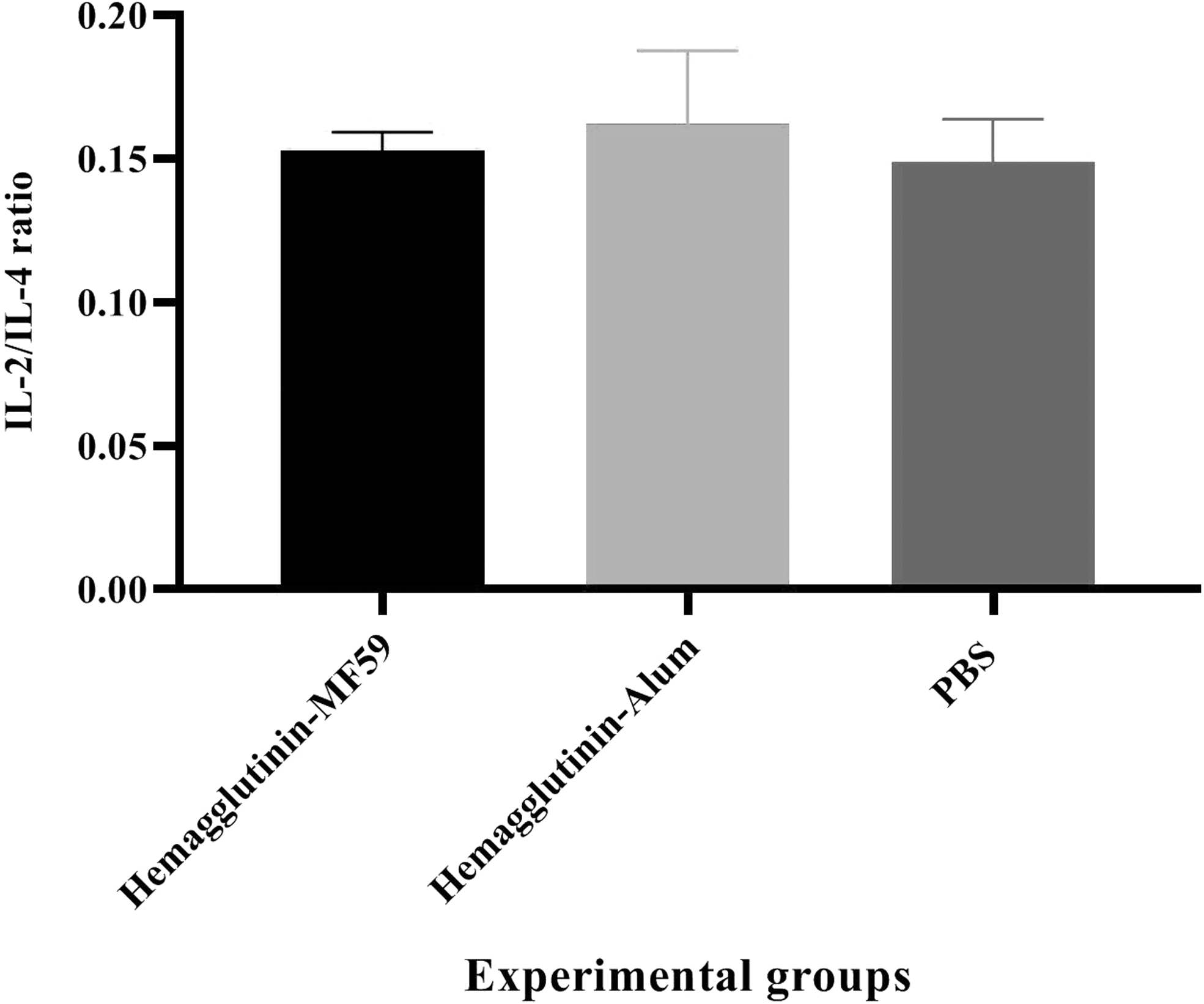
IL-2/IL-4 ratio in vaccine mice (N = 5/group). There were no significant differences among hemagglutinin formulated in the MF59 adjuvant, alum-based influenza vaccine, and PBS control group for the IL-2/IL-4 ratio (p > 0.4286).
Gr-B release
In this study, Gr-B release was assessed to show the CTL activity of each vaccine formulation. Mice immunized with the HA-MF59 and HA-alum vaccines showed a significant increase in Gr-B release as compared with the PBS group (659.09 pg/mL vs. 412.69 pg/mL and 537.21 pg/mL vs. 412.69 pg/mL, p < 0.0262). Moreover, mice immunized with HA-MF59 showed a significant increase in Gr-B release as compared with those immunized with HA-alum (659.09 pg/mL vs. 537.21 pg/mL, p = 0.0401; Fig. 6).
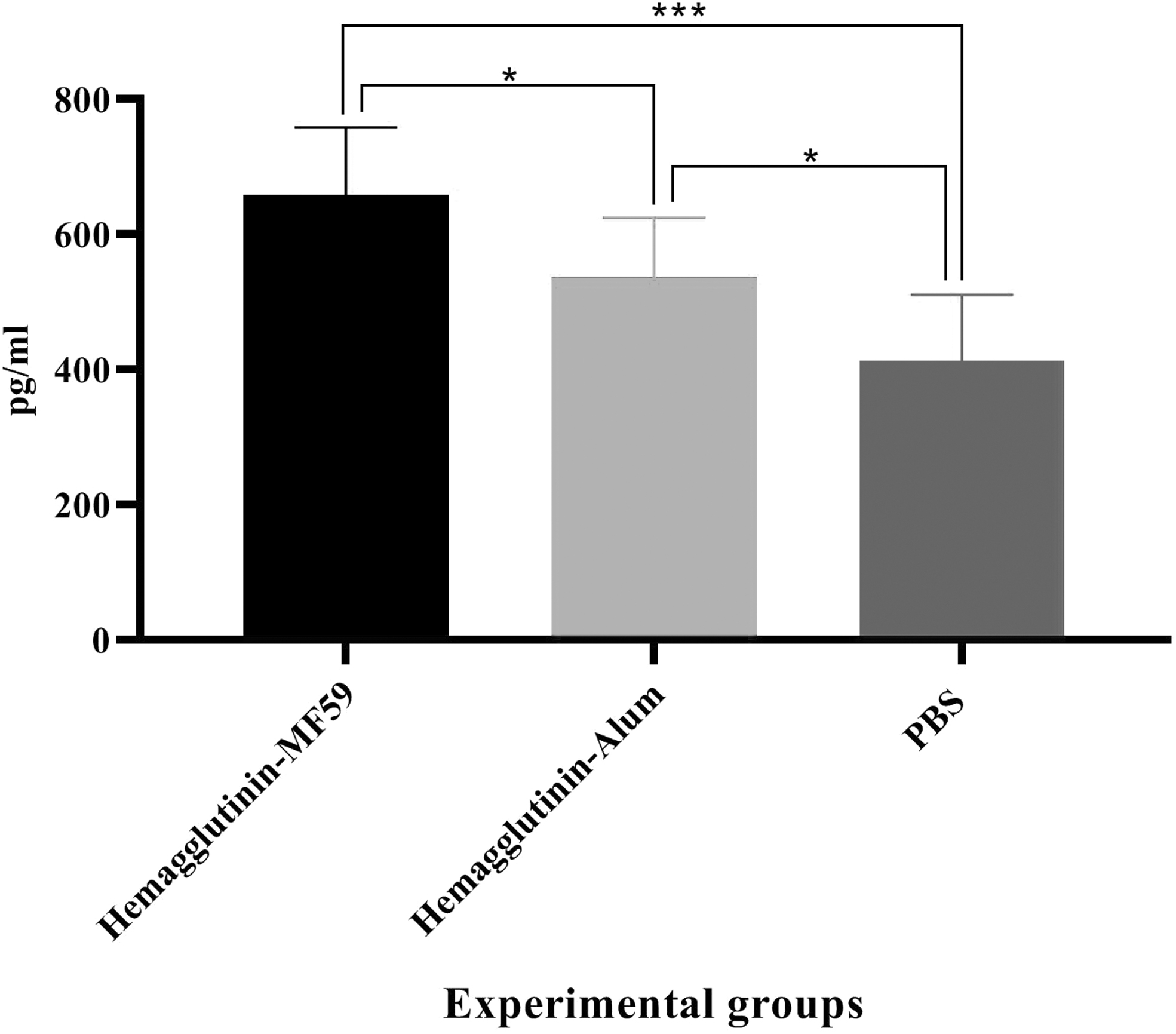
CTL activity in the experimental mice (N = 5/group). Immunization with recombinant hemagglutinin formulated in the MF59 adjuvant and alum-based influenza vaccine revealed a considerable increase as compared with the PBS group (***p = 0.0003 and *p = 0.0262, respectively). In addition, immunization with recombinant hemagglutinin formulated in the MF59 adjuvant revealed a considerable increase as compared with the alum-based influenza vaccine (*p = 0.0401). CTL, cytotoxic T lymphocyte.
Hemagglutination inhibition
Results from the HI assay, as a criterion of humoral immune response bioassay against the virus, showed that HA-MF59 and HA-alum vaccines showed a significant increase in the HI titer versus the PBS control group (2.67 vs. 0.07 and 2.09 vs. 0.07, p = 0.0022). In addition, the HA-MF59 group resulted in a significant increase in the HI titer as compared with the HA-alum vaccine (2.67 vs. 2.09, p = 0.0303; Fig. 7).
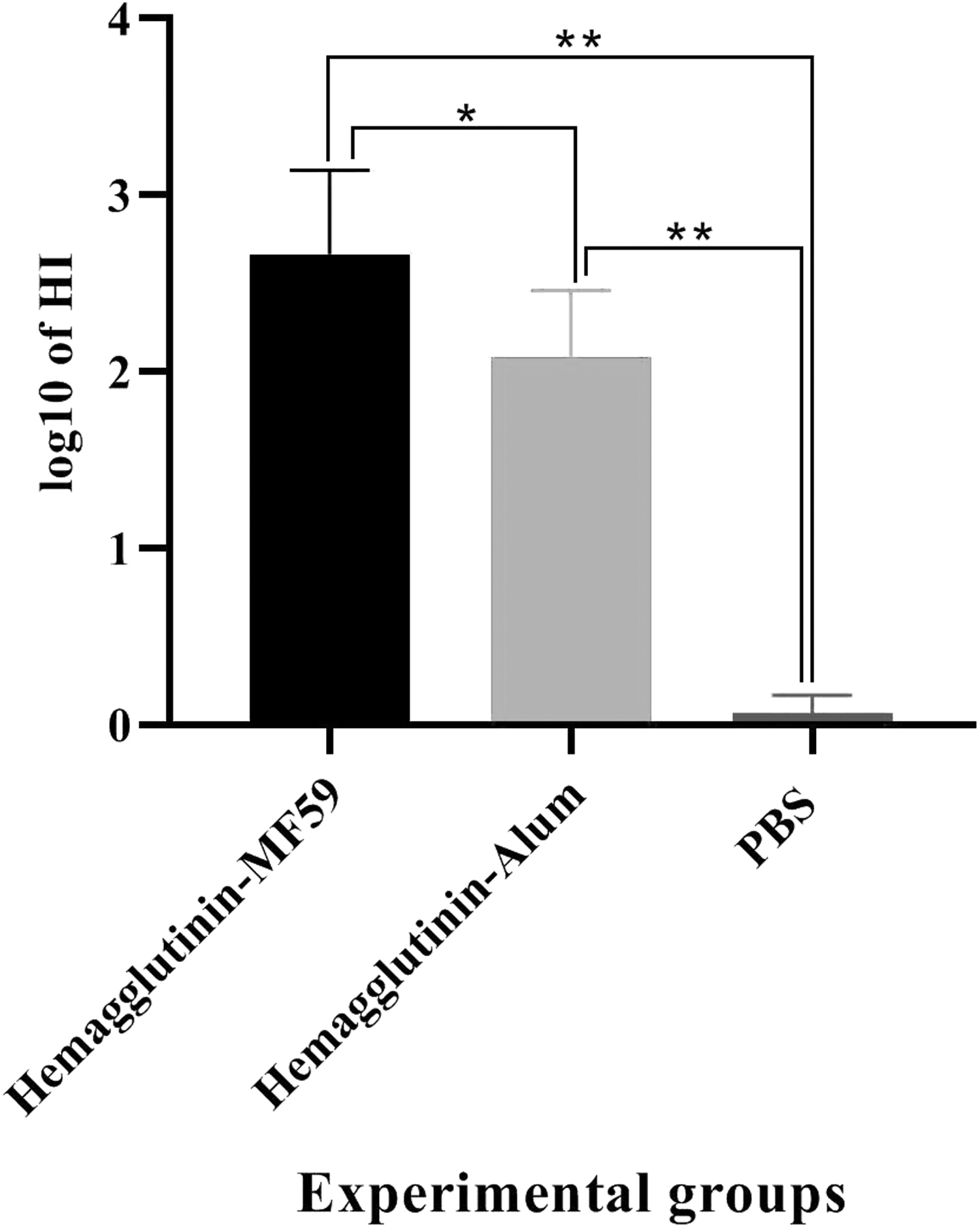
HI test on the chicken RBCs in the experimental groups using mouse adapted influenza H1N1 viruses (N = 5/group). HI response of the MF59-adjuvanted vaccine showed a considerable increase as compared with the alum-based influenza vaccine and PBS group (*p < 0.0303 and **p < 0.0022, respectively). HI, hemagglutination inhibition; RBC, red blood cell.
Survival rate
Result from the survival rate of vaccinated mice after monitoring for 10 days showed that mice immunized with the HA-MF59 and HA-alum vaccines indicated 90% and 80% survival rates, respectively. The mice in the PBS group, but not in the vaccinated group, showed a 10% survival rate (Fig. 8).
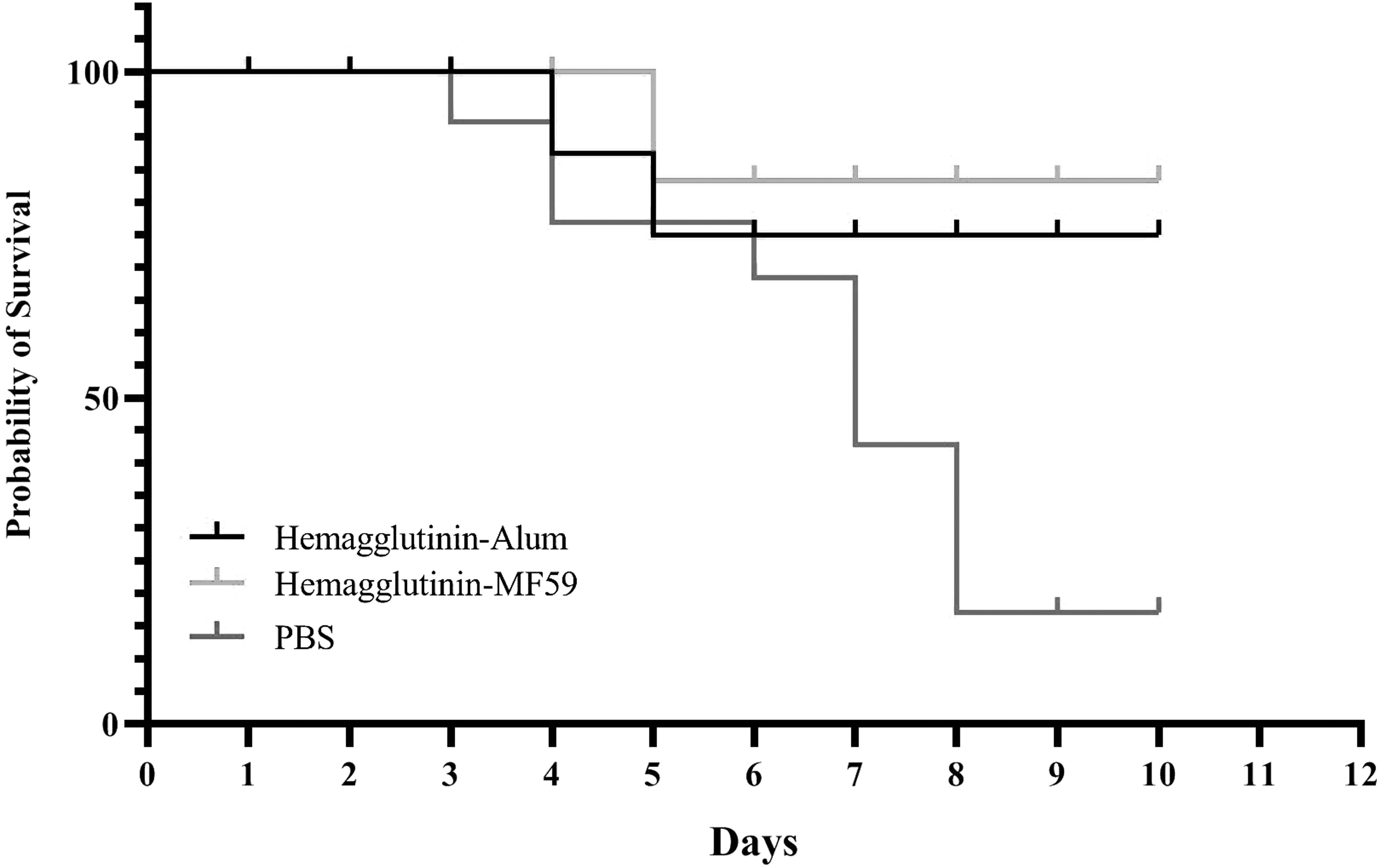
Experimental influenza challenge in the experimental mice (N = 10/group). The experimental groups were infected inside the noise with 5 × 104 PFU of mice H1N1 adapted influenza virus and the survival rate was monitored for the next 10 days.
Discussion
Vaccination against seasonal influenza resulted in controlling the incidence and mortality rate over the world. However, research in the field of the influenza vaccine is still a hot topic. As many countries have limitations in providing the vaccine for their countries, they are trying to produce in-house vaccines (Zahmati et al., 2021). In the previous study, recombinant human H1N1 HA was expressed in insect cells as an in-house vaccine candidate. Nevertheless, the immunologic mechanisms of this vaccine remained to be clarified (Rashedi et al., 2020). In this study, the cytokine pattern and the CTL activity of the vaccine were studied to clarify the immune polarization.
In addition, humoral immune responses and the vaccine efficacy were studied after a mouse adapted viral challenge. Our results showed that the MF59-based H1N1 influenza vaccine revealed more robust humoral immune response, IFN-γ and IL-4 cytokines, and even the IFN-γ/IL-4 ratio when compared with the alum-based H1N1 influenza vaccine. Both vaccines, although contained the HA antigen, were formulated in different adjuvants, one in the MF59 adjuvant whereas the other in the alum adjuvant. It seems that a stronger Th1/Th2 cytokines responses in MF59-based H1N1 influenza vaccine may be due to the nature of the adjuvant.
In a study, Ou et al. showed that a split H7N9 vaccine adjuvanted in MF59 was better than the alum-formulated vaccine in the triggering humoral immune response as well as IL-2 and IL-4 responses (Ou et al., 2016). In addition, an advantage in the induction of humoral immune responses versus alum-based vaccine was detected in primates using the MF59 adjuvant in the HBsAg vaccine formulation, confirming our findings in the induction of humoral immune responses (Traquina et al., 1996). Moreover, Wack et al. (2008) indicated that formulation of the influenza virus in MF59 emulsion and aluminum hydroxide adjuvant has a higher potency in the MF59-adjuvanted vaccine for antibody and IFN-γ responses.
These findings are in accordance with our findings that revealed more potency of HA-MF59 in triggering humoral immune responses and induction of the Th1 pattern. It is also proved that the Th1 cytokine response through induction of the CTL response is able to clear infected cells and viral clearance (Jansen et al., 2019; Tsoras et al., 2020). Therefore, the CTL response is next assessed through Gr-B release as a standard method for the detection of the CTL activity (Mahdavi et al., 2017; Mahdavi et al., 2011). The result showed that MF59-adjuvanted vaccine revealed a significant increase versus alum-based vaccine. This finding is in accordance with IFN-γ response, showing that the MF59 adjuvant is more effective than the alum adjuvant in the stimulation of cellular immune responses.
The CTL response is critical for killing viral infected cells and protection against influenza infection (Jansen et al., 2019; O'Hagan et al., 2011). Humoral immune responses by production of specific antibodies capable of binding to the surface proteins of virus involve in the neutralizing the virus and thereby can prevent the spread of infection. Therefore, experimental viral challenge was next performed on the vaccinated mice. The results of viral challenge showed that the recombinant HA vaccine in the MF59 adjuvant showed a 90% survival rate, whereas the alum-based vaccine and PBS group showed 80% and 10% survival rate, respectively.
Overall, results from experimental challenge showed that vaccination increases the survival rate of mice versus the control group. This may show that antibodies formed in the mice neutralized the virus and prevented the spreading of the virus. In addition, cellular immunity by stimulating TH1 and CTL responses might involve in the killing of infected cells and clearance of the virus by elimination of infected cells, which finally lead to an increase in the survival rate of vaccinated mice. Furthermore, MF59-adjuvanted vaccine showed a 10% increase in the survival rate in comparison with the alum-adjuvanted vaccine. This finding may be due to higher cellular and humoral immune responses in the MF59-adjuvanted vaccine, which resulted in a higher survival rate.
However, we believe that this advantage is due to the nature of the MF59 adjuvant used in the vaccine formulation. Studies showed that MF59-based influenza vaccine is highly potent in the fast priming CD4+ T cell responses, induction of long-lasting T and B cell responses as well as triggering cross-protection through broadening the immune response beyond the influenza strains used in the vaccine formulation. Such evidence proved the high performance of the MF59-based influenza vaccine, whereas the alum-based vaccine is not as efficient as MF59-based influenza vaccine (O'Hagan et al., 2011; Puig Barberà and González Vidal, 2007; Vesikari et al., 2009).
Finally, we showed that our in-house vaccine was highly effective and able to induce an immune response better than the alum-based vaccine. However, long-term immune responses and other immunologic mechanisms of this vaccine remained to be clarified; in the future, these aspects of vaccine potency need to be determined in animal models. In addition, cross-protection of influenza vaccine, as a critical potency about this vaccine, is another parameter requiring to be assessed in our future perspective about this vaccine.
Conclusion
In conclusion, our findings revealed that recombinant HA of the H1N1 influenza virus formulated in the MF59 adjuvant is more powerful than the alum-based vaccine in the stimulation of cellular and humoral immune responses.
Footnotes
Acknowledgments
We thank Dr. Ali Khodadadi from Ahvaz Jundishapur University of Medical Sciences and Mr. Emad Heydarnia for article editing. We also thank Dr. Akbar Khorasani for vaccine formulation from Department of FMD of Razi Vaccine and Serum Research Institute of Iran (Karaj, Iran) and Dr. Louiza Chitour, Regional Sales and Marketing Manager from Business Unit of SEPPIC Company, for providing MF59 adjuvant.
Authors' Contributions
Dr. Sepasi and Dr. Ghafourian are from Ahvaz Jundishapur University of Medical Sciences (Ahvaz, Iran); Dr. Taghizadeh is from Razi Vaccine and Serum Research Institute (Karej, Iran), and Dr. Mahdavi is from Motamed Cancer Center, Academic Center for Education, Culture and Research (Tehran, Iran) and Tehran University of Medical Sciences (Tehran, Iran), all authors were involved in study planing and performing.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This study has received grant from Deputy of Ahvaz Jundishapur University of Medical Sciences (Grant No. CMRC-0028).