
Abstract
Respiratory syncytial virus (RSV) causes annual epidemics of infections affecting the whole population. In vitro, it has been shown to infect and persist in human dendritic cells (DCs) for prolonged periods. Initially persistence is associated with low levels of replication before the virus becomes dormant. Reactivation of viral replication can be triggered many months later. Infection of DCs is likely to influence the host's ability to generate effective long-term memory responses. A well-established animal was utilized to confirm that RSV both infects and persists in pulmonary DCs in vivo. Mice were infected with a modified strain of RSV expressing red fluorescent protein (RSV-RFP) when replicating. Clinical symptoms of infection were monitored using weight change and inflammatory cell counts from bronchoalveolar lavage, which correlated with the RSV viral titer (quantitative polymerase chain reaction). Lung tissues were collected at 3, 5, 7, and 21 days postinfection (dpi) to assess leukocyte populations by flow cytometry. Clinical symptoms and RSV viral load peaked at 5 dpi. RSV-RFP was most prevalent in macrophages at 3 dpi and also observed in B cells and DCs. At 21 dpi, RSV-RFP remained evident in a subset of conventional DCs (CD103+CD11b+) even though both clinical symptoms and pulmonary inflammation had resolved. These results confirm that in this well-established mouse model, RSV persists in lung conventional DCs following resolution of the acute infection. Further work is required to explore whether the virus continues with low-level replication before becoming dormant in vivo, as has been described in vitro.
Background
The respiratory syncytial virus (RSV) causes annual epidemics of respiratory disease affecting the whole population, although its greatest impact is on the very young (Everard, 2008; Nair et al., 2010) and is estimated to be responsible for ∼200,000 deaths per annum among infants worldwide (Nair et al., 2010). The virus also has a significant impact on the elderly (Falsey, 2007) and those with chronic obstructive pulmonary disease (Wilkinson et al., 2006).
There is still no effective vaccine or treatment despite RSV being listed as a research priority by the World Health Organization (WHO) and more than five decades of research (Modjarrad et al., 2016). This lack of progress is surprising given that there are only two major RSV subtypes that do not demonstrate significant antigenic drift (Mejias et al., 2020).
Despite the lack of significant antigenic shift, individuals are reinfected throughout life, suggesting that long-term immunity is poor. This suggestion is supported by studies indicating that neutralizing antibody levels wane rapidly after an infection (Falsey et al., 2006; Sande et al., 2013).
These observations suggest that RSV is able to prevent the induction of effective long-term immune responses, which in turn results in poor herd immunity (Everard et al., 2009; Everard, 1999), with very young infants being particularly at risk due to low levels of passively acquired neutralizing antibodies. Elucidating the mechanisms that undermine the development of effective long-term memory responses to the virus is likely to be crucial for developing successful strategies to prevent RSV-induced illness.
While RSV is known to target airway epithelial cells for replication (Fonceca et al., 2012), it has also been shown to infect monocytes (Bartz et al., 2002; Domurat et al., 1985), including cultured myeloid-derived and differentiated human dendritic cells (DCs) (Hobson and Everard, 2008; Jones et al., 2006; Schwarze, 2008).
DCs are key antigen-presenting cells (APCs) within the lungs and are critical for induction of memory immune responses (Merad et al., 2013; Neyt and Lambrecht, 2013). It has been shown that RSV infection in both humans (Gill et al., 2008; Kerrin et al., 2017) and mouse models (Beyer et al., 2004; Lukens et al., 2009) is associated with a significant increase in DC numbers in the upper and lower airways.
It has also been shown in long-term in vitro studies that following infection of human monocyte-derived DCs, the virus continues to replicate at low levels for some months before becoming latent. Replication can be reactivated several months later following exposure to nitric oxide (Hobson and Everard, 2008) and this reactivation can also occur spontaneously the following winter (personal communication, Everard).
The observation that RSV can infect and persist in DCs may be key to understanding the virus's apparent ability to impair the development of effective long-term immunity. The ability to persist in pulmonary DCs may also provide the virus with a natural human reservoir during the interval between the characteristic winter epidemics during which the virus is rarely isolated (Everard, 2008; Nair et al., 2010).
A well-established murine model of RSV infection was utilized to explore the dynamics of virus–host cell interactions through and beyond an acute infection. To clearly demonstrate microlocalization of RSV in the lung and airways over the course and resolution of an acute infection, this study utilized a genetically modified strain of RSV designed to fluoresce when actively replicating (Grieves et al., 2010; Hobson and Everard, 2008).
At various time points during and after the resolution of an acute infection, bronchoalveolar lavage (BAL) samples were obtained to study the progressive rise and fall in RSV titers in the airways from infection to resolution and to study the associate airway inflammatory response at the same time points. At the same time points, lung tissue digests were obtained to study changes within lung tissue. Multiparameter flow cytometry was used to track fluorescing RSV and identify colocalization of the virus with particular monocyte cell populations, including DCs.
Materials and Methods
Virus culture
Mice were infected with a genetically modified strain of RSV derived from the RSV wild-type clone, MP224, designed to express red fluorescent protein (RFP) when replicating (Grieves et al., 2010; Hallak et al., 2000; Hobson and Everard, 2008) (gift from Professors Mark Peeples and Peter Collins, Nationwide Children's Hospital, Columbus, OH, USA, and National Institute of Allergy and Infectious Diseases, Bethesda, MD, USA, respectively).
Briefly, the virus was added to HeLa monolayers grown in flasks until the cytopathic effect reached ∼50% (48 h). The remaining medium slurry and HeLa monolayer were harvested and centrifuged 500 g. The supernatant was filtered by centrifugation to concentrate viral particles and remove remaining cellular debris. Plaque assay analysis was used to determine the titer of the RSV inoculum, as described previously, whereby RSV plaque formation was indicated by fluorescence (Grieves et al., 2010; Hobson and Everard, 2008).
Mice and RSV infections
Female BALB/c mice at 7–9 weeks of age (Animal Resource Centre, Murdoch, Western Australia) were housed in a controlled environment with a 12-h light–12-h dark cycle and unrestricted access to food and water. Mice were anesthetized with 3% isoflurane and infected intranasally with 1 x 105 pfu/mL live RSV in Dulbecco's modified Eagle's medium, with the total inoculation volume being no >40 μL. Mice were euthanized with pentobarbital (250 mg/kg) following inhaled methoxyflurane anesthesia.
Mice were weighed daily during the acute period of infection until sacrifice at 0, 3, 5, 7, and 21 days postinfection (dpi). Clinical disease scores were also assessed according to the following criteria, as described previously (Scott et al., 2016):
Score 0—normal appearance, healthy, and active.
Score 1—barely ruffled fur, mildly/intermittent hunched appearance, and otherwise healthy.
Score 2—moderately ruffled fur, elevated respiratory rate, hunched appearance with a crab-like gait, intermittent stillness, and reduction of curious behavior.
Score 3—ruffled fur, labored breathing, hunched appearance with a crab-like gait, and unresponsive to stimuli.
BAL analysis
Euthanized mice were tracheotomized for BAL, which was used to examine inflammatory cell infiltrate into the lungs following infection. Briefly, BAL fluid was collected by slowly infusing and withdrawing a 1-mL aliquot of 0.9% saline from the lung three times. Cell viability was determined by trypan blue staining, and differential cell counts were assessed with Leishman stain (Sigma-Aldrich, St. Louis, MO, USA).
Bronchoalveolar cell analysis for RSV titer
A portion of bronchoalveolar cells were snap-frozen in liquid nitrogen for analysis of the viral load by real-time quantitative polymerase chain reaction (qPCR). RNA lysis buffer was added to the frozen pellets and homogenized by pipetting on ice. Samples were centrifuged at 16,000 g for 5 min and total RNA extracted from resulting supernatants using the PureLink RNA extraction kit (Ambion, New Haven, CT, USA). A minimum of 2 mg was used as a template for cDNA synthesis using random primers and M-MLV reverse transcriptase (Promega, Southampton, United Kingdom).
The presence of RSV was measured using a commercially available TaqMan real-time PCR kit designed to determine the RSV titer by interpolation from a standard curve (RSVSpp, GeneSig, Southampton, United Kingdom). The PCR was analyzed using an Applied Biosystems ABI 5400 thermocycler.
Flow cytometry analysis
Lung tissue was harvested at 0, 3, 5, 7, and 21 dpi. These were prepared as single-cell preparations suitable for flow cytometry analysis by collagenase and DNase digest, as described [26]. Selected cell subpopulations were identified using an optimized panel of fluorescently conjugated monoclonal antibodies (BioLegend, BD Pharmingen) directed against cell surface antigen markers: CD11b-v500 (clone M1/70), CD11c-AF700 (clone HL3), CD103-PE (clone M290), F4/80-FITC (clone BM8; BioLegend, San Diego, CA, USA), Ly6G/C-APC-Cy7 (clone RB6–8C5), IA/I-E-AF647 (clone M5/114.14.2), and B220/CD45R-PE-C F594 (clone RA3–6B2), as described in Supplementary Figure S1.
A pooled cell preparation for each biological replicate was used to assess background fluorescence (nonstained control) and nonspecific staining (fluorescence minus one control). Data from all stained and nonstained tissues were acquired on a BD FACS Canto (Becton Dickinson, NJ, USA), and the RFP signal from fluorescently tagged RSV was also collected. Cell subpopulation analysis and RSV colocalization were completed used FlowJo software (Tree Star, Ashland, OR, USA), with gating strategies presented in Supplementary Figure S1.
Statistical analyses
A minimum of six biological replicates collected from experiments were completed in duplicate for noninfected controls, and each time point post-RSV infection was used for analysis. Statistical significance was assessed by the Kruskal–Wallis test, with Dunn's multiple comparisons, compared with control samples (0 dpi) with p < 0.05. The GraphPad statistical package and graphing tools were used to complete all analyses.
Results
Clinical course of RSV infection in adult BALB/c mice
Mice infected with RSV-RFP experienced greatest weight loss at 5 dpi (2.3 ± 0.7%; p < 0.05), returning to baseline at 7 dpi, followed by a significantly increased weight gain at 21 dpi compared with day 0 baseline weights (5.4 ± 1.5%; p < 0.01; Fig. 1A). Clinical symptoms (clinical score) were evident at 3 dpi, peaking at 7 dpi and returning to baseline at 21 dpi (Fig. 1B).
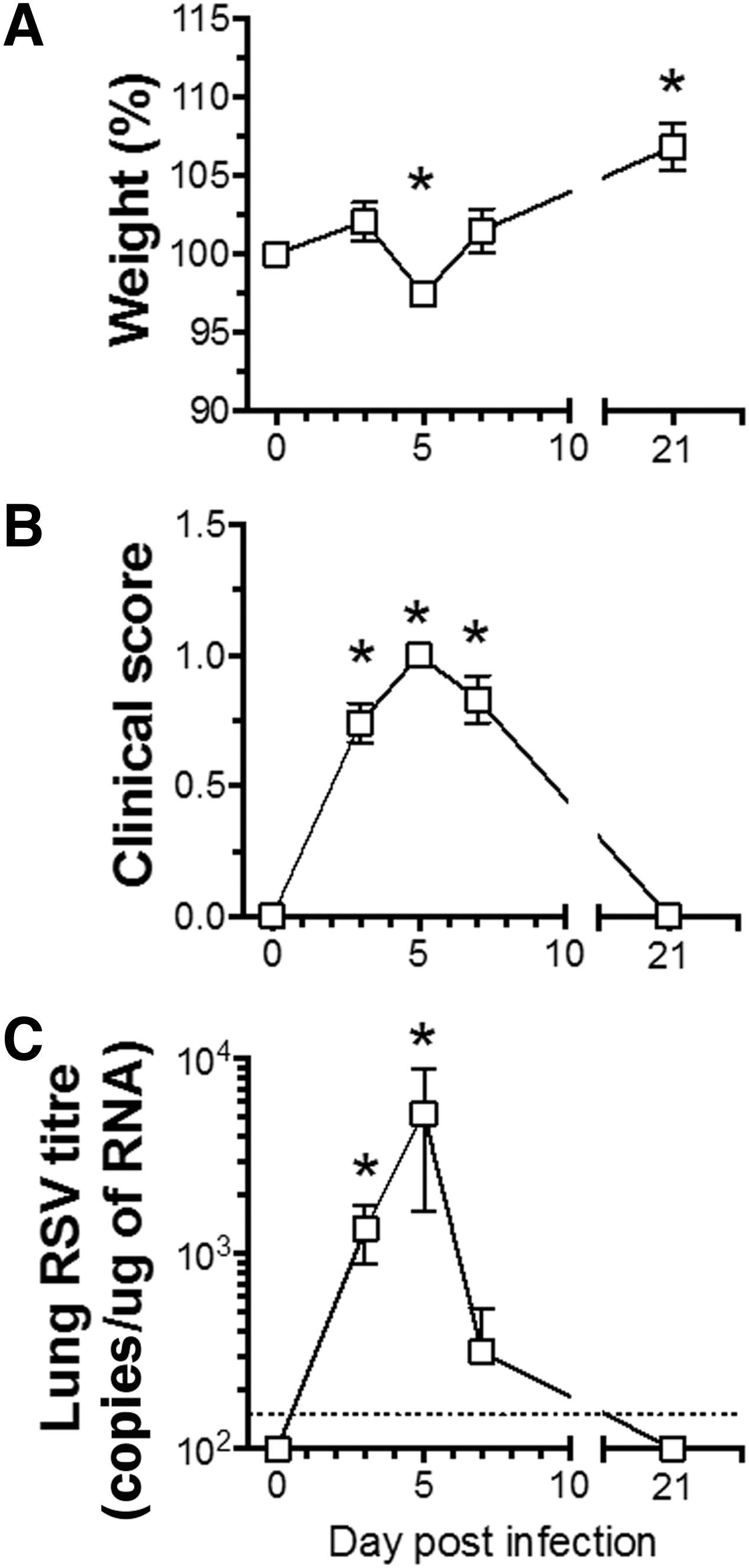
Clinical course of RSV infection in adult BALB/c mice. BALB/c mice were infected with RSV-RFP and followed up to 21 dpi.
Viral titers, as determined in bronchoalveolar cells using qPCR, were significantly elevated at 3 dpi, peaking at 5 dpi, declining by 7 dpi, and returning to undetectable levels by 21 dpi (Fig. 1C).
Inflammatory cell responses in BAL fluid following RSV-RFP infection
Total BAL cell numbers were significantly elevated at 5 dpi, returning to baseline by 7 dpi (Fig. 2A), while macrophage numbers were significantly reduced at 3, 5, and 7 dpi, returning to baseline by 21 dpi (Fig. 2B). Total numbers of neutrophils were significantly elevated at 3, 5, and 7 dpi and lymphocytes at 5 dpi, with both cell types returning to baseline by 21 dpi (Fig. 2C, D).
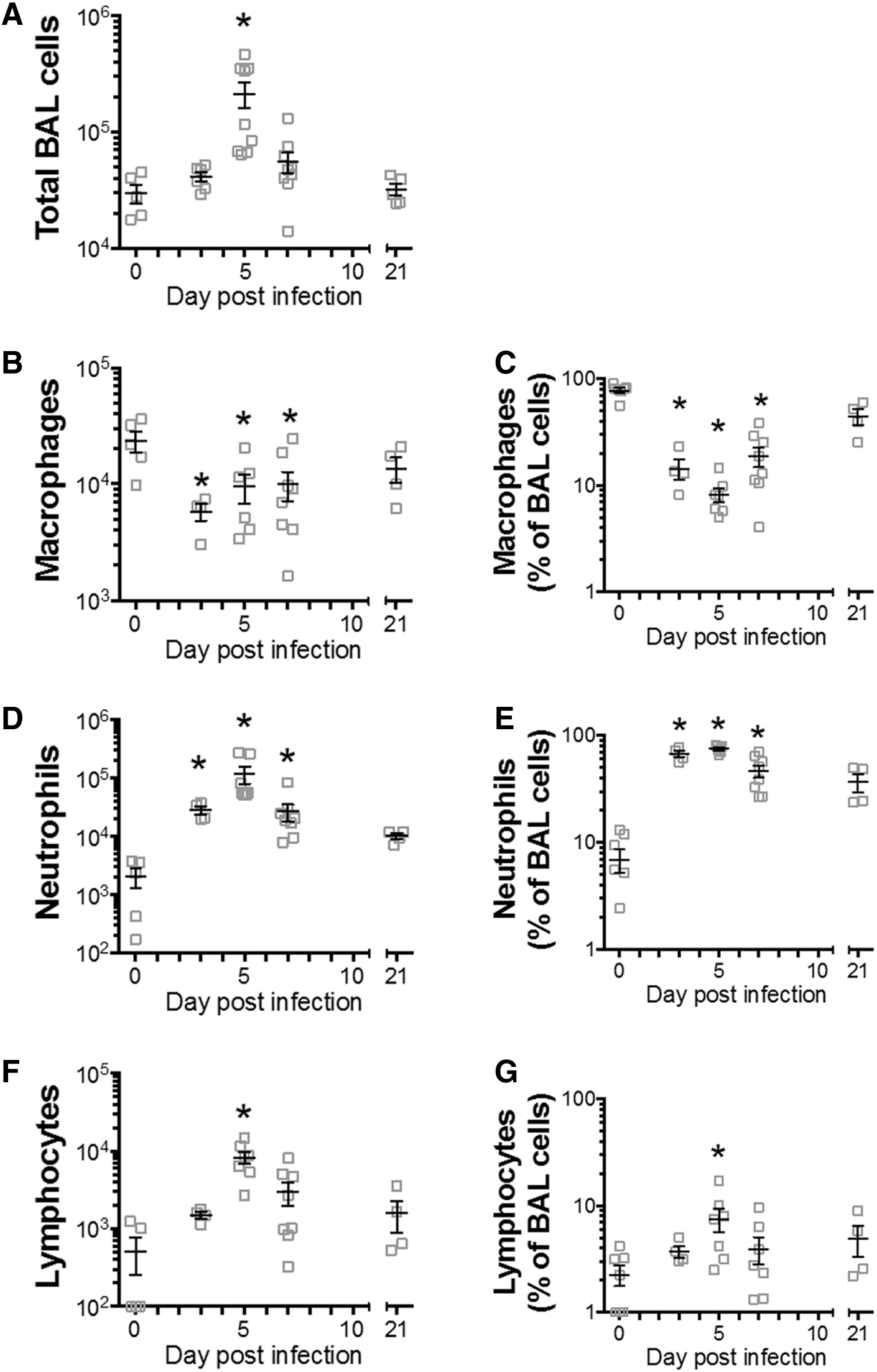
Neutrophilic inflammation in BAL fluid following RSV-RFP infection. BALB/c mice were infected with RSV-RFP, and BAL was obtained up to 21 dpi.
Analysis of the percentage frequency of each cell type in BAL showed a similar significant trend as for absolute numbers (Fig. 2C, E, and G). This analysis showed a very high percentage of neutrophils in BAL, reaching close to 100% of all BAL cells at 3 and 5 dpi, then declining, but still significantly elevated at 7 dpi, and remaining elevated, but no longer significantly elevated above baseline at 21 dpi (Fig. 2E).
Macrophage percentages decreased at 3, 5, and 7 dpi, returning to baseline by 21 dpi (Fig. 2C), while the percentage of lymphocytes was elevated at 5 dpi, returning to baseline at 7 and 21 dpi (Fig. 2G). Thus, RSV infection induced a significant inflammatory cell response in BAL at 3, 5, and 7 dpi that was dominated by neutrophils, which returned to baseline by 21 dpi.
Eosinophil cell numbers were not significantly different in infected compared with uninfected control mice at all time points (p > 0.05, data not shown).
Lung tissue leukocyte subtype responses following RSV-RFP infection
A significant increase in the percentage of lung tissue conventional DCs (cDCs) was noted on 7 and 21 dpi (Fig. 3A), with lung tissue macrophages also significantly increased at 21 dpi (Fig. 3B). Lung B cell numbers were significantly increased at 7 dpi. They remained elevated at 21 dpi, although this was not statistically significant (Fig. 3C).
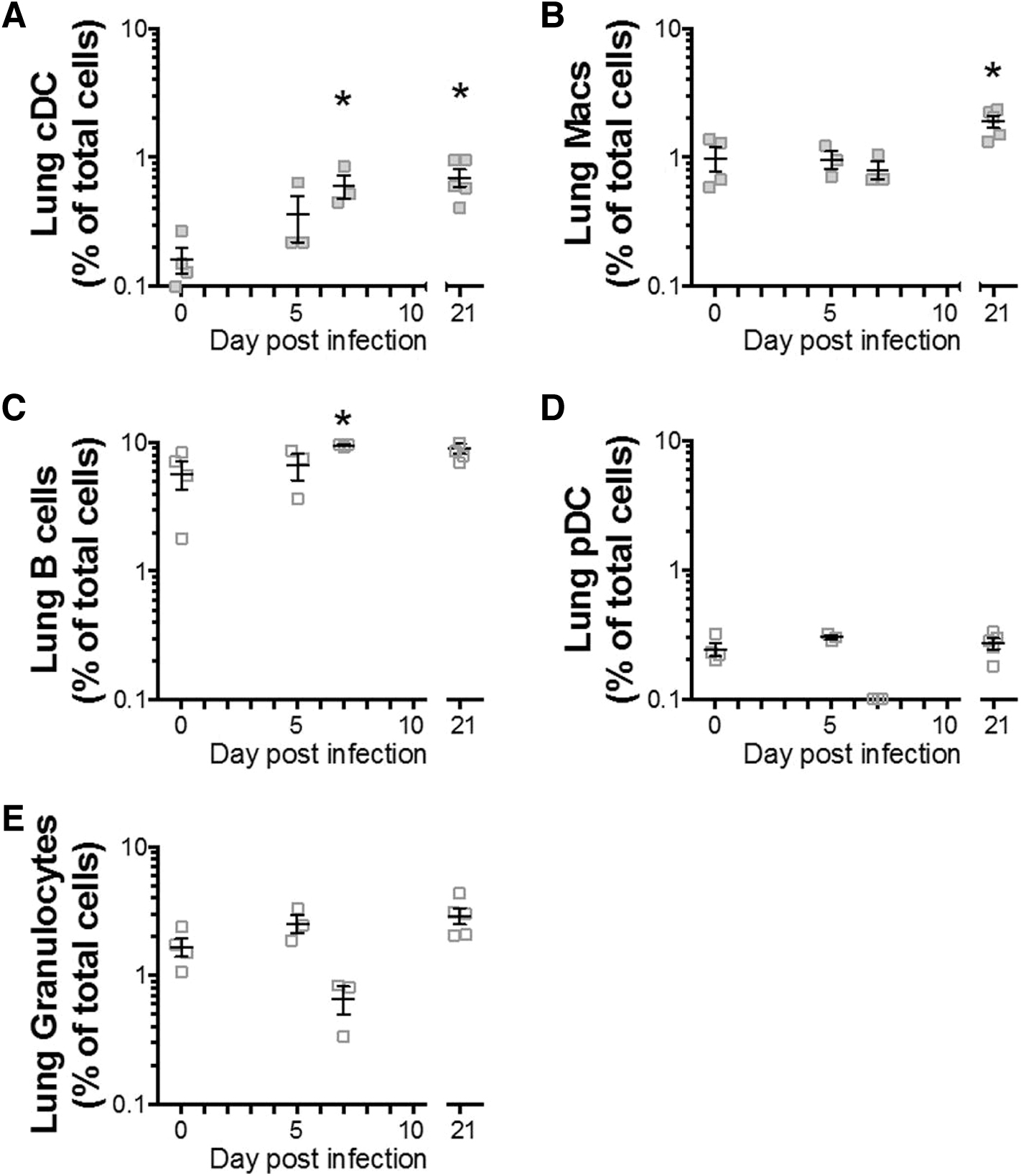
Lung tissue leukocyte subtype responses following RSV-RFP infection. BALB/c mice were infected with RSV-RFP, and lung single-cell suspension was analyzed by flow cytometry to identify
No increase in plasmacytoid DCs (pDCs) or granulocyte numbers was observed in lung tissue at any time point following infection (Figs. 3D, E). Thus, lung tissue APC subsets showed a dominant response to RSV in lung tissue at 7 dpi, with cDCs and macrophages remaining significantly elevated above baseline at 21 dpi.
Analysis of MHC II (I-A/I-E) expression as a marker of APC activation
Expression of MHC class II is an important regulator of APC function, being elevated upon activation and correlating with antigen-presenting capacity, indicating immune cell maturation (Villadangos et al., 2005). Analysis of the intensity of expression of the MHC class II marker, I-A/I-E (referred to as MHC II), on lung APC subsets after RSV-RFP infection revealed significantly increased expression on cDCs (Fig. 4A), macrophages (Fig. 4B), and B cells (Fig. 4C) at 21 dpi, with macrophages also showing an earlier increase at 7 dpi (Fig. 4B).

Increased expression of MHC class II (I-A/I-E) on lung APCs after RSV infection. BALB/c mice were infected with RSV-RFP, and lung single-cell suspension was analyzed by flow cytometry to identify MHC II expression on cDCs macrophages and B cells. MHC II MFI was significantly increased at 21 dpi in cDCs and B cells, whereas macrophage MHC II expression increased at 7 dpi and remained elevated at 21 dpi compared with baseline (0 dpi). Data are displayed as mean ± SEM; n = 6–9 per group. Statistical analysis was done using the Kruskal–Wallis test, with Dunn's multiple comparisons, compared with baseline (0 dpi), *p < 0.05. APC, antigen-presenting cell; MFI, mean fluorescence intensity.
Thus, the maturing impact of RSV infection on cDCs and other APC subsets appeared long-lasting following resolution of clinical symptoms.
Characterization of cell subsets of lung tissue expressing active RSV-RFP
We next characterized phenotypes of cells of lung tissue containing RSV using the marker RFP, expressed only by actively replicating the virus (Fig. 5A). The earliest time point at which RSV-RFP could be detected was 5 dpi in lung MHC II-positive APC populations (Fig. 5A), corresponding to the peak viral titer determined by qPCR (see Fig. 1C).
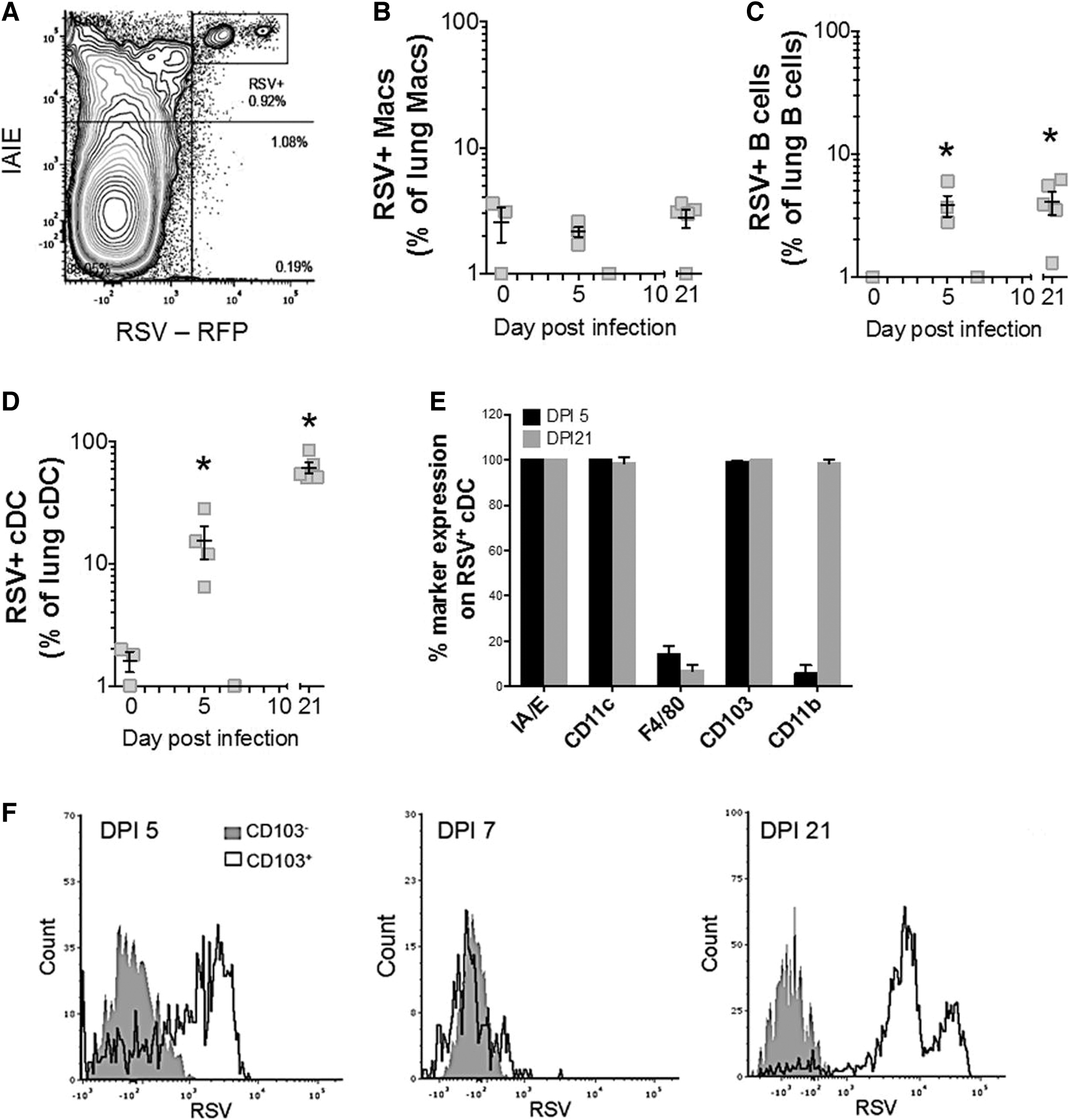
Long-term detection of RSV-RFP in lung cDCs. BALB/c mice were infected with RSV-RFP and lung single-cell suspensions were analyzed by flow cytometry to identify RSV-RFP-positive (+) cell subsets.
Although MHC II expression was noticeably high in macrophages at the time point after RSV infection (see Fig. 4B), low levels of RSV-RFP were observed in samples collected across all time points (Fig. 5B). RSV-RFP was detected in lung B cells (Fig. 5C) and cDCs (Fig. 5D) at 5 and 21 dpi of the lung MHC II-positive APC populations assessed (Fig. 5A).
Lung cDCs had the highest rates of infection, with ∼20% of cDCs expressing RSV-RFP at 5 dpi and ∼80% (Fig. 5D) with a significantly elevated RSV-RFP signal at 21 dpi (Table 1). RSV-RFP was not detected in lung pDCs at any time point (data not shown). Further analysis to determine the RSV-RFP-positive lung cDC phenotype revealed a cell subset with classical DC markers.
Mean Fluorescence Intensity of Respiratory Syncytial Virus Expressing Red Fluorescent Protein Signal on Lung Conventional Dendritic Cells, Macrophages, And B Cells from Respiratory Syncytial Virus-Infected Mice

These included high levels of CD11c and MHC II, with low levels of F480, as well as high levels of the intraepithelial cDC marker, CD103 (CD103+). These markers were also coexpressed with CD11b at 21 dpi (CD11b+) (Fig. 5E). In addition, RSV-RFP was only detected in lung CD103+CD11b+ cDCs at 5 and 21 dpi (Fig. 5F), with a subset showing high RSV-RFP levels in this population at 21 dpi only (Table 1).
Discussion
This is the first in vivo study to explore the pattern of active RSV replication in different immunological cells over the course of an acute infection and to demonstrate persistence of the virus in a subset of pulmonary DCs after the clinical symptoms of infection have resolved.
Cellular inflammation in this model was consistent with results from studies in human infants, in that the differential cell counts of BAL samples were dominated by intense neutrophilia (Everard et al., 1994; Smith et al., 2001). This response peaked at day 5 and resolved by 21 dpi. These results corresponded to the timing of changes in weight, clinical symptom severity scoring, and RSV viral load in the lung and again parallel the natural illness in humans.
Despite a neutrophil-dominated inflammatory response observed within the airways during acute illness, flow cytometry analysis showed that APCs were the dominant cells observed in lung tissue during acute infection. While inflammatory cell profiles observed from BAL began to resolve following 5 dpi, lung APC populations increased at 7 dpi, and most stayed elevated until 21 dpi. This analysis also showed elevated numbers of cDCs and macrophages, which remained significantly above baseline at 21 dpi.
Interestingly, granulocytes and pDC numbers in lung tissues assessed by flow cytometry were not significantly altered over the course of infection. The lack of a significant increase in granulocytes in lung tissue is attributable to the very rapid migration of neutrophils from arterioles through the epithelium to the luminal surface in response to the high levels of chemokines such as IL-8 being produced by the epithelium (Abu-Harb et al., 1999).
RSV replication in lung APC populations was biphasic, peaking at 5 and 21 dpi in cDCs and to a lesser extent in B cells. At 21 dpi, 80% of cDCs were found to carry RSV. Further subpopulation analysis of cDCs showed that CD103 expression was dominant at 5 dpi, with coexpression of CD11b observed at 21 dpi. In contrast, negligible RFP signal (indicating RSV) was detected in pDCs and macrophages.
These data suggest that RSV preferentially infects and replicates in cDCs and B cells and that RSV replication persisted in these cells well after the clinical symptoms of infection had subsided. While previous studies have reported elevated numbers of cDCs following RSV infection in mice, this is the first study to identify preferential harboring of RSV by CD103+CD11b+ cDCs in the lung following resolution of RSV-induced inflammation. This is also the first study to demonstrate coexpression of CD103 and CD11b in a single subpopulation of mucosal cDCs colocalized with RSV.
cDCs are known to exist in close proximity to the airway lumen (Stumbles et al., 2003) and are likely to be exposed to virus infecting the airway epithelium, from where they have been shown to migrate to regional lymph nodes. This migration pattern has been observed during the acute infection phase for CD103+ cells in murine models of RSV and influenza (Beyer et al., 2004; Kandasamy et al., 2013).
Further studies have demonstrated the propensity of CD103 cDCs for carrying and cross-presenting viral antigens, with a likely significant role in induction of immune memory responses (Beauchamp et al., 2010; Bedoui et al., 2009; Kandasamy et al., 2013; Schwarze, 2008). Recently, CD103+CD11b+ cDCs from the intestinal mucosa have been shown to play a role in IgG and IgA T cell-independent induced responses to flagellin (Flores-Langarica et al., 2018), clearly identifying this subpopulation as a link between adaptive immune responses to serological memory and systemic protection.
Given the weak, long-term immune responses, which characterize RSV infections throughout life, the influence of actively persisting RSV harbored by CD103+CD11b+ cDCs in poor immune memory responses warrants further investigation.
Certain viruses are known to infect and replicate in DCs, with a suggestion that some may have evolved to make use of these cells as a “Trojan horse” to subvert normal immune responses (Yewdell and Brooke, 2012). Data generated by our group suggest that RSV may be one of those viruses. Previously, we have demonstrated that RSV viral replication in vitro will continue in a proportion of monocyte-derived DCs for many months after infection before becoming latent (Jones et al., 2006).
We further demonstrated that this latent infection can be reactivated months later following exposure to nitric oxide (Hobson and Everard, 2008). In this study, RSV was shown to preferentially infect and replicate in cDCs and B cells, two key cells involved in generation of a long-term immune response, and this may be key to understanding why the human host does not appear to produce robust long-term antibody responses.
Persistence in the CD103+CD11b+ subset of DCs and B cells after resolution of acute infection and its associated inflammation suggests that the virus may also use these cells as a niche in which to persist within the host.
The results obtained in this study are consistent with previous in vitro observations of ongoing replication of RSV in DCs following exposure to the virus. Further studies, extending the time frame of observations, are required to determine whether the virus subsequently becomes dormant within these cells in vivo, and if so, can replication be reactivation at a later date, as has been shown in vitro.
Footnotes
Authors' Contributions
A.M.F. and N.S. completed all experiments. A.M.F., J.L.J., P.S., and D.S. completed the analysis. M.L.E. conceptualized the idea. A.M.F. and M.L.E. completed the article, with review and editing done by all authors.
Ethics Approval
All experiments were approved by the Telethon Kids Institute Animal Ethics Committee (permit No. 266) and carried out in accordance with the recommendations of the Australian Code for the Care and use of Animals for Scientific Purposes, 8th edition (2013).
Availability of Data and Materials
The corresponding author will provide any further information regarding materials or datasets upon reasonable request.
Preprint
An earlier draft of this article was posted as a preprint at Authorea (DOI: 10.22541/au.165579307.76212895/v1).
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by the McCusker Medical Research Foundation and Perth Children's Hospital Foundation.
Supplementary Material
Supplementary Figure S1
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.