
Abstract
Current evidence shows higher production of cytokines and antibodies against severe acute respiratory coronavirus 2 (SARS-CoV-2) in severe and critical cases of Coronavirus Disease 2019 (COVID-19) in comparison with patients with moderate or mild disease. A recent hypothesis proposes an important role of genotoxicity and cytotoxicity in the induction of the cytokine storm observed in some patients at later stages of the disease. Interestingly, in this study, we report significantly higher levels of interleukin (IL)-1β, IL-6, MCP-1, and IL-4 cytokines in mild COVID-19 patients versus severe cases, as well as a high frequency of karyorrhexis (median [Me] = 364 vs. 20 cells) and karyolysis (Me = 266 vs. 52 cells) in the mucosal epithelial cells of both groups of patients compared with uninfected individuals. Although we observed higher levels of anti-SARS-CoV-2 IgM and IgG antibodies in COVID-19 patients, IgM antibodies were significantly higher only in mild cases, for the N and the S viral antigens. High levels of IgG antibodies were observed in both mild and severe cases. Our results showed elevated concentrations of proinflammatory and anti-inflammatory cytokines in mild cases, which may reflect an active innate immune response and could be related to the higher IgM and IgG antibody levels found in those patients. In addition, we found that SARS-CoV-2 infection induces cytotoxic damage in the oral mucosa, highlighting the importance of studying the genotoxic and cytotoxic events induced by infection and its role in the pathophysiology of COVID-19.
Introduction
The infection by the severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2), the causal agent of Coronavirus Disease 2019 (COVID-19), has been characterized by stages of infection, immune response, and recovery. An aberrant hyperactivation of the immune response has been described in severe cases of COVID-19 and other viral infections (Fajgenbaum and June, 2020; Mazzoni et al., 2021; Montazersaheb et al., 2022; Parasher, 2021). Particularly, severe cases of COVID-19 develop an acute respiratory distress syndrome (ARDS) because of a cytokine storm characterized by the overproduction of several proinflammatory cytokines, coupled with different COVID-19 risk factors (age and comorbidities). In addition, increased levels of ferritin, D-dimer, C reactive protein, and procalcitonin, have been linked to a worse outcome (Boechat et al., 2021; Lo Tartaro et al., 2022).
On the other hand, the early production of cytokines is necessary for an adequate immune response against the virus. The innate immune response to viral infections can be initiated through the recognition of viral, but also host-derived nucleic acids (Chen et al., 2022b; Imre, 2020; Verburg et al., 2022; Zhang and Myers, 2018), which can be produced due to the ability of the virus to induce genotoxicity and cytotoxicity in the host cell (Ribeiro et al., 2018; Roberts et al., 2007; Zhang and Myers, 2018). These processes may contribute to the production of cytokines through the activation of inflammatory pathways (Gu et al., 2021; Murthy et al., 2020; Ren et al., 2021). Moreover, genotoxicity and cytotoxicity can be triggered in other inflammatory conditions like diabetes and obesity, known comorbidities of COVID-19 disease, inducing the production of proinflammatory cytokines (Oduro et al., 2022; Seifi et al., 2014; Torres-Bugarín et al., 2009).
Evidence on the genotoxic and cytotoxic effects of SARS-CoV-2 infection is scarce, however, some studies reported the production of micronuclei (MN) or cytopathic markers (karyorrhexis, and karyolysis) in epithelial cells of the oral cavity and peripheral blood cells from infected individuals with SARS-CoV-2 (Berber et al., 2021; Pinto et al., 2021).
Cytokines released by innate immune cells such as monocytes, macrophages, and NK cells activate other cells. Upon disease progression, T cells and specific antibodies released by B lymphocytes help limit virus propagation and induce immune protection (Li et al., 2006; Wang et al., 2020). Timely regulatory mechanisms, which include regulatory cytokines, such as interleukin (IL)-4, IL-10, and transforming growth factor (TGF)-β are essential to avoid overwhelming responses such as the pathogenic cytokine storm that may cause tissue damage and organ failure (Tisoncik et al., 2012). Thus, the correct balance of pro- and anti-inflammatory cytokines is key to the outcome of an inflammatory process.
Similar to the early cytokine production triggered by the innate defense system (Diamond and Kanneganti, 2022), the adaptive immune response in charge of the specific antibody production against the virus has been linked to COVID-19 severity (Primorac et al., 2022; Sette and Crotty, 2021). Some studies report higher titers of anti-SARS-CoV-2 IgA, IgM, and IgG antibodies coupled to elevated levels of proinflammatory cytokines, particularly IL-6, in severe versus mild cases of COVID-19 (Codina et al., 2021; Latifi-Pupovci et al., 2022). Consequently, higher antibody and cytokines levels have been proposed as predictors of COVID-19 outcomes. In contrast, early elevated cytokine levels have been suggested as an indicator of an active immune response, which can promote higher antibody production at a later stage of infection by the recruitment of monocytes, macrophages, and T cells (Jing et al., 2022). A regulated cellular and humoral immune response results in the elimination of the virus without concomitant immunopathology.
The proposed positive feedback between genomic instability and cytokine and antibody production during SARS-CoV-2 infection, suggests the presence of genotoxicity and cytotoxicity in COVID-19. The present study aimed to evaluate the production of cytokines and anti-SARS-CoV-2 antibodies, as well as the genotoxicity and cytotoxicity in unvaccinated patients with mild and severe COVID-19.
Materials and Methods
Study design, data collection, and ethics statement
Between April to November 2020, 53 participants, 16 mild and 22 severe patients with SARS-CoV-2 infection confirmed by quantitative real time-polymerase chain reaction (qRT-PCR) at the public Hospital General “Dr. Manuel Gea Gonzalez” (HGMGG) and 15 clinically healthy adult uninfected volunteers with a SARS-CoV-2 negative test were invited to enroll in the present study. Uninfected individuals were included as the reference group. Each case was classified as mild or severe according to the Chinese Clinical Guide for COVID-19 Pneumonia Diagnosis and Treatment published on March 4, 2020 (ACC, 2020). This classification was confirmed using the most recent guidelines established for COVID-19 severity stratification by the Centers for Disease Control and Prevention (CDC, 2023).
Age (years) was analyzed as the percentage distribution of participants by deciles, and further classified as older adults (≥60 years) to describe the distribution of noncommunicable diseases. Obesity distribution was obtained according to World Health Organization (WHO) standards for body mass index (BMI, kg/m2): BMI ≥30 obesity, 25–29.9 overweight, 18.5–24.9 normal, or <18 underweight/malnourished. The participants referred comorbidities such as hypertension, diabetes, and current smoking status.
Mild and severe COVID-19 groups consisted of unvaccinated subjects, whose demographic and clinical information was collected in electronic health records and assigned a unique ID code to each registry to prevent data duplication and anonymize the identity of the participants. The study was registered under the internal number IN216121 and was approved by the Research and Ethics Committees of the HGMGG (reference no. 12-26-2020). We obtained informed consent from all participants before recruitment and all procedures were conducted under the Declaration of Helsinki and the Federal Law on Personal Data Protection in Mexico.
Serum samples and cytokine assay
Serum samples were obtained as previously described (Tuck et al., 2009) and stored in aliquots at −80°C until use. The concentrations of interferon (IFN)-γ, IL-1β, IL-6, tumor necrosis factor (TNF)-α, IP-10, MCP-1, IL-2, IL-12p70, IL-8, IL-17A, IL-4, IL-10, and TGF-β were analyzed using the LEGENDplex Hu Essential Immune Response Panel (13-plex) (BioLegend, San Diego, CA) according to the manufacturer's instructions.
SARS-CoV-2 IgG and IgM detection
For the evaluation of the production of IgG and IgM antibodies against the SARS-CoV-2, we used a recombinant SARS-CoV-2 Nucleocapsid protein (N) and a previously reported peptide from the Spike protein subunit 2 (S2) (Li et al., 2021b). IgM and IgG (anti-S2 and anti-N) levels were measured in serum samples as previously described (Avila et al., 2020; Li et al., 2021b; Sun et al., 2020a). The cutoff point was determined as the mean of the optical density of samples from unvaccinated volunteers plus two standard deviations. Consequently, the results were expressed as an ELISA index (EI), where EI = samples absorbance/absorbance of the cutoff point. EI >1 values were considered positive.
Oral epithelium samples, genotoxicity, and cytotoxicity assays
Genotoxic and cytotoxic damage due to SARS-CoV-2 infection was assessed in 1,000 oral cavity epithelial cells by Acridine Orange staining. In addition to MN, other nuclear abnormalities related to genotoxicity and cytotoxicity like karyorrhexis and karyolysis were also evaluated (Thomas et al., 2009).
Statistical analysis
We evaluated the normal distribution of the data by the Shapiro–Wilk test, showing a non-normal distribution; thus, values were expressed as the median (Me) [interquartile range]. Demographic and clinical factors, such as age, sex, smoking status, and comorbidities (diabetes, hypertension, and obesity) showed a confounding effect in our exploratory analysis, which is expected given the effect of these factors in COVID-19 disease (Barek et al., 2020; Chenchula et al., 2023). The significance of the differences in the levels of blood analytes, cytokines, genotoxicity/cytotoxicity, and antibodies, between SARS-CoV-2− and SARS-CoV-2+ or uninfected volunteers, mild, and severe patients, was performed using an adjusted quantile regression test.
Demographic data were described as percentages and were compared by the chi-square test. The database was constructed in Excel and the analyses were performed using SPSS 23 (SPSS, Inc., Chicago, IL) and STATA 17 (StataCorp., College Station, TX), while graphics were designed with GraphPad Prism 9 (GraphPad Software, Inc., San Diego, CA). We considered a p ≤ 0.05 value as a statistically significant threshold in all tests.
Results
Demographic and clinical characteristics of COVID-19 patients
During the first wave of COVID-19, we collected 38 samples from participants with SARS-CoV-2+ PCR tests; of these, 27 (71.1%) were male. Significant differences between SARS-CoV-2− and SARS-CoV-2+ individuals were observed in the 30–39 (p < 0.0001) and in the ≥60 (p = 0.001) age groups. Most SARS-CoV-2- individuals were in the 20–29 (3/15 = 21.4%) and 30–39 (8/15 = 57.1%) age groups, whereas SARS-CoV-2+ patients were older adults (21/38 = 56.8%). In addition, 21 patients (56.8%) suffered from one or more of the following comorbidities, such as hypertension (14/38 = 36.8%), diabetes (9/38 = 23.7%), obesity (7/38 = 18.4%), and 35.5% were current smokers (11/31). Significant differences between infected and uninfected individuals were only evident in the hypertension (p = 0.005) and smoking status (p = 0.009) groups (Table 1).
Demographic Characteristics of the Study Population
Bold values indicates statistically significant data.
Notes: age is expressed in years, while age groups were defined by deciles, except for those ≥60. p-values for X 2.
Missing values = 1 in SARS-CoV-2− and 1 in SARS-CoV-2+.
It should be noted that some categories can be counted more than once and there is one missing value.
p-Values correspond to the comparison between SARS-CoV-2+ versus SARS-CoV-2− infected subjects with or without some comorbidity.
Missing values for infected subjects = 7.
SARS-CoV-2, severe acute respiratory syndrome coronavirus 2.
Clinical parameters in COVID-19 patients such as oxygen saturation, D-dimer, and creatinine median concentration among others are shown in Table 2. Several analytes evaluated showed significant differences between mild and severe COVID-19 cases. We found lower concentrations of total bilirubin (TB; 0.7 vs. 0.9 mg/dL, p = 0.024), aspartate aminotransferase (AST; 33 vs. 41 U, p = 0.027), and lactate dehydrogenase (LDH; 372.0 vs. 423.5 U, p = 0.029), and significantly higher concentrations of troponin (0.04 vs. 0.01 U, p = 0.012) and leukocyte counts (10.6 vs. 8.0 109/L, p = 0.012) in severe versus mild COVID-19 cases. Compared with patients that survived, those who died from COVID-19 had significantly higher concentrations of alanine aminotransferase (ALT; 28.0 vs. 27.0 U, p = 0.013), AST (40 vs. 35 U, p = 0.020), and platelets (266 vs. 209 103/μL, p = 0.041), while hemoglobin concentrations were lower (14.0 vs. 15.3 U, p = 0.018).
Clinical Parameters of Coronavirus Disease 2019 Patients According to Disease Severity and Outcome
Bold values indicates statistically significant data.
Notes: data are expressed as median [IQR]. p-values were calculated by the quantile regression test adjusted by age, sex, and comorbidities (diabetes, hypertension, and obesity) since the Shapiro–Wilk test showed a non-normal distribution of the data.
Missing values for infected subjects: 3.
ALT, alanine aminotransferase; AST, aspartate aminotransferase; CKBM, creatine kinase; DB, direct bilirubin; IB, indirect bilirubin; IQR, interquartile range; LDH, lactate dehydrogenase; TB, total bilirubin.
Serum cytokine levels in mild and severe disease
Although SARS-CoV-2+ patients showed higher concentrations of some cytokines, such as IFNγ (28.0 vs. 9.7 pg), IL-6 (181.2 vs. 62.2 pg), IP-10 (891.3 vs. 192.9 pg), and IL-10 (20.6 vs. 7.1 pg) compared with uninfected individuals, the differences did not reach statistical significance (Supplementary Table S1). However, when analyzed by disease severity, we observed statistically significant higher concentrations of IL-1β (24.2 vs. 5.6 pg, p = 0.017), IL-6 (231.3 vs. 163.0 pg, p = 0.023), MCP-1 (105.2 vs. 74.1 pg, p = 0.046), and the anti-inflammatory cytokine IL-4 (102.8 vs. 31.8 pg, p = 0.031) in the mild COVID-19 group. Moreover, when comparing severe COVID-19 cases to the uninfected volunteers, we found lower concentrations of IL-1β (5.6 vs. 9.3 pg, p < 0.0001), TNF-α (5.3 vs. 8.9 pg, p < 0.0001), IL-8 (6.2 vs. 11.4 pg, p < 0.0001), and IL-17A (5.7 vs. 20.8 pg, p = 0.024), and higher levels of IFN-γ (16.9 vs. 9.7 pg, p = 0.001), IP10 (737.2 vs. 192.9 pg, p = 0.003), and MCP-1 (74.1 vs. 71.1 pg, p = 0.049) in the severe COVID-19 patients (Fig. 1).
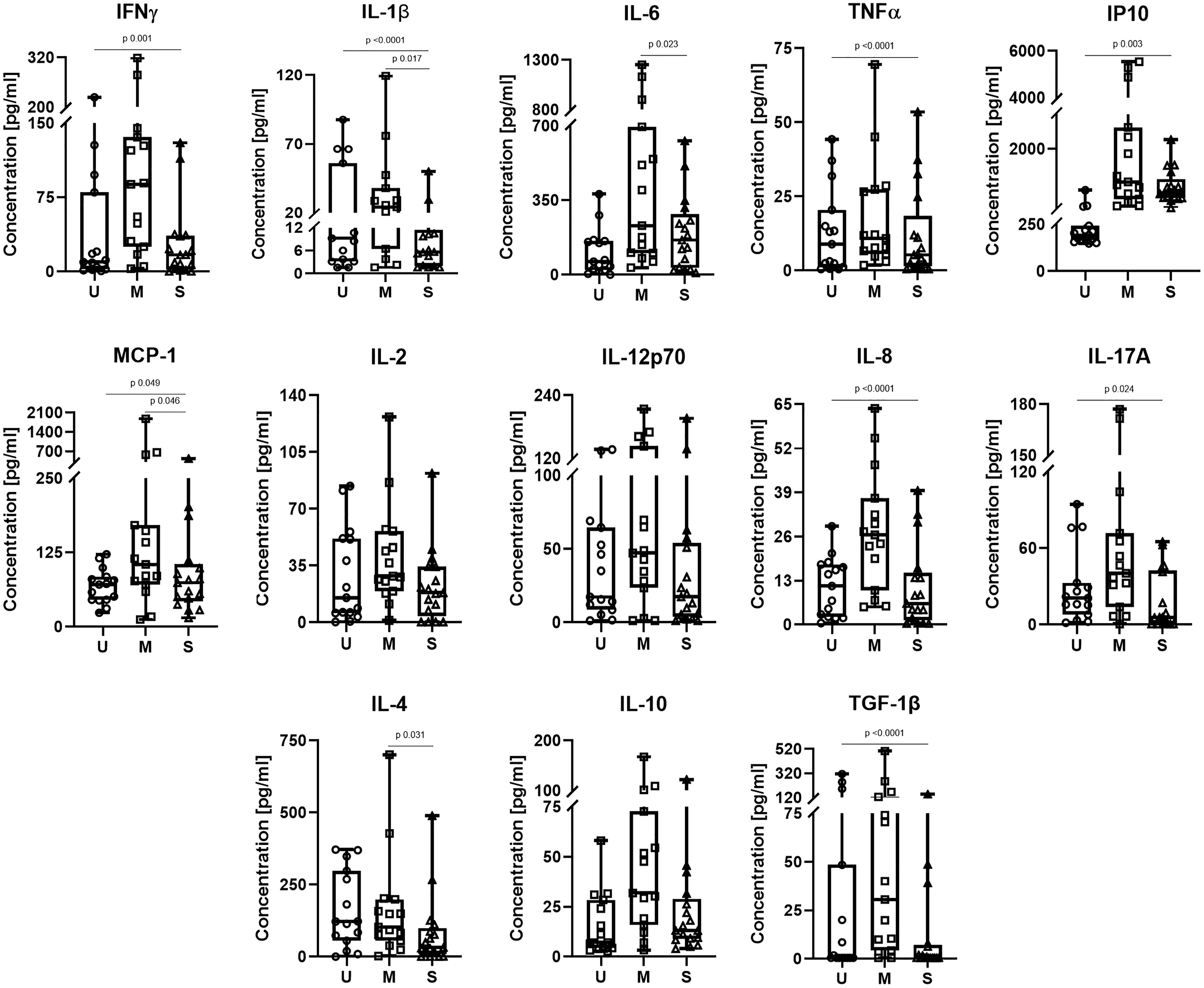
Comparison of serum cytokines between uninfected volunteers (U), mild (M), and severe (S) COVID-19 patients. Box and whiskers plot shows the median concentrations of each cytokine. p-Values were calculated by the quantile regression test adjusted by age, sex, and comorbidities (diabetes, hypertension, and obesity) due to a non-normal distribution of the data. Missing values = 6. COVID-19, Coronavirus Disease 2019.
Genotoxic and cytotoxic markers in COVID-19 patients
For the evaluation of the genotoxic and cytotoxic effects of SARS-CoV-2 infection, we measured the frequency of several biomarkers in epithelial cells of the oral mucosa. Both mild and severe COVID-19 patients showed a significantly lower frequency of normal cells without signs of cellular damage (uninfected volunteers: Me = 903 vs. COVID-19 patients: Me = 403, p = 0.004). In contrast, compared with SARS-CoV-2− volunteers a higher frequency of karyorrhexis (Me = 364 vs. 20, p = 0.002) and karyolysis (Me = 266 vs. 52, p = 0.689) was found in COVID-19 patients (Fig. 2; Supplementary Table S2). Similar levels of karyorrhexis (mild = 364; severe = 353) and karyolysis (mild = 276; severe = 232) were observed regardless of disease severity (Fig. 2). The frequency of cells with MN and other genomic biomarkers was not significantly different between mild versus severe COVID-19 patients or uninfected volunteers (Supplementary Table S2).
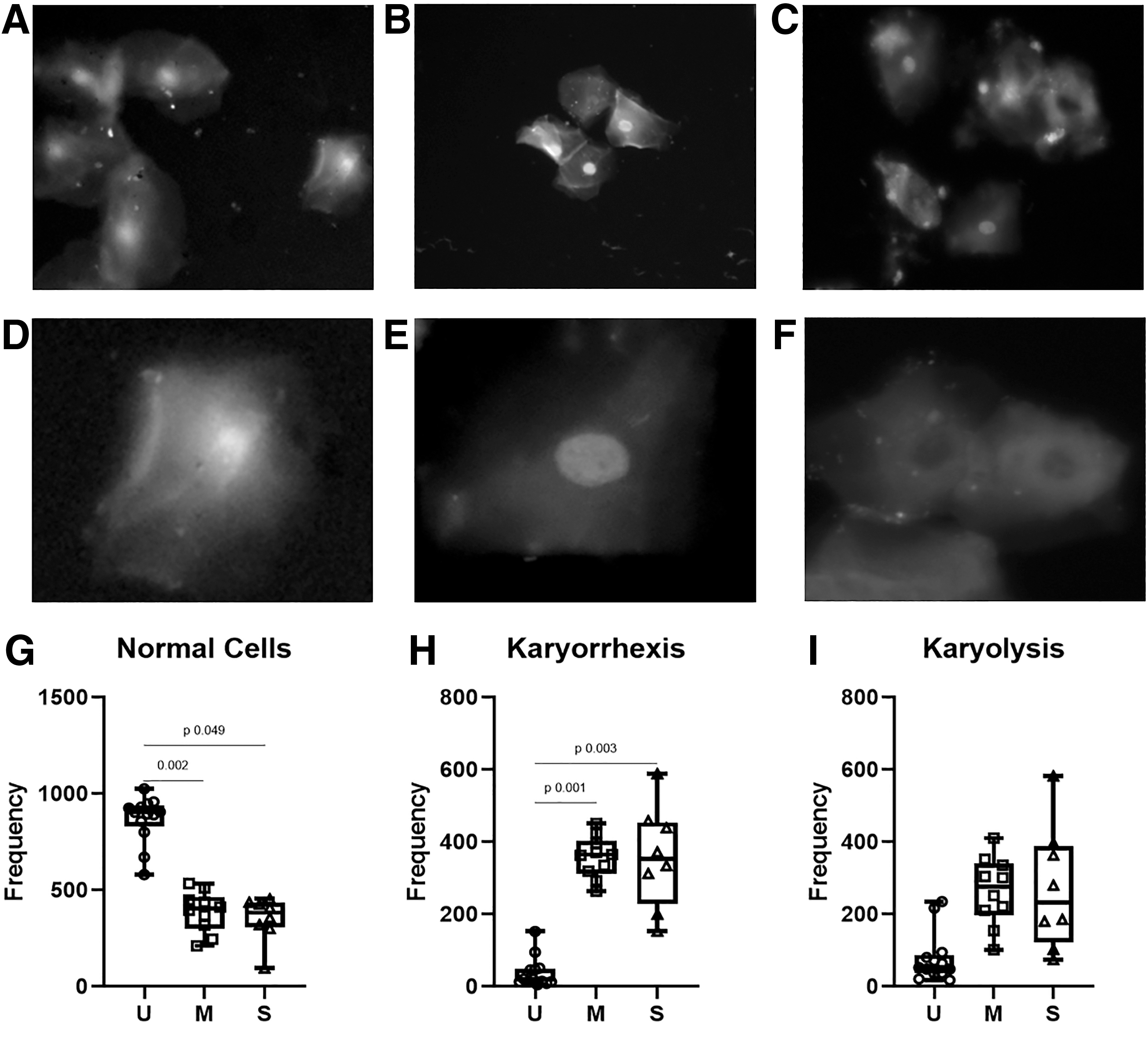
Nuclear abnormalities in uninfected volunteers (U), mild (M), and severe (S) COVID-19 patients. Representative pictures of cells from
IgG and IgM anti-SARS-CoV-2 antibodies in mild and severe COVID-19 patients
To evaluate the production of anti-SARS-CoV-2 antibodies, specific IgM and IgG antibody levels against the recombinant N protein and S2 were measured and the EI was calculated. As expected, infected subjects had higher IgM and IgG antibody levels against SARS-CoV-2 antigens than uninfected volunteers. We observed significant differences for anti-N IgG (2.5 vs. 0.4, p < 0.001) and anti-S2 IgG (2.8 vs. 0.8 p < 0.001). IgM levels did not reach statistical significance as compared with the uninfected individuals (Supplementary Table S3). We found IgM and IgG antibodies against N in 21 patients (58.3%), while only IgG antibodies were detected in 41.6% of infected individuals. Similarly, both isotypes against S2 were detected in 38.9% (14/36) of serum samples, and 47.2% showed IgG antibodies but not IgM (Supplementary Table S3). When we separated individuals with mild and severe disease, we found a significantly higher EI for IgM antibodies against the N viral antigen in patients with mild disease as compared with uninfected volunteers (1.41 vs. 0.46, p = 0.041).
For IgG antibodies against N, we observed a significantly higher EI in patients with both mild and severe disease as compared with uninfected volunteers (mild = 2.54; severe = 2.48 vs. uninfected volunteers = 0.40, p < 0.001) (Fig. 3). A similar result was found for anti-S2 IgM (mild = 1.07 vs. uninfected volunteers = 0.43, p = 0.006) and anti-S2 IgG levels (mild = 2.65; severe = 2.85 vs. uninfected volunteers = 0.76, p < 0.0001) (Fig. 3).

Serum levels expressed as EI of IgG and IgM antibodies against nucleoprotein (N) or Spike-subunit 2 (S2) SARS-CoV-2 proteins in mild (M) and severe (S) COVID-19 patients. Uninfected volunteers (U). p-Values were calculated by the quantile regression test adjusted by age and sex. Missing values = 2. EI, ELISA index; SARS-CoV-2, severe acute respiratory syndrome coronavirus 2.
Discussion
Since the rise of the COVID-19 pandemic in 2019, several studies have reported different aspects of the pathophysiology of the disease. Particularly, the development of complications in severe and critical COVID-19 patients, such as ARDS or sepsis, have been described and associated with an aberrant immune response and deregulated production of cytokines (Chen et al., 2021; Wilson et al., 2020). A recent hypothesis suggests that the release into the cytoplasm of nuclear DNA contained in MN (genotoxicity marker), could feed the production of proinflammatory cytokines found in COVID-19 cases (Kirsch-Volders and Fenech, 2021). Few studies have reported the genotoxic or cytotoxic effects of SARS-CoV-2 in patient samples. However, none describe cytokines or anti-SARS-CoV-2 antibody levels or associate them with disease severity. Consequently, this study aimed to assess the production of cytokines and anti-SARS-CoV-2 antibodies, as well as the genotoxicity and cytotoxicity induced by SARS-CoV-2 infection in unvaccinated people during the first wave of the pandemic.
In the samples obtained from 38 unvaccinated SARS-CoV-2+ patients, we found a higher proportion of males and elderly subjects with comorbidities like hypertension, diabetes, obesity, and smokers. These observations are supported by current evidence, which shows that these demographic and clinical characteristics are risk factors for COVID-19 (Bigdelou et al., 2022; Kharroubi and Diab-El-Harake, 2022).
With respect to laboratory parameters, we observed higher levels of TB and AST in mild cases and lower levels of LDH as compared with severe cases. In patients who died from COVID-19, higher levels of AST and ALT were found. Several studies have shown a positive association between abnormal levels of hepatic function markers (bilirubin, AST, ALT, and LDH) with disease severity and a worse disease outcome (Chen et al., 2022a; Ouali et al., 2020; Saini et al., 2022; Salık et al., 2021). Hepatocytes (2.6%) and cholangiocytes (59.7%) express Angiotensin-Converting Enzyme 2 (ACE2) (Sonzogni et al., 2020), making these two cell types potential SARS-CoV-2 targets (Fiel et al., 2021). Indeed, COVID-19 patients show altered levels of AST and ALT resulting in the hepatic dysfunction (Ahmad et al., 2021). Even though SARS-CoV-2 can directly damage hepatocytes, an additional hypothesis suggests that liver damage, denoted by the increase of hepatic enzyme levels, may result in exacerbated cytokine production (Ali et al., 2021; Elemam et al., 2023; Taneva et al., 2021).
Previous studies have reported higher serum levels of IL-6 in several hepatic diseases (Schmidt-Arras and Rose-John, 2016), as well as a positive correlation between IL-6 and AST levels in hospitalized COVID-19 patients (Effenberger et al., 2021). Nevertheless, in patients with mild or moderate manifestations, some reports evidence a transient increase in hepatic enzymes AST and ALT, which return to normal after the patient recovers (Zhang et al., 2020). Additionally, we found higher levels of troponin and leukocytes in patients with severe COVID-19 and higher platelet numbers in people who died from the disease. Current evidence shows that myocardial injury and aberrant blood cell counts have been commonly found in severe cases and COVID-19-associated deaths (García de Guadiana-Romualdo et al., 2021; Imazio et al., 2020; Palladino, 2021). Further studies are needed to explain the mechanisms underlying the systemic inflammation triggered by SARS-CoV-2 infection that leads to multiple organ damage, including hepatic dysfunction (Ali et al., 2021).
Cells from the oral mucosa express high levels of ACE2 and Transmembrane Serine Protease 2 (TMPRRS2) and may represent a direct entry route for SARS-COV-2 (Huang et al., 2021; Okui et al., 2021; Sawa et al., 2021; Xu et al., 2020). Moreover, epithelial cells from oral mucosa have been widely used to study the rate of division of proliferating cells, genomic stability, and susceptibility for cell death (Bonassi et al., 2011). Genotoxic and cytopathic markers in cells from the oral mucosa and peripheral blood cells from COVID-19 patients have been described (Berber et al., 2021; Pinto et al., 2021). Overall, our results are consistent with this evidence since we found lower normal cells coupled to a higher frequency of cells undergoing karyorrhexis or karyolysis in the epithelial cells of the oral cavity of COVID-19 patients. During karyorrhexis, chromatin-filled granules are spread in the cytoplasm while karyolysis, results in nucleus loss. Both processes may be a result of increased genotoxicity (Altoukhi et al., 2021).
In our study, we did not find significantly higher MN frequencies in COVID-19 patients. This finding could be explained by high turnover of the oral epithelium and the presence of elevated levels of karyorrhexis and karyolysis that represent later stages of cytotoxic damage. Likewise, nonsignificant differences may be due to adjustments for age, sex, and comorbidities in our data, as current studies that report the frequency of genotoxicity and cytotoxicity in COVID-19 patients (Berber et al., 2021; Pinto et al., 2021) did not establish whether they made adequate adjustments for these confounding factors, which can significantly contribute to higher levels of MN, karyorrhexis, and karyolysis (Ferraz et al., 2016; Seifi et al., 2014; Torres-Bugarín et al., 2009). Therefore, if the hypothesis regarding the association between genotoxicity, cytokine production, and severity is correct, it would be interesting to evaluate the genotoxicity and cytotoxicity in a larger cohort to establish if such association exists. A study including the patients' follow-up would provide information on the dynamics of these markers during the disease.
The finding that both mild and severe cases of COVID-19 have similar levels of karyorrhexis and karyolysis suggests that cytopathic viruses, such as SARS-COV-2, induce cytotoxicity upon infection. Thus, we would expect to have similar levels of karyorrhexis and karyolysis in infected individuals because of viral infection independent of disease severity. However, since mild cases produce higher levels of both pro- and anti-inflammatory cytokines, this could be an indication of a more regulated immune response, which would result in controlled inflammation that is necessary for the elimination of the virus (Li et al., 2021a; Li et al., 2020). Accordingly, several studies show the induction of cytotoxicity in infections by respiratory viruses, such as, influenza, respiratory syncytial virus, rhinovirus, and SARS-CoV-2 (Berber et al., 2021; Bossios et al., 2005; Montgomery et al., 2020; Pinto et al., 2021; Welliver et al., 2008). For example, a COVID-19 study reports the significant induction of cytotoxicity in SARS-CoV-2+ patients, but no differences in mild versus severe cases (Berber et al., 2021).
Thus, it is possible that the cytotoxicity induced by SARS-CoV-2 is associated with the inflammatory process derived from the infection at early stages, but not with the severity of the disease, in which other factors such as the presence of comorbidities have been clearly associated.
In the present study, we measured the concentration of 13 cytokines in the serum of COVID-19 patients, finding higher concentrations of 11 cytokines in SARS-CoV-2-infected individuals, but none reached statistical significance probably because of the small sample number. Also, we adjusted cytokine concentration for age, sex, and comorbidities, confounding factors that can induce elevated cytokine levels and therefore alter cytokine measurements in patients with COVID-19 (Gouda et al., 2018; Klein and Flanagan, 2016; Rea et al., 2018; Zatterale et al., 2020). Interestingly, when grouping according to disease severity, we found that patients with mild disease had higher circulating cytokine levels. This finding seems to contrast current studies that report higher production of several cytokines like IFNγ, IL-1β, IL-6, IP-10 IL-8, IL-17A, and IL-10 in severe cases of COVID-19 (Cabaro et al., 2021; Chang et al., 2022; Kalinina et al., 2022; Zeng et al., 2020).
Nevertheless, supporting our findings, other studies have reported that patients with mild disease have higher cytokine levels (Ghazavi et al., 2021), as well as higher titers of anti-SARS-CoV-2 antibodies, as a consequence of the efficient recruitment of different immune cells by the regulated secretion of several cytokines at an early stage of the illness (Jing et al., 2022).
Additionally, a single-cell sequencing assay demonstrated marked differences between the immune cell composition of mild and critical intubated COVID-19 patients (Wauters et al., 2021). The authors reported an active cytokine production in both groups; however, they found a more regulated response in mild patients characterized by the expression of genes associated with the IFN and autoregulatory responses. Particularly, higher levels of macrophages and CD8+ lymphocytes with elevated expression of IFN type I and II were found in mild cases. In contrast, immune cells from critical patients showed reduced effector function due to persistent proinflammatory stress. Furthermore, early studies report higher viral loads in mild cases in comparison to severe patients (Liu et al., 2020), while recent evidence associated higher viral loads with age, symptoms (gastrointestinal and respiratory), vaccination status, and mortality (Hastie et al., 2023), highlighting the importance of adjusting for comorbidities as was the case in the present study.
Higher cytokine levels in mild COVID-19 may reflect an active immune response that is able to control the infection, while in severe and critical cases this response is diminished due to the stress to which the immune system is subjected to. A not mutually exclusive explanation is the development of a deregulated response that results in immunopathology.
The balance between Th1 and Th2 responses is important during COVID-19. However, T regs and other tolerogenic cells, as well as the cytokines they release are key players in determining the outcome of the inflammatory process. Cytokines drive and are involved in this balance. Innate immune mechanisms help identify and initiate further adaptive processes, such as a Th1 response that helps limit viral infections. An early Th1 response is crucial for limiting SARS-CoV-2 infection (Gil-Etayo et al., 2022). However, if this response extends and lacks regulatory mechanisms, poor outcomes may ensue, characterized for example, by a cytokine storm (Lucas et al., 2020). Thus, it is possible that recovery from COVID-19, like other infections, depends on the balance of pro- and anti-inflammatory cytokines, resulting in a normal regulated immune response in due time. Accordingly, Fathi et al., showed that during disease and the recovery phase, Th1/Th2 cytokines change to provide a regulated response in recovered patients (Fathi et al., 2020).
The positive feedback between genotoxicity/cytotoxicity and cytokine release can also contribute to a good production of anti-SARS-CoV-2-specific antibodies. A previous study reported that early production of several cytokines, particularly, IL-6 in the first 2 weeks after infection with SARS-CoV-2 leads to higher specific IgG antibody titers (Jing et al., 2022). Although we do not have information on disease onset, we did find higher cytokine levels in mild cases, including IL-6, as well as elevated levels of anti-SARS-CoV-2 IgM and IgG antibodies in both mild and severe COVID-19 patients. Moreover, we found higher levels of anti-S2 IgM and anti-N IgG antibodies in mild cases, similar to those recently reported (Cervia et al., 2022). Therefore, it is plausible that a higher frequency of genotoxic and/or cytotoxic events may indirectly favor the humoral response through the production of cytokines in the early stages of the disease, resulting in virus neutralization, the recruitment of various types of immune cells at later stages and antibody-mediated cytotoxicity that limit viral shedding.
Finally, our study has some limitations that are inherent to cross-sectional studies, mainly the sample size, particularly in the measurement of genotoxic and cytotoxic events. Nevertheless, in our samples, we found the same tendency of demographic and clinical characteristics described in other studies with a larger cohort. Additionally, due to restrictive measures implemented in our work centers during the first wave of the pandemic, we used different uninfected volunteer samples collected during the same period for cytokine and antibody measurements in contrast to the samples collected for the analysis of genotoxicity and cytotoxicity. Since we did not have access to some clinical information such as symptomatology and the temporality of disease onset, we estimate that the samples were collected between the 4th and 14th day after viral infection. Accordingly, it has been shown that PCR tests are positive after at least 4 days of SARS-CoV-2 infection and the IgG-specific antibodies are evident at around 1 week after symptom onset (Puhach et al., 2023; Sun et al., 2020b; Younes et al., 2020).
Recent diagnostic strategies based on the detection of genes associated with severity, show high sensitivity to discriminate between mild and severe case of COVID-19 (Gravrand et al., 2023). Nevertheless, the current classification of COVID-19 severity (CDC, 2023) has proven to be accurate. The use of different diagnostic tools could strengthen the current triage of patients with COVID-19.
Conclusions
Larger cohorts are needed to associate genotoxicity, cytotoxicity, and the parameters measured. Nevertheless, our results showed interesting evidence of a higher cytokine and an effective IgM and IgG anti-SARS-CoV-2 antibody production in mild cases suggesting an effective immune response against SARS-CoV-2 infection and recovery. Our findings on the presence of cytotoxicity during SARS-CoV-2 infection in mild and severe patients suggests that the virus induces cell damage, which could be triggering the early inflammatory response that in the case of mild COVID-19 is properly regulated. The data presented highlight the relevance of studying the role of genotoxicity and cytotoxicity in the pathophysiology of COVID-19.
Footnotes
Acknowledgment
This work was supported by UNAM Postdoctoral Program (POSDOC).
Authors' Contributions
Conceptualization: G.A., M.R.V., T.P.T., A.M.S., P.M., A.F., P.O.W., and F.M.; data curation: M.A.F.R.; formal analysis: M.A.F.R., T.P.T., and F.M.; funding acquisition: G.A., A.F., P.O.W., and F.M.; investigation: M.A.F.R., G.A., A.M.S., M.S., M.C.V., C.J.C.A., M.C.R., and C.S.O.; project administration: G.A., M.R.V., A.M.S., J.P.R.H., P.M., A.F., P.O.W., and F.M.; resources: M.R.V., J.P.R.H., P.M., A.F., and P.O.W.; supervision: M.R.V., A.M.S., P.M., A.F., P.O.W., and F.M.; writing––original draft: M.A.F.R., T.P.T., and F.M.; writing––review and editing: M.A.F.R., G.A., M.R.V., T.P.T., A.M.S., M.S., M.C.V., C.J.C.A., M.C.R., C.S.O., J.P.R.H., P.M., A.F., P.O.W., and F.M.; all authors have read and agreed to the published version of the article.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This study was financed by grant IN216121 * from PAPIIT-UNAM, Programa Institucional de Salud y Ambiente of the Instituto de Investigaciones Biomédicas (UNAM), and by the Facultad de Medicina, Universidad Nacional Autónoma de México (UNAM), México.
Supplementary Material
Supplementary Table S1
Supplementary Table S2
Supplementary Table S3
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.