
Abstract
Studies assessing the gut mucosal immune balance in HIV-infected patients using intestinal samples are scarce. In this study, we used intestinal mucosal specimens from the ileocecal region of seven immunological nonresponders (INRs), nine immunological responders (IRs), and six HIV-negative controls. We investigated T helper 17 (Th17) and T regulatory (Treg) cell counts and their ratio, zonula occludens-1 (ZO-1), intestinal fatty acid-binding protein (I-FABP), tumor necrosis factor-α, CD4+ T cell counts, HIV DNA, and cell-associated HIV RNA. The results showed that INRs had lower Th17 and higher Treg cell counts than IR, resulting in a significant difference in the Th17/Treg ratio between IRs and INRs. In addition, INRs had lower ZO-1 and higher I-FABP levels than IRs. The Th17/Treg ratio was positively associated with ZO-1 and negatively associated with I-FABP levels. There was a positive correlation between Th17/Treg ratio and CD4+ T cell counts and a negative correlation between the Th17/Treg ratio and HIV DNA in the intestine. Our study suggests that the imbalance of Th17/Treg in the intestine is a characteristic of incomplete immune reconstitution to antiretroviral therapy and is associated with intestinal damage.
Introduction
Combination antiretroviral therapy (ART) efficiently controls HIV replication, slows disease progression, and reduces AIDS-related morbidity and mortality in people with HIV (PWH). However, ∼10–40% of PWH do not achieve CD4+ T cell count normalization despite persistent viral suppression by ART. These patients are known as immunological nonresponders (INRs) and show severe immunological dysfunction (Yang et al., 2020). AIDS and non-AIDS events were significantly increased in INRs compared with those immunological responders (IRs) who achieved both viral suppression and CD4+ T cell reconstitution (Baker et al., 2008; Engsig et al., 2014; Opportunistic Infections Project Team of the Collaboration of Observational HIV Epidemiological Research in Europe (COHERE) in EuroCoord et al., 2012; Pacheco et al., 2015; Takuva et al., 2014; Yang et al., 2020). However, up to date, the mechanisms underlying INRs have not been fully elucidated.
The gastrointestinal (GI) tract, which is composed of an epithelial layer, mucus layer, and underlying lamina propria, serves as a critical structural and immunological barrier that maintains intestinal homeostasis. The GI tract is the primary site of HIV infection and replication. HIV replication in the GI tract results in intestinal damage and immune dysfunction and increases GI permeability and microbial translocation (Alzahrani et al., 2019; Godfrey et al., 2019). The combination of structural damage to the GI tract and mucosal immunodeficiency accelerates the progression of HIV. INRs display more serious intestinal epithelial cell damage and tight junction protein alterations than IRs (Meyer-Myklestad et al., 2022; Somsouk et al., 2015; Tincati et al., 2016). Moreover, INRs have lower CD4+ T cell counts in the GI tract than IRs. However, studies on the association between intestinal immune imbalance and immune reconstitution using intestinal specimens are lacking.
T helper 17 (Th17) cells are defined by their ability to secrete interleukin 17 (IL-17). Th17 are abundant in gut-associated lymphoid tissues (GALT) and play important roles in protection against bacterial and fungal infections, as well as in maintaining mucosal barrier integrity (Higgins et al., 2006; Kanwar et al., 2010; Kleinschek et al., 2006; Mangan et al., 2006; Ye et al., 2001). HIV infection preferentially targets Th17, and depletion in GALT is associated with disruption of mucosal integrity and increased microbial translocation (Brenchley and Douek, 2008; Brenchley et al., 2006; Valverde-Villegas et al., 2015).
In addition, the response of Th17 cells to HIV infection in GALT induces an inflammatory environment. T regulatory (Tregs) and Th17 cells are both derived from naive CD4+ T cells, having complementary and interconnected functions. Tregs primarily perform suppressive functions by inhibiting immune activation and inflammation. The intestinal balance between Th17 and Treg is a state of equilibrium in the intestinal immune system that plays an important role in protection against pathogens, preserving intestinal integrity, and maintaining homeostasis.
The Th17/Treg ratio is typically used as an index of intestinal mucosal immunity. Studies have suggested that maintaining the intestinal Th17/Treg ratio during the therapeutic intervention may be an important strategy for successful immune-based HIV cure (Kanwar et al., 2010; Valverde-Villegas et al., 2015). Most studies have demonstrated that Th17 cell depletion in GALT is associated with disease progression and cannot be fully restored, even after long-term ART (Brenchley et al., 2008; Prendergast et al., 2010). Moreover, an inverse correlation between the counts of Th17 cells and Tregs has been reported during HIV infection, whereby better clinical status is associated with higher Th17 and lower Treg counts in the blood.
The Th17/Treg ratio in the blood is significantly lower in most PWH than in uninfected subjects, even after continued control of HIV replication under ART (Kök et al., 2015; Loiseau et al., 2016). To date, there is a lack of studies performed on intestinal specimens designed to understand the relationship between the Th17/Treg ratio in the intestine and immune recovery after long-term ART. An increased understanding of the immune mechanisms underlying the unfavorable prognosis of INR could identify potential therapeutic targets for ART and improve the health outcomes in PWH.
Given the relationship between HIV disease and the Th17/Treg balance in the blood, we hypothesized that intestinal Th17/Treg ratio depletion might be more serious in INRs than IRs on long-term ART, and this might be associated with damage to intestinal integrity and immune recovery. To address this hypothesis, we collected intestinal biopsies from the ileocecal region of the INRs, IRs, and healthy controls (HCs). Furthermore, we assessed the relationship between the intestinal Th17/Treg ratio and intestinal damage, virus levels, and immune recovery.
Materials and Methods
Participating subjects
Twenty-two participants, including 6 HCs and 16 HIV patients who were treated with ART for more than 2 years with undetectable plasma HIV RNA were enrolled in this study. According to their peripheral CD4+ T cell counts, these HIV patients were classified as IRs (n = 9) if their CD4+ T cell counts were above 500 cells/μL, and as INRs (n = 7) if their CD4+ T cell counts were below 200 cells/μL. Intestinal tissues were collected from volunteer participants using flexible endoscopy. Samples were collected from the ileocecal region. Blood samples were also collected from these participants. Participant characteristics extracted from the clinical data are shown in Table 1. All participants signed written informed consents.
Clinical Characteristics of Enrolled Participants in This Study
All indicators, except gender, are shown as median (interquartile range).
ART, antiretroviral therapy; HCs, healthy controls; INRs, immune nonresponders; IRs, immune responders; NA, not acquired.
Immunofluorescent staining
Sections of paraffin blocks were placed on slides, baked for 1 h at 65°C, then treated in xylene for 10 min, anhydrous ethanol for 2 min, 95% ethanol for 1 min, 80% ethanol for 1 min, and 75% ethanol for 1 min. The antigen repair process was then performed. The sections were immersed in 3% hydrogen peroxide for 20 min to remove devitalized endogenous peroxidase. Then, the sections were blocked with 5% bovine serum albumin for 0.5 h. Next, the samples were stained with antibodies against IL-17 (ab189377, 4K5F6; Abcam, UK), Foxp3 (ab215206, EPR22102-37; Abcam), CD8 (ZA-0508, SP16; ZSGB-BIO, China), and CD4 (ZM-0418, UMAB64; ZSGB-BIO) at 4°C overnight, to determine the presence of Th17 cells, Tregs, CD4+, and CD8+ T cells; nuclei were stained with 4,6-diamidino-2-phenylindole (ZLI-9557; ZSGB-BIO).
Upon completion of staining, the sections were immediately examined under a fluorescence microscope (Leica, Germany). To evaluate the number of Th17 cells, Tregs, CD4+, and CD8+ T cells in the intestine, those cells were counted in five randomly selected visual fields of the same magnification per section to calculate the average count–number of Th17 cells, Tregs, CD4+, and CD8+ T cells in HCs, IRs, and INRs.
Immunohistochemistry staining
Formalin-fixed paraffin tissue sections were baked at 65°C, dewaxed in xylene, antigen repaired, blocked for endogenous peroxidase activity, and incubated with primary antibodies against zonula occludens-1 (ZO-1) (ab221547, EPR19945-296; Abcam), intestinal fatty acid-binding protein (I-FABP) (ab60272; Abcam), or tumor necrosis factor-α (TNF-α) (ab270264, TNF/1500R; Abcam), separately, overnight at 4°C; and subsequently treated with anti-rabbit–horseradish peroxidase (ZB-2301; ZSGB-BIO) as a secondary antibody at room temperature for 2 h. The 3,3′-Diaminobenzidine (ISH-7001; ZSGB-BIO) was used for staining for another 15 min. Finally, the sections were counterstained with Hematoxylin (BSBA-4025; ZSGB-BIO) for cell nuclei detection. Representative images were obtained using a confocal microscope (Olympus, Japan). Immunohistochemistry (IHC) score was determined to assess the staining degree of ZO-1 and I-FABP using the area and intensity scores. The area score was calculated according to the area of staining: <30% = 1 point, 30–60% = 2 points, and >60% = 3 points.
The intensity score was judged by the color depth: no staining or brownish yellow = 1 point, brown = 2 points, and dark brown = 3 points. The ZO-1 and I-FABP score were calculated by multiplying the area score by the intensity score in five visual fields at the same magnification, which was taken randomly on each slide. These methods were described in the literatures (Cui et al., 2020; Li et al., 2017; Lu et al., 2020).
RNAscope in situ hybridization
The RNAscope assay was performed using HIV-1 clade B antisense probes targeting the HIV gag-pol gene (317691; ACD, USA). After baking, dewaxing, antigen repair, endogenous peroxidase activity blocking, and protease digestion, intestinal sections were incubated with the probe for 2 h at 40°C. The amplifiers and detection solution from the RNAscope Multiplex Fluorescence Kit (323100; ACD) were added sequentially for hybridization signal amplification. The images were acquired using an Aperio VERSA scanning system (Leica).
Enzyme-linked immunosorbent assay
Enzyme-linked immunosorbent assay (ELISA) was performed using the Human ZO-1 ELISA Kit (HUES02538; Genie, UK), a Human I-FABP ELISA Kit (RPES2815; Genie), and a Human TNF-α ELISA Kit (MM-0122H1; YGYR-Biotech, China). The 96-well plates, respectively coated with ZO-1, I-FABP, and TNF-α antibodies were incubated with samples and standards at 37°C for 2 h. Then, horseradish peroxidase-conjugated antibodies were added to the wells and incubated for 1 h. Tetramethylbenzidine (TMB) substrate solution, TMB chromogen solution, and acidic stop solution were sequentially added. An EPOCH plate reader (BioTek, USA) was used to measure the optical density of the wells at a wavelength of 450 nm. Standard curves were constructed using the standard absorbance and concentration values.
Quantification of HIV DNA and cell-associated HIV RNA
The HiPure Total RNA Plus Mini Kit (R4121-02; Magen, China) was used to extract RNA from the peripheral blood mononuclear cells (PBMCs) and the intestinal tissues. Total DNA was extracted from PBMCs using the QIAsymphony DNA Mini Kit (931236; Qiagen, Germany) and from intestinal tissues using the Qiagen DNeasy Blood & Tissue Kit (69506; Qiagen). A real-time fluorescence-based HIV DNA/RNA Quantitative Detection Kit (SUPI-1116/SUPI-0103; SUPBIO, China) was used to quantify HIV DNA and cell-associated (CA) HIV RNA. The detection range was from 10 copies/106 cells to 5 × 106 copies/106 cells.
Quantification of ZO-1 and TNF-α by polymerase chain reaction
First, a tissue total RNA Extraction Kit (R4121-03; Magen) was used to extract RNA from the intestinal tissues. As a second step, total RNA samples were quantified using spectrophotometry at 260 nm in accordance with the manufacturer's instructions. Two hundred sixty per 280 nm absorbance ratios ranged from 1.8 to 2.0. An extract of 1 g total RNA was reverse transcribed into complementary DNA (cDNA), and then the cDNA was amplified in the polymerase chain reaction (PCR) solution. The SLAN96P PCR Detection System (Hongshi, China) was used to perform quantitative real-time PCR calibrated with FAM Carboxyfluorescein and VIC Phosphoramidite fluorescent dyes to quantify intestinal ZO-1 and TNF-α. The primer sequences used for real-time PCR were as follows: ZO-1, 5′-CGG ATG GTG CTA CAA GTG-3′ (Forward) and 5′-CAG GGG AGT CTA TTC TAT G-3′ (Reverse); TNF-α, 5′-ACC TCT CTC TAA TCA GCC CTC T-3′ (Forward) and 5′-GGG TTT GCT ACA ACA TGG GCT A-3′(Reverse).
Statistical analyses
Statistical analyses were performed using GraphPad Prism 8.0 (GraphPad Software, USA). Correlation heatmaps were generated using the Origin 2021 software (OriginLab, USA). The Mann–Whitney U and Kruskal–Wallis nonparametric test were used to evaluate statistical differences between two and multiple groups. Spearman's rank correlation coefficient was used to analyze correlations between variables. Statistical significance was set at p < 0.05.
Results
The Th17/Treg ratios were significantly decreased in INRs
As shown in Figure 1A and B, Th17 and Treg cells were detected in the intestinal tissues of HCs, IRs, and INRs through immunofluorescent staining of IL-17 and Foxp3, respectively. INRs had significantly lower Th17 cell counts compared with IRs and HCs (p < 0.05 and p < 0.01, respectively). No significant differences were noted between HCs and IRs (Fig. 1C). Moreover, INRs had higher Treg cell counts than those compared with IRs, with no significant differences between HCs and IRs, and HCs and INRs (Fig. 1D). Furthermore, the INRs had significantly lower Th17/Treg ratios compared with HCs and IRs, with no significant difference between HCs and IRs (Fig. 1D).
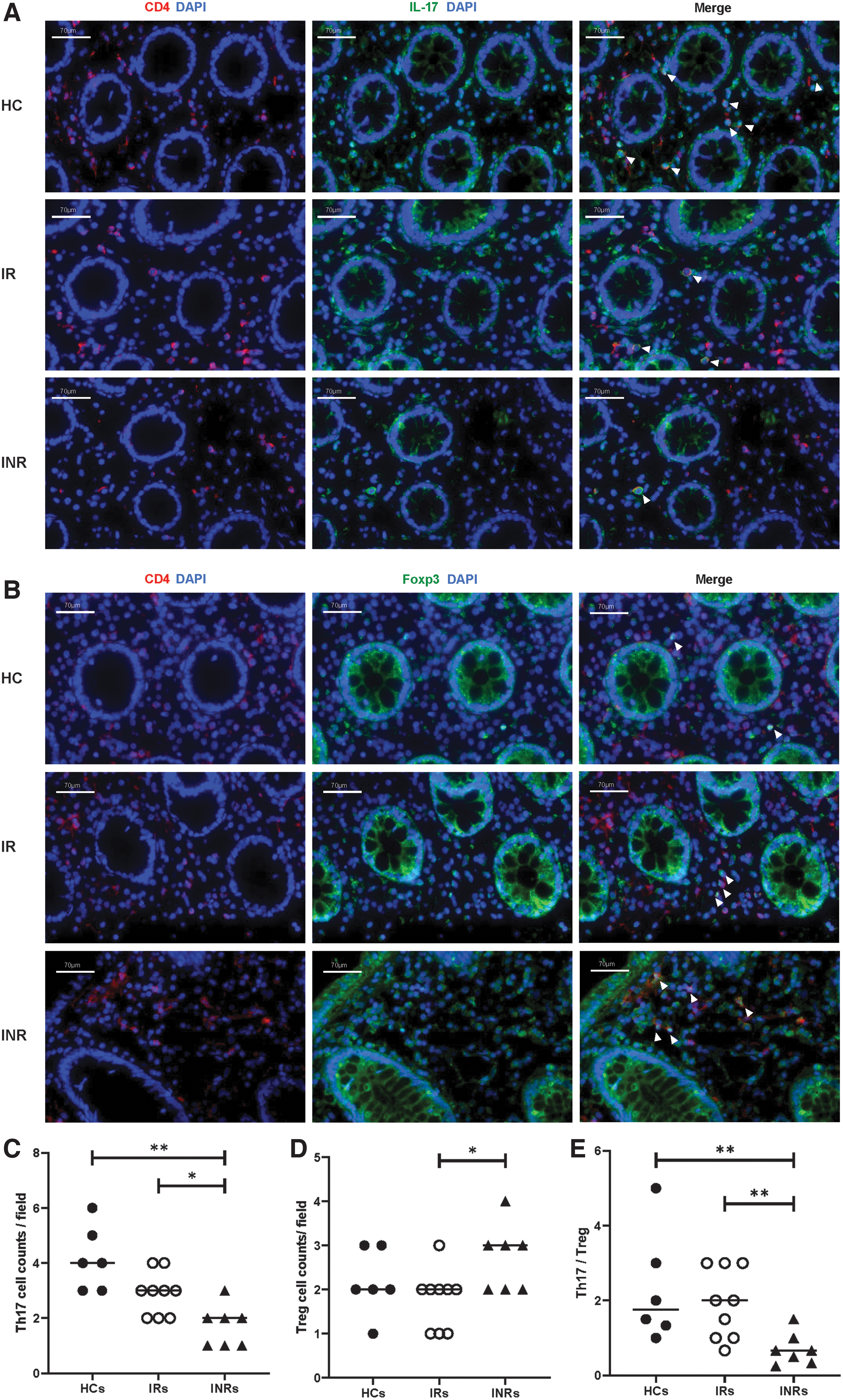
Detection of intestinal Th17 and Treg by immunofluorescent staining
ZO-1 levels positively correlated with intestinal Th17/Treg ratios.
ZO-1 plays an important role in maintaining epithelial integrity and determining intestinal permeability (Kuo et al., 2021). Immunohistochemical analysis revealed the presence of ZO-1 protein in the intestinal tissues of the three study groups: HCs, IRs, and INRs (Fig. 2A). The ZO-1 scores were significantly lower in INRs than in HCs (Fig. 2B, p < 0.05). There was no significant difference between the ZO-1 scores in HCs and IRs, IRs and INRs (Fig. 2B). Moreover, the quantitative polymerase chain reaction (qPCR) analysis revealed the presence of messenger RNA (mRNA) levels of ZO-1 in the intestinal tissues of HCs, IRs, and INRs (Fig. 2C). INRs had lower mRNA levels of ZO-1 than those in HCs (Fig. 2C, p < 0.05) and IRs (Fig. 2C, p < 0.05), with no significant difference between HCs and IRs (Fig. 2C). Moreover, as shown in the correlation heatmap (Fig. 2D), Th17/Treg was positively associated with the ZO-1 score (R = 0.45, p < 0.05) and ZO-1 mRNA (R = 0.60, p < 0.01). Compared with Th17 and Treg cell counts, the correlation between Th17/Treg and ZO-1 levels was the most significant.

Detection of intestinal ZO-1 and exploring the correlations with Th17, Treg, Th17/Treg
I-FABP levels negatively correlated with the intestinal Th17/Treg ratios
I-FABP has been recognized as a marker of intestinal damage (Bykova et al., 2022; Nuzzo et al., 2021; Vollrath et al., 2022). I-FABP was detected in the intestinal tissues of HCs, IRs, and INRs. The I-FABP scores were significantly higher in INRs than in HCs (Fig. 3A, B, p < 0.05), and there was no significant difference between HCs and IRs, and IRs and INRs (Fig. 3A, B). In addition, I-FABP blood levels were higher in INRs than in HCs (Fig. 3C, p < 0.01) and IRs (Fig. 3C, p < 0.01), with no significant difference between IRs and HCs (Fig. 3C). These results indicate more serious intestinal damage in INRs than in HCs or IRs.
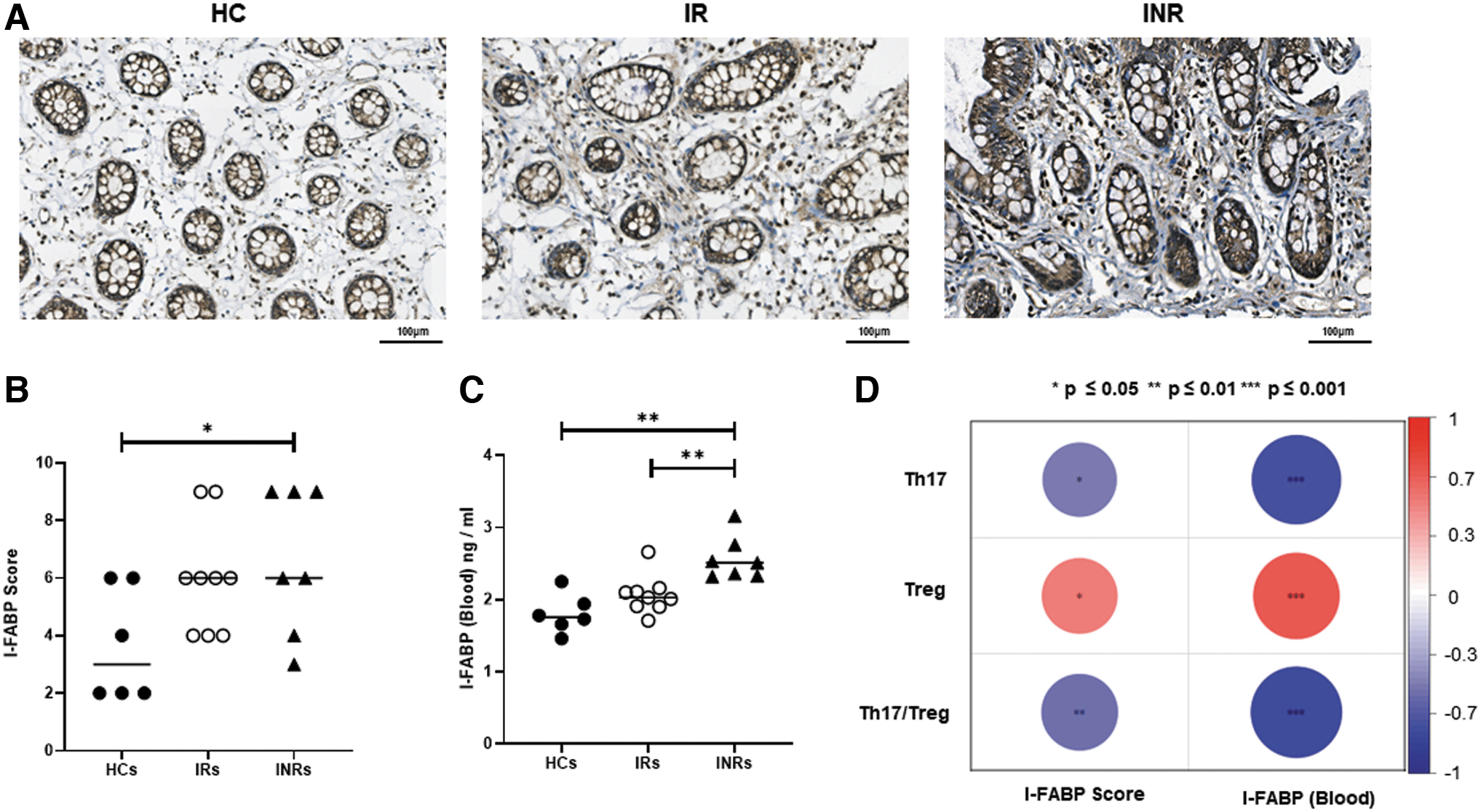
Investigation of the relationships among Th17, Treg, Th17/Treg, I-FABP score, and blood I-FABP
Moreover, the investigation of the correlations between the I-FABP score and intestinal Th17, Treg, and Th17/Treg reveled that the I-FABP score was negatively correlated with Th17/Treg (Fig. 3D, R = −0.55, p < 0.01) and Th17 (Fig. 3D, R = −0.49, p < 0.05). On the other hand, the I-FABP score positively correlated with Treg (Fig. 3D, R = 0.50, p < 0.05). There were significant negative correlations between blood I-FABP and Th17/Treg (Fig. 3D, R = −0.84, p < 0.001) and blood I-FABP and Th17 (Fig. 3D, R = −0.76, p < 0.001), and positive correlations between blood I-FABP and Treg (Fig. 3D, R = 0.69, p < 0.001).
The Th17/Treg ratios positively correlated with intestinal TNF-α levels in IRs and INRs
TNF-α is an important inflammatory cytokine in the intestine. Intestinal TNF-α was assessed using IHC. TNF-α antibodies were used to stain the intestinal sections of the 22 participants (Fig. 4A). Immunohistochemical analysis showed higher expressions of TNF-α in IRs and INRs compared with HCs (Fig. 4A). These findings were confirmed by qPCR (Fig. 4B, p < 0.01, p < 0.05). Moreover, blood TNF-α levels were lower in HCs than those in IRs (Fig. 4C, p < 0.05) and INRs (Fig. 4C, p < 0.05); no significant differences were noted between IRs and INRs (Fig. 4C). In addition, Tregs derived from the intestine negatively correlated with TNF-α (p < 0.01), whereas Th17/Treg positively correlated with TNF-α (p < 0.01) in IRs and INRs. These correlations are illustrated in the heatmap (Fig. 4D).
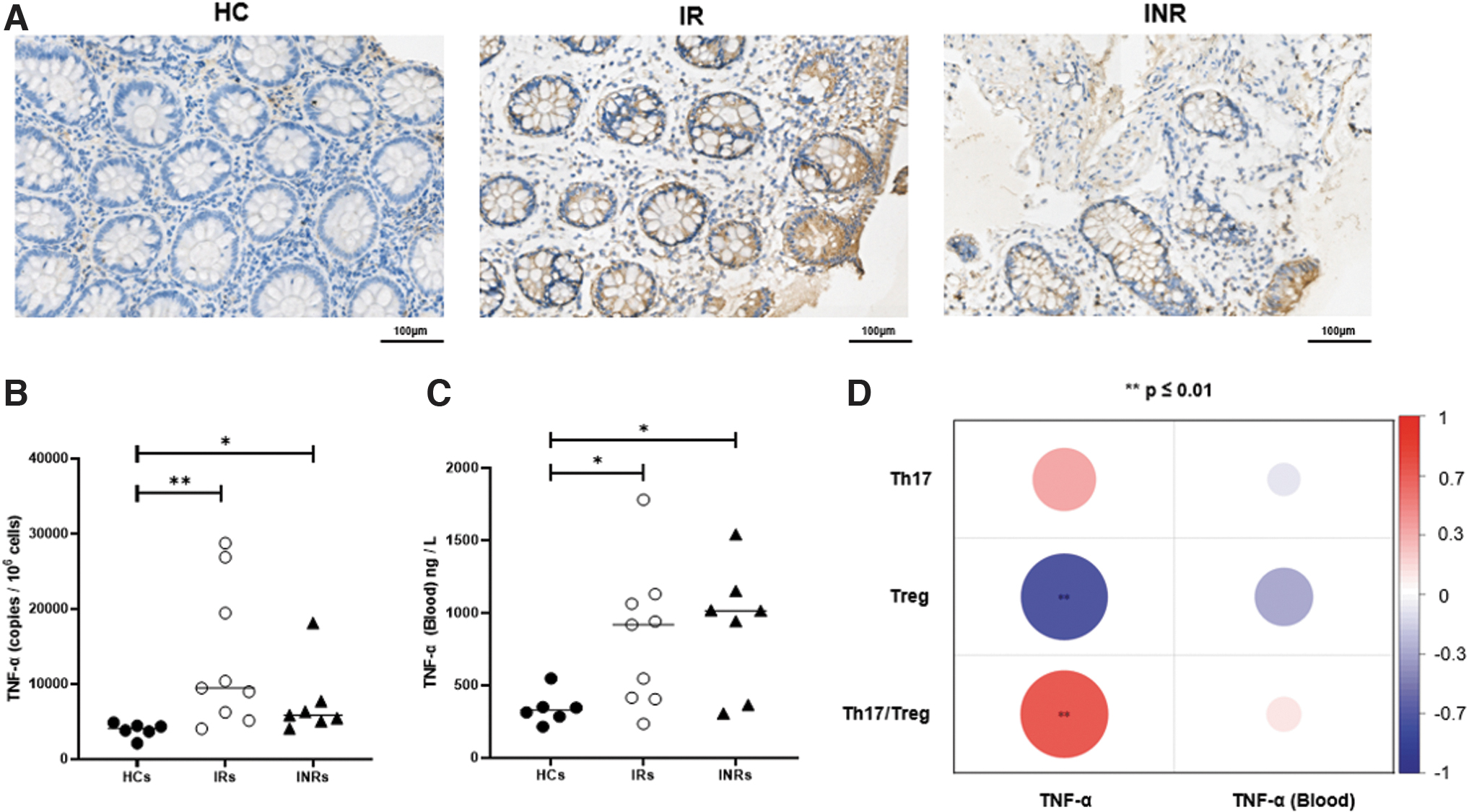
The correlations of TNF-α detected from blood and intestine with Th17, Treg, and Th17/Treg.
HIV DNA levels negatively correlated with the intestinal Th17/Treg ratios
RNAscope in situ hybridization of the intestinal tissues showed positive signals in the IRs and INRs. No positive signals were detected in HCs (Fig. 5A). Moreover, the qPCR analysis showed an increased HIV DNA and CA HIV RNA in INRs compared with IRs (Fig. 5B, p < 0.01 and p < 0.001, respectively). Moreover, increased intestinal HIV DNA and CA HIV RNA levels were detected in the INRs compared with the IRs (Fig. 5C, p < 0.05 and p < 0.001, respectively). Interestingly, intestinal HIV DNA levels were negatively associated with the Th17/Treg ratios (p < 0.01) and Th17 cells (p < 0.05) and positively associated with Tregs (p < 0.01). Tregs were also positively correlated with HIV DNA extracted from PBMCs (p < 0.05), as displayed in the heatmap (Fig. 5D). Thus, HIV DNA was negatively associated with the intestinal Th17/Treg ratio.
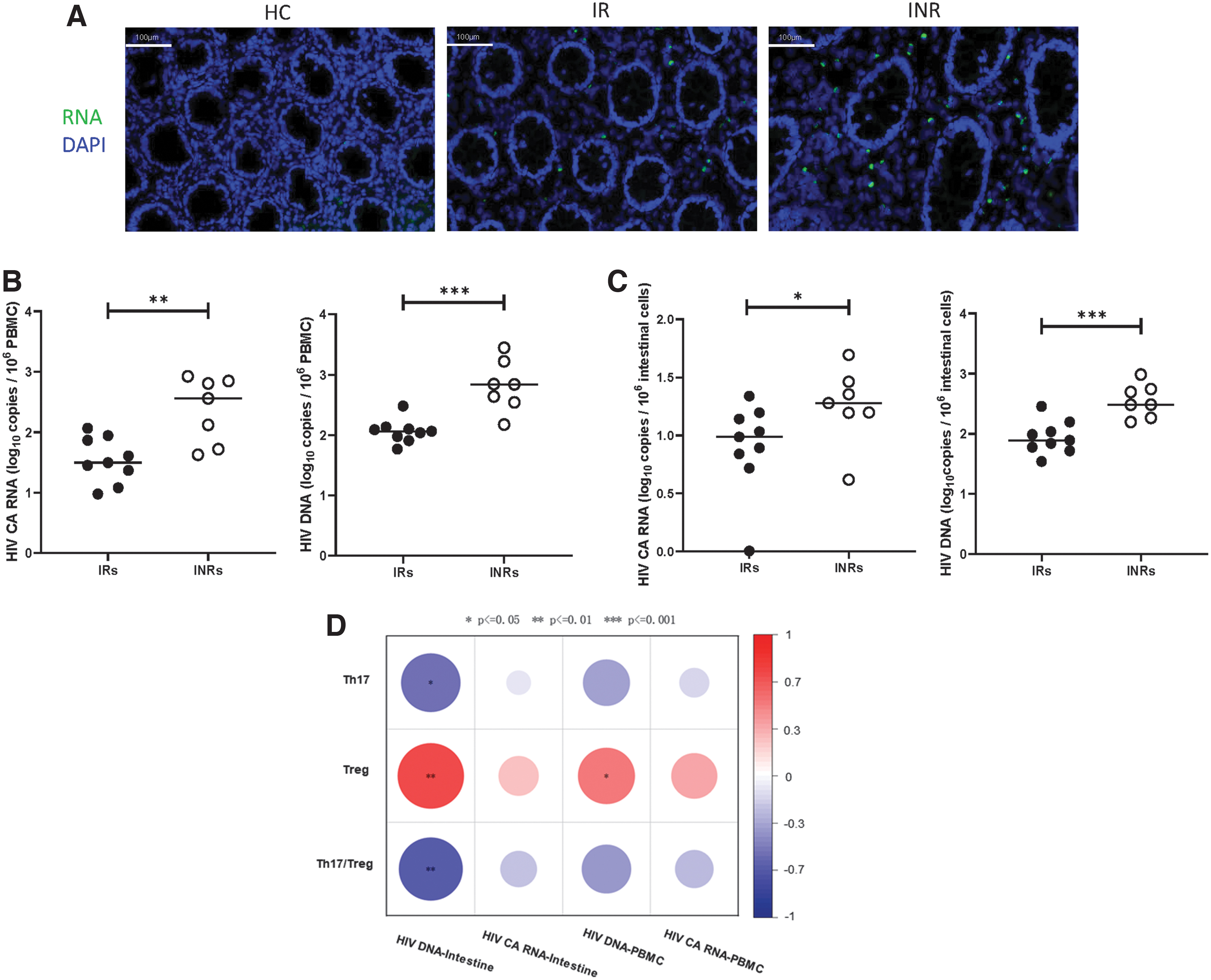
Exploring the correlations of CA HIV RNA and HIV DNA with Th17, Treg, and Th17/Treg in the intestine
Analysis of the correlation between CD4 and CD4/CD8 in blood and intestine with the Th17/Treg ratio
Immunofluorescence staining was used to detect CD4+ T cells and CD4/CD8 in the intestine (Fig. 6A). As displayed in Figure 6B, immunofluorescence staining analysis showed significantly higher CD4+ T cell counts in HCs compared with IRs (Fig. 6B, p < 0.05) and INRs (Fig. 6B, p < 0.01). In addition, there was no statistically significant difference between IRs and INRs in the intestine. In contrast, CD8+ T cell counts in HCs were lower than those in INRs (Fig. 6B, p < 0.05), and no difference was observed between IRs and INRs, and HCs and IRs.
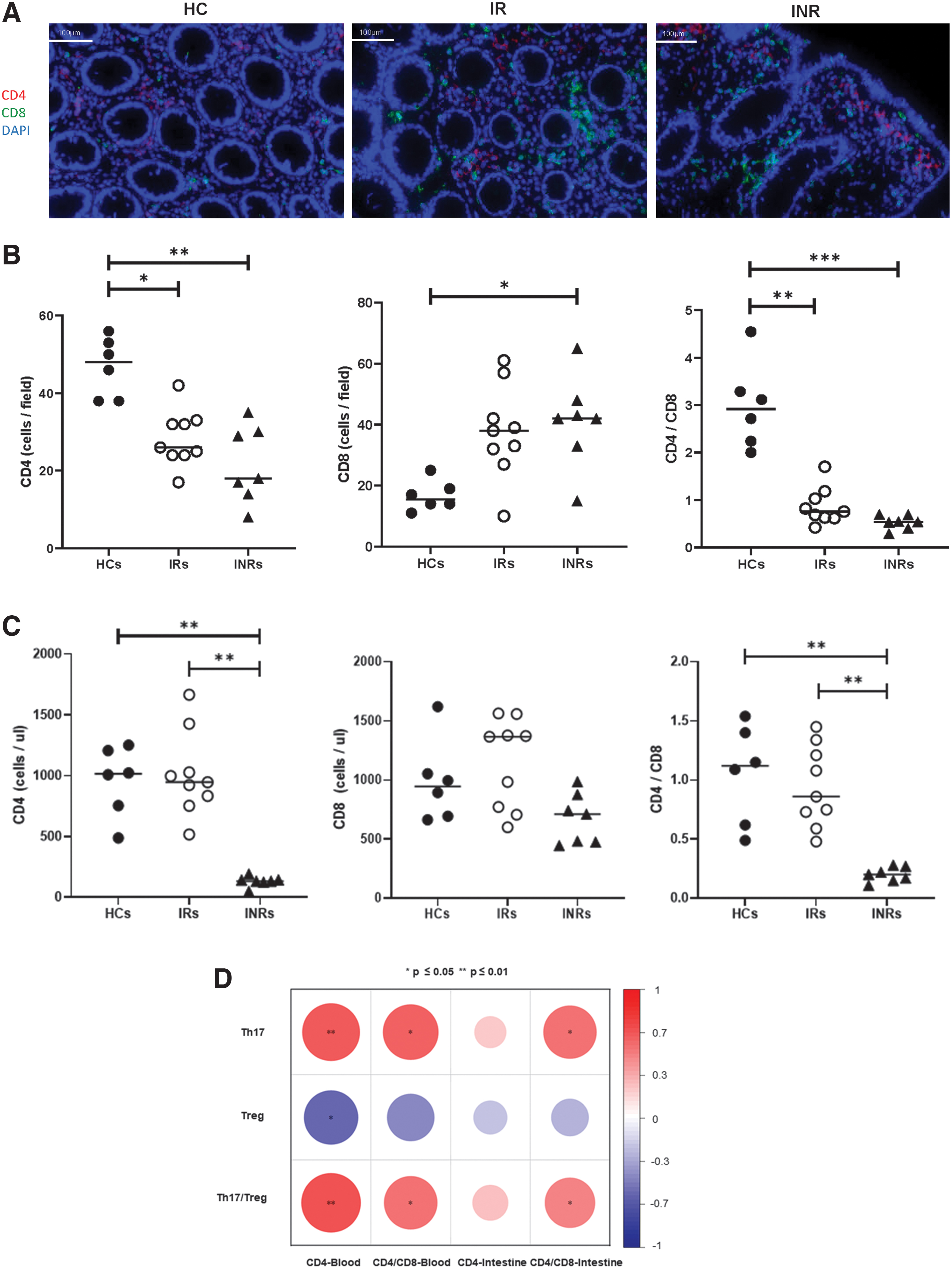
Detection of CD4, CD4/CD8 in peripheral blood and intestine, displaying their correlations with Th17, Treg, and Th17/Treg.
The CD4/CD8 ratios derived from HCs were higher than those derived from IRs (Fig. 6B, p < 0.01) and INRs (Fig. 6B, p < 0.001). Moreover, the analysis of CD4, CD8, and CD4/CD8 levels in peripheral blood showed significantly lower CD4+ T cell counts in INRs compared with IRs (Fig. 6C, p < 0.01) and HCs (Fig. 6C, p < 0.01), with no significant difference between HCs and IRs (Fig. 6C). CD8+ T cell counts have no significant difference between HCs, IRs, and
Finally, CD4+ T cell and CD4/CD8 derived from the intestine and blood were subjected to correlation analyses with Th17/Treg (Fig. 6D). It was clear that CD4+ T cell in the blood was positively correlated with Th17/Treg (Fig. 6D, p < 0.01), Th17 (Fig. 6D, p < 0.01), and negatively correlated with Treg (Fig. 6D, p < 0.05); CD4/CD8 in the blood was positively correlated with Th17/Treg (Fig. 6D, p < 0.05), Th17 (Fig. 6D, p < 0.05), and CD4/CD8 in the intestine was positive for Th17/Treg (Fig. 6D, p < 0.05), and Th17 (Fig. 6D, p < 0.05).
Discussion
In this study, we compared the Th17, Treg, and Th17/Treg ratio in the intestine between HCs, IRs, and INRs and found that the Th17/Treg ratio was the most obvious difference between INRs and IRs. We also analyzed the correlation between Th17, Treg, and the Th17/Treg ratio in the intestine and ZO-1, a marker of intestinal epithelial integrity (Hunt et al., 2014; Mudd and Brenchley, 2016), and blood I-FABP level, a marker of epithelial cell damage (Meyer-Myklestad et al., 2022). The results showed that ZO-1 was most positively associated with the Th17/Treg ratio, and I-FABP was most negatively associated with the Th17/Treg ratio. These results indicate that the balance of Th17/Treg cells is crucial for intestinal epithelial integrity and is related to immune reconstruction.
These results are in line with previous reports that state a percentage of Th17 cannot be restored and that proportions of Tregs are still higher after long-term ART in patients who are HIV-positive compared with HIV-negative persons (Depincé-Berger et al., 2016; Fert et al., 2022; Meyer-Myklestad et al., 2022; Paradowska-Gorycka et al., 2020; Song et al., 2019; Van de Wijer et al., 2021; Wacleche et al., 2017; Yukl et al., 2010). Our results are also consisted of the report that higher blood I-FABP level is the character of INRs and downregulates ZO-1 in gut in INRs (Serrano-Villar et al., 2016).
Many factors may explain the significant decrease in the Th17/Treg ratio in the intestine of INRs. First, the increased number of Treg and decreased number of Th17 in the intestine of INRs could be linked to enhanced indoleamine 2,3-dioxygenase (IDO) expression. Increased IDO activity is associated with HIV disease progression and mortality in people live with HIV (PLWH) (Yang et al., 2020). It has been reported that IDO-1 induces tryptophan (Trp) to kynurenine (Kyn), which can promote CD4+ T cell differentiation toward Treg (Jenabian et al., 2015; Jenabian et al., 2013; Renault et al., 2022) and inhibits the generation of Th17 (De Luca et al., 2007; Romani et al., 2008; Wacleche et al., 2017), leading to a decrease in the Th17/Treg ratio (Baban et al., 2009; Valverde-Villegas et al., 2015). Additionally, IDO-1 is highly expressed by GALT macrophages (Favre et al., 2010; Favre et al., 2009; Smith et al., 2011; Wacleche et al., 2017); studying GALT macrophages and their IDO expression is beneficial for further clarification of the underlying mechanism.
Second, increased frequencies of Treg are associated with intestinal lymphoid collagen deposition, mucosal fibrosis, and architectural disorganization, and GALT CD4+ T cell reconstitution in GALT is negatively correlated with fibrosis in this tissue (Khan et al., 2017; Preza et al., 2015). From Figures 2 and 3, we can see that fibrosis and architectural disorganization are obvious in INRs. Third, the homeostasis of Th17/Treg cells mainly depends on environmental factors, and the gut microbiome may lead to changes in immune responses in GALT during ART. In addition, GI injury, microbial translocation, and non-AIDS-related diseases appear to be interrelated. Therefore, monitoring and maintaining intestinal Th17/Treg balance during ART may be critical for assessing the success of immune reconstitution.
TNF-α plays two important roles in the intestinal–epithelial barrier. TNF-α is an important cytokine in the immunological response against viruses. However, excessive production of TNF-α can have harmful effects on the intestinal–epithelial barrier. The overproduction of TNF-α can induce intestinal epithelial cell apoptosis (Brenchley and Douek, 2008). IRs and INRs displayed significantly higher TNF-α levels than HCs in this study. Although there was no statistically significant difference between IRs and INRs, IRs tended to have higher TNF-α levels. This is consistent with a report that there was no obvious difference in inflammatory cytokines between INRs and IRs (Meyer-Myklestad et al., 2022) TNF-α was positively associated with the Th17/Treg ratio, which may be because high Th17 and low Treg both contribute to TNF-α production. Therefore, INRs showed that intestinal epithelial cell damage and Th17/Treg imbalance were more prominent features than inflammation.
Consistent with previous reports (Zhang et al., 2021), we demonstrated that INRs have higher HIV DNA and CA HIV RNA both in the blood and intestine than IRs. Compared with blood, CD4+ T cell counts and CD4/CD8 ratios showed slower recovery in intestinal biopsies, even in IRs.
In conclusion, our study suggests that the imbalance of Th17/Treg cells in the intestine is a characteristic of incomplete immune reconstitution to ART and is associated with intestinal damage. Additional studies are needed to further explore the mechanism and to investigate whether improving the homeostasis of Th17/Treg cells increases intestinal immune recovery. Considering the small number of participants in this study, a larger cohort is needed to confirm this conclusion.
Footnotes
Authors' Contributions
Y.G., Y.J., F.W., Y.C., and P.M. conceived and designed this study. Y.G., X.G., L.F., and Z.W. collected clinical samples and performed the experiments. M.Q., C.Z., X.F., J.S., W.X., and B.Y. collected clinical information. J.Z., R.X., F.W., Y.C., and P.M. provided the experimental reagents. Y.G., X.G., L.F., Z.W., and M.Q. performed the statistical analysis and figures. Y.G., X.G., L.F., and Y.J. wrote the article. M.Q., C.Z., X.F., J.S., W.X., J.Z., R.X., P.M., Y.C., and F.W. provided comments and supervisions. All authors reviewed and approved the final article.
Ethics Approval
This study was approved by the Research Ethics Committee of the Fifth Medical Center of the PLA General Hospital in accordance with the Declaration of Helsinki (Protocol numbers: ky-2021-7-6-1).
Author Disclosure Statement
The authors declare no conflicts of interest.
Funding Information
This study was supported by the National Key R&D Program of China (2022YFA1303600), the National Natural Science Foundation of China (Nos. 82272317, 82171732), the Chongqing Medical Scientific Research Project (2020FYYX118), the Chongqing Talent Cultivation Program (cstc2021ycjh-bgzxm0275), the Joint Medical Research Projects of Chongqing Municipal Health Committee and Chongqing Municipal Science and Technology Bureau (2022QNXM032), and the Chongqing Science and Technology Bureau (cstc2020jscx-cylhX0001).