
Abstract
Respiratory virus infections are the main causes of pediatric diseases. Human metapneumovirus (hMPV) is an enveloped RNA virus similar to severe acute respiratory syndrome coronavirus type 2, both of which have emerged as important new respiratory viruses. Recent studies have found that interleukin-4 (IL-4) is involved in the replication of a variety of viruses, and its role differs in different viruses. The purpose of this study was to investigate the effect of IL-4 on hMPV and to elucidate its mechanism of action. We found that hMPV infection promoted the expression of IL-4 in human bronchial epithelial cells. The replication of the virus was reduced using small interfering RNA knockdown of IL-4 expression, while the addition of exogenous recombinant human IL-4 to IL-4 knockdown cells restored viral replication ability. These results demonstrate that the expression of IL-4 is closely related to the replication of hMPV; moreover, further experiments revealed that IL-4 promotes the replication of hMPV through a mechanism dependent on the Janus kinase/signal transductor and transcription activator 6 signaling pathway. Therefore, anti-IL-4 strategies may be a promising avenue for the treatment of hMPV infection, representing an important breakthrough for children at risk from hMPV infection.
Introduction
In recent years, respiratory viruses have had a serious impact on the world's public health and economy, especially in the case of the COVID-19 pandemic caused by severe acute respiratory syndrome coronavirus type 2 (SARS-CoV-2). Numerous studies have confirmed the importance of human metapneumovirus (hMPV), often ranking it as the second or third most common cause of severe acute respiratory illnesses in children. Like SARS-CoV-2, hMPV infection causes a variety of symptoms, including cough, wheezing, pneumonia, and bronchiolitis (Panda et al., 2014). hMPV is a member of the Metapneumovirus genus, a pulmonary virus subfamily of Paramyxoviridae, and is a nonsegmented, negative-strand, enveloped RNA virus (Kikuta, 2006).
hMPV can affect people of all age groups, although children, the elderly, and people with underlying cardiopulmonary conditions and immune deficiencies are most susceptible to infection (Papenburg and Boivin, 2010). Owing to a lack of understanding of the pathogenesis of hMPV, there is no effective vaccine or drug to prevent or treat hMPV infection in clinical settings (Van Den Bergh et al., 2022).
Studies have found that interleukin-4 (IL-4) levels increase significantly after hMPV infection in children (Gu et al., 2017; Xiang et al., 2022). IL-4 is the signature cytokine of the type 2 immune response, which can enhance immunoglobulin E-mediated humoral immunity, has the ability to kill cells, and has important immunomodulatory activity. IL-4 also affects a variety of immune cells, such as B cells, airway epithelial cells, and endothelial cells (Iwaszko et al., 2021). Signal transductor and transcription activator 6 (STAT6), a member of the STAT family, is one of the most important downstream targets of IL-4, and transcriptional activation of the STAT family is mainly activated by membrane receptor-related Janus kinases (JAKs) (Fu et al., 2019).
The JAK/STAT6 pathway plays a role in regulating cell growth, differentiation, survival, and pathogen resistance. Protein inactivation, mutation, and overexpression in the JAK/STAT6 pathway can greatly affect the infection success of many viruses, including hepatitis B virus (HBV), respiratory syncytial virus, and human immunodeficiency virus (HIV) (Chen et al., 2017; Quan et al., 2017; Yang et al., 2019).
In recent years, cytokines, especially IL-4, have received considerable attention in the field of viral infection, and many studies have shown that IL-4 is involved in the replication of various viruses (Contoli et al., 2015; Gauthier and Tremblay, 2010; Lin et al., 2003; Qin et al., 2008; Reese et al., 2014). However, whether IL-4 affects hMPV remains unclear. This study aimed to explore whether IL-4 is involved in hMPV replication and elucidate its mechanism of action.
Materials and Methods
Cell culture
Human bronchial epithelial (16HBE) and Vero E6 cell lines were purchased from the China Typical Culture Treasure Center (CCTCC). Cells were grown in Dulbecco's modified Eagle medium (DMEM; Gibco, Grand Island, NY, USA) containing 10% fetal bovine serum (FBS; Gibco) and 1% penicillin-streptomycin (NCM Biotech, Suzhou, China). The cells were routinely cultured at 37°C in a 5% CO2 incubator. Once the cells had grown to a dense monolayer, they were digested with 0.25% pancreatic enzyme (NCM Biotech) and cultured in a ratio of 1:3–1:5.
Culture and preservation of viruses
hMPV strain NL/1/00 was reconstructed by our laboratory through reverse genetic techniques. The virus was amplified in Vero E6 cells using DMEM containing 3% FBS, 1% penicillin-streptomycin, and 0.1% pancreatic enzyme (virus maintenance solution). The virus particles were freeze-thawed at −80℃ and released into the medium, after which the cell fragments were removed using high-speed centrifugation. The supernatant was stored at −80℃.
Real-time quantitative PCR analysis
According to the manufacturers' instructions, an RNA extraction kit (Magen, Guangzhou, China) was used to extract cellular RNA after different treatments, and an RT kit (ABclonal, Wuhan, China) was used for reverse transcription of RNA into cDNA. SYBR Green dye was used to detect the expression of IL-4-related genes, and the 2-ΔΔCt method was used to calculate the relative expression of target genes. Table 1 lists the sequences of primers used in this study.
Primer Sequence Information
IL, interleukin; JAK, Janus kinase; STAT6, signal transductor and transcription activator 6.
The reaction procedure was 95°C for 3 min, 95°C for 5 sec, 63°C for 30 sec, and 39 cycles. The reaction was performed in a CFX96 fluorescent thermal cycler (Bio-Rad, Hercules, CA, USA) and the data were analyzed using the CFX Manager software (Bio-Rad).
Detection of hMPV titer
The plasmid containing the hMPV F gene was diluted 10-fold to detect the copy number of hMPV and to draw a standard curve, which was extracted from a similar number of 16HBE cells using an RNA extraction kit (Magen). An RT kit (ABclonal) was used for reverse transcription into cDNA. The TaqMan probe method was used for absolute quantification of the viruses in each group using the conserved sequence of the hMPV F gene, and the copy number of the viruses was calculated using a standard curve.
Sequences were as follows: probe, 5′-TTGCCAACACACGAACTCCATTCCC-3′; forward primer, 5′-GAGCAATAGCACTCGGTGTTG-3′; reverse primer, 5′-TCACAAAATCTTTCA GCTCTCTCAC-3′. The reaction conditions were 95°C for 30 sec, 95°C for 5 sec, 60°C for 30 sec, and 44 cycles.
Transfection of 16HBE cells with short interfering RNA
Short interfering RNA (siRNA) sequences targeting IL-4 were designed and synthesized by the RIBOBIO Corporation (Guangzhou, China). The sequences were as follows: No. 1 siRNA, 5′-AGAGTACGTTGGAAAACTT-3′; No. 2 siRNA, 5′-AGACGATCATGAGAGAGAA-3′; No. 3 siRNA, 5′-AGTGCGATATCACCTTACA-3′; negative control (NC) siRNA, 5′-GGCUCUAGAAAAGCCUAUGCdTdT-3′ and 3′-dTdTCCGAGAUCUUUUCGGAUACG-5′.
For the transfection test, 2 × 105 16HBE cells per well were inoculated into 24-well plates for culture the day before transfection (18–24 h) and the cell density reached ∼70–80% on the second day. Then, the cells were transfected with 40 pmol siRNA using 1.6 μL Lipofectamine 8000 (Beyotime, Shanghai, China) according to the manufacturer's instructions. After 24 h, the transfected cells were infected with 10 multiplicity of infection hMPV. Cells were collected 24, 48, and 72 h after infection and stored for further analysis.
Western blotting
Cells in each group were collected in six-well plates and radioimmunoprecipitation assay buffer and phenylmethylsulfonyl fluoride were mixed into the lysate according to the proportion of each group (Beyotime). The cells were scraped into the lysate using a disposable cell scraper, and the lysate was transferred to a 1.5 mL Eppendorf tube. Vortex centrifugation was used to fully cleave the proteins. After centrifugation at 4℃ and 12,000 rpm for 30 min, the supernatant was gently aspirated into an Eppendorf tube, and protein concentration was detected using the bicinchoninic acid method. Protein loading buffer and cracking solution were mixed in a ratio of 1:4, boiled at 95°C for 5 min to denature the protein, and frozen at −80℃ for later use.
The protein samples were separated using sodium dodecyl-sulfate polyacrylamide gel electrophoresis and transferred to a polyvinylidene fluoride membrane. The samples were closed at room temperature for 15 min using a rapid sealing solution (Beyotime), incubated with primary antibody solution at 4°C overnight, washed with PBS with Tween-20 three times (Beyotime), incubated with secondary antibody solution at room temperature for 1 h, and then washed three times. Finally, hypersensitive enhanced chemiluminescence solution (4A Biotech, Beijing, China) was used for color development, and the ImageJ software was used to analyze gray values. The following antibodies were used: rabbit anti-IL-4 (1:1,000; Abcam, Cambridge, UK); mouse anti-hMPV N protein (1:1,000; Merck Millipore, USA); rabbit anti-p-JAK2, JAK2, p-STAT6, STAT6, β-actin (1:1,000; Cell Signaling Technology, Danvers, MA, USA); donkey anti-mouse (1:5,000; Abcam); and goat anti-rabbit (1:2,000; Cell Signaling Technology) (Supplementary Data).
Immunofluorescence analysis
16HBE cells were immobilized at room temperature for 15 min with 4% paraformaldehyde and then incubated at room temperature for 1 h with immunostaining blocking solution (Beyotime). The cells were incubated with mouse anti-hMPV N protein (1:500; Merck Millipore) overnight at 4°C. The cells were washed thrice with PBS and incubated with goat anti-mouse IgG H&L Alexa Fluor 488 (1:200; Abcam) at 37°C for 1 h. After washing three times with PBS, an anti-fluorescence quenching sealing solution containing 4,6-diamino-2-phenylindole was added (Beyotime), and fluorescence was observed under a fluorescence microscope.
Statistical analysis
All data were collected from three separate experiments. Graphs were generated using the GraphPad Prism software, version 8.0.2 (San Diego, CA, USA). t-Tests were used for comparison between two groups of data, and one-way analysis of variance tests were used for comparison among multiple groups of data. Data are expressed as the mean ± standard deviation. The statistical significance is indicated as follows: *p < 0.05, **p < 0.01.
Results
hMPV successfully infects 16HBE cells
We first examined the ability of hMPV to replicate in 16HBE cells to assess whether 16HBE cells represent targets associated with hMPV infection. As shown in Figure 1a, the cells were infected and cultured for 24, 48, 72, and 96 h, and the viral load in the cells was quantified at each time point. Quantitative analysis of viral RNA showed that the viral titer increased with an increase in infection time, peaking at 72 h postinfection (hpi), and then decreased. Then, immunofluorescence analysis was performed on cells uninfected with hMPV and those infected with hMPV for 24, 48, 72, and 96 h. The results showed that the expression of hMPV-N protein increased with the increase in infection time, reaching a peak at 72 h after infection, and then decreased (Fig. 1b). These data indicate that hMPV successfully infects 16HBE cells.
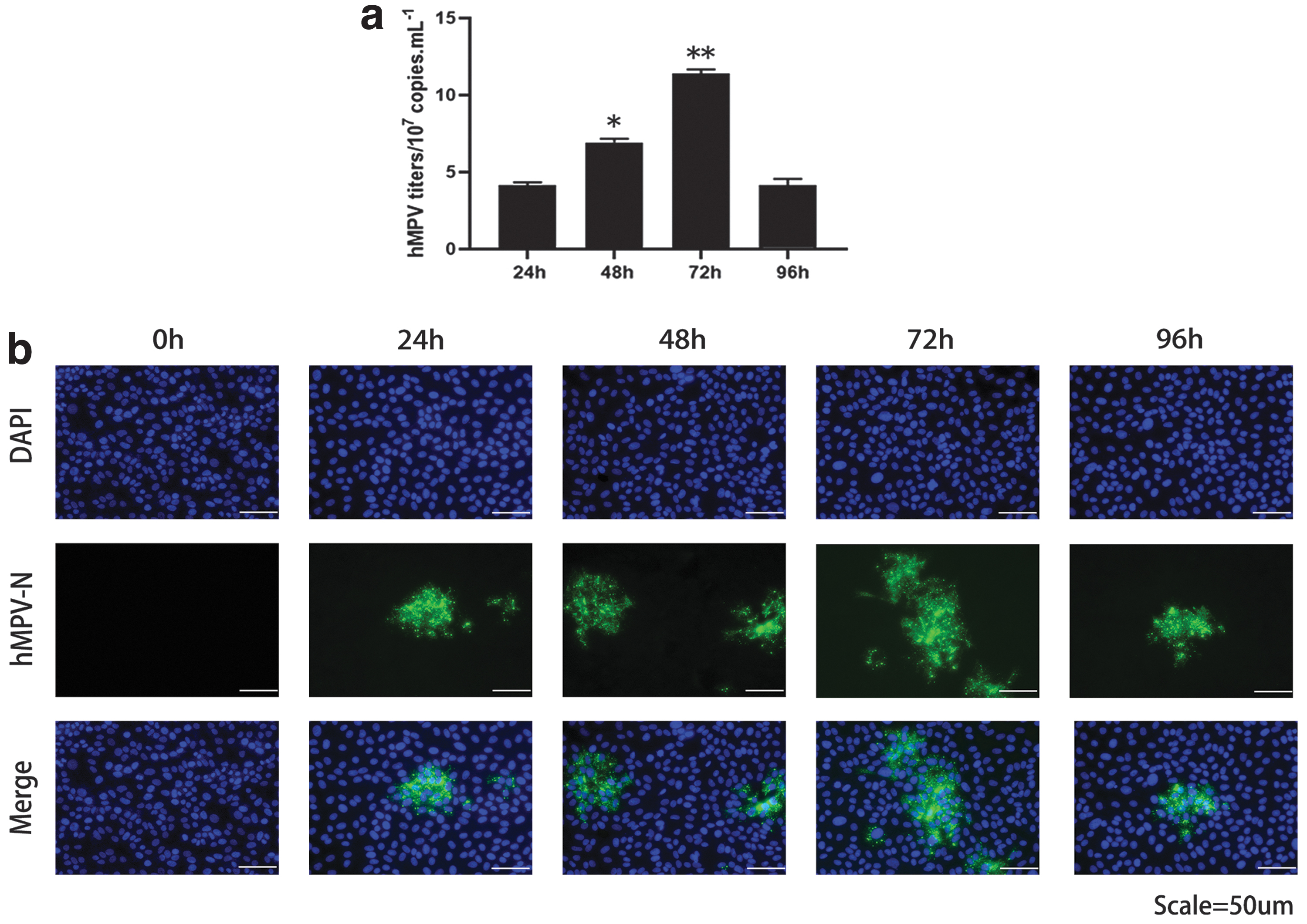
hMPV successfully infected 16HBE cells.
hMPV infection promotes IL-4 expression in 16HBE cells
We analyzed IL-4 mRNA and protein expression in 16HBE cells after 24, 48, and 72 h of hMPV infection. Beginning at 24 h after hMPV infection, IL-4 mRNA and protein expression increased significantly in a time-dependent manner (Fig. 2a–c). IL-4 mRNA and protein expression increased with viral titer in a dose-dependent manner (Fig. 2d–f). These data suggest that hMPV infection promotes IL-4 expression in 16HBE cells.

hMPV infection increased IL-4 expression.
Knockdown of IL-4 reduces hMPV replication in 16HBE cells
To further investigate whether IL-4 was involved in hMPV replication, 16HBE cells were transfected with siRNAs targeting the IL-4 gene (si-IL-4) or negative controls (si-NC). Western blotting results showed that both si-IL-4s reduced the exogenous expression of IL-4 protein (Fig. 3a, b). The results also showed that both si-IL-4s effectively reduced the endogenous expression of IL-4 mRNA (Fig. 3c). si-IL-4 No. 2 was used for all subsequent experiments. Next, we examined the effect of IL-4 knockdown on hMPV replication in 16HBE cells. IL-4 knockdown cells were infected with hMPV, and viral replication was detected at 24, 48, and 72 hpi. First, we detected the expression of hMPV-N protein in cells and found that its expression level was decreased in IL-4 knockdown cells (Fig. 3d, e). After IL-4 knockdown, the virus titer decreased at 24, 48, and 72 hpi, with the most significant decrease observed at 72 hpi (Fig. 3f).
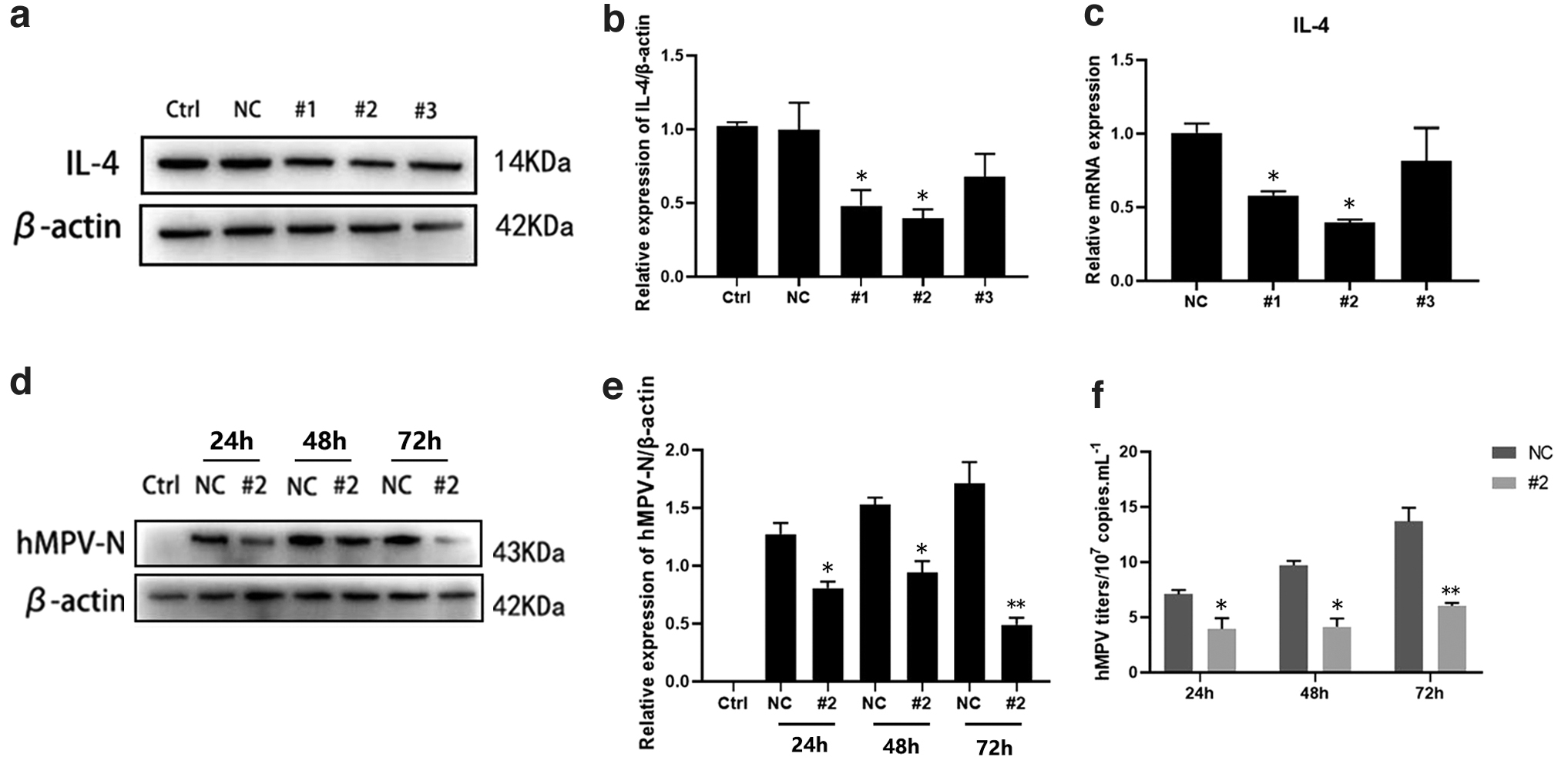
IL-4 silencing inhibited hMPV replication.
We then performed immunofluorescence analysis on hMPV-infected IL-4 knockdown cells, which revealed that the expression of hMPV-N protein was reduced in IL-4 knockdown cells compared with that in the NC group (Fig. 4a). Finally, we stimulated IL-4 knockdown cells with 100 ng/mL recombinant human IL-4 (rhIL-4) for 48 h, infected them with hMPV, and detected viral replication at 24 hpi. As shown in Figure 4b and c, the expression of hMPV-N protein in IL-4 knockdown cells increased after stimulation with IL-4. The results showed that the titer of hMPV in IL-4 knockdown cells was rescued following the exogenous addition of IL-4 (Fig. 4d). These results demonstrate the role of IL-4 in promoting hMPV replication in 16HBE cells.
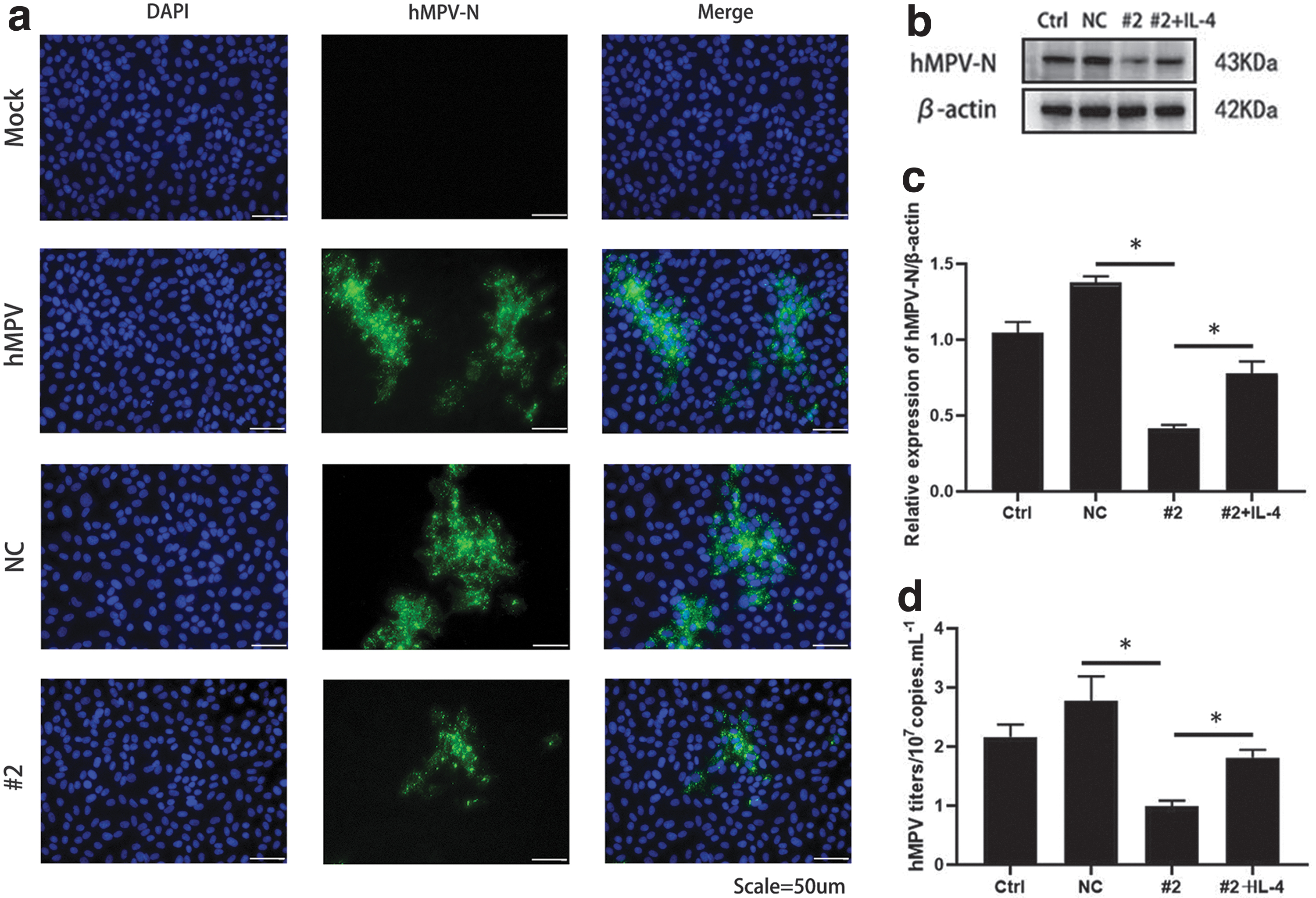
IL-4 promoted hMPV replication.
IL-4 promotes hMPV replication in 16HBE cells
We stimulated 16HBE cells with 100 ng/mL rhIL-4 for 48 h, infected them with hMPV, and then detected viral replication at 24 hpi. As shown in Figure 5a and b, hMPV-N protein expression increased after IL-4 stimulation compared with that in the IL-4 unstimulated (NS) group. Virus titers in cells and collected supernatants were detected, which revealed that virus titers in the IL-4 stimulation group were higher than those in the NS group (Fig. 5c, d). These data suggest that IL-4 promotes hMPV replication in 16HBE cells.
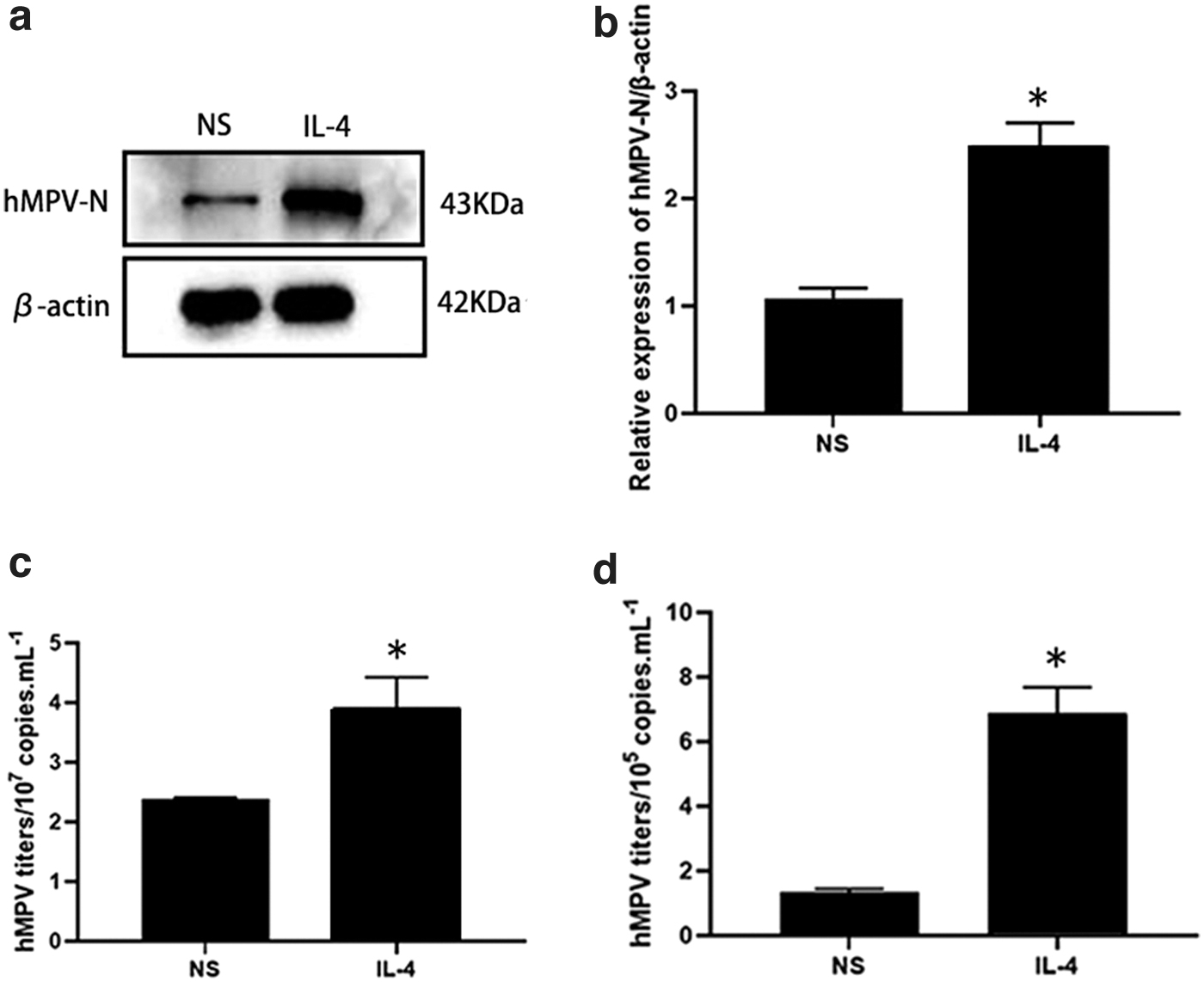
IL-4 enhanced hMPV replication.
JAK/STAT6 pathway is involved in IL-4 expression during hMPV infection
Previous studies have reported that the JAK/STAT6 pathway plays an important role in IL-4 signaling (Athari, 2019; Nakashima et al., 2022; Renu et al., 2020; Tsiogka et al., 2022). In this study, we found that hMPV infection activated the JAK/STAT6 signaling pathway in 16HBE cells (Fig. 6a–c). To determine whether the JAK/STAT6 signaling pathway is involved in hMPV-induced IL-4 expression, we infected IL-4 knockdown cells with hMPV and then detected the changes in p-JAK2 and p-STAT6 at 72 hpi. Reduced levels of JAK2 and STAT6 phosphorylation were observed in IL-4 knockdown cells (Fig. 6d–f). These results suggest that hMPV-induced IL-4 expression is dependent on the JAK/STAT6 signaling pathway (Supplementary Figs. S1 and S2).

JAK/STAT6 signaling pathway involved in hMPV-induced IL-4 expression.
Discussion
Respiratory viral infections are the main cause of morbidity and mortality in neonates, infants, and young children, among whom hMPV is a major contributor (Li et al., 2019). hMPV was discovered as a respiratory virus in the 20th century. Most studies have focused on the epidemiology of this condition, and there is currently no vaccine or specialized treatment for hMPV infection. Multivirus coinfection is clinically common, and hMPV can be cotransmitted with SARS-CoV-2 (Hashemi et al., 2021; Jongbloed et al., 2021; Scott et al., 2021). As the COVID-19 pandemic continues, the widespread outbreak of the virus reminds us of the importance of studying viruses and exploring new antiviral strategies.
hMPV can attack the body through a variety of strategies and cause changes in the infected body that are conducive to viral infection, such as increased cytokine levels. Studies have shown that IL-4 levels in children infected with hMPV are significantly higher than those in healthy children. IL-4 levels are also significantly higher in hMPV-positive patients than in patients diagnosed with influenza virus A or influenza virus B infections (Xiang et al., 2022). In addition, the serum IL-4 level in children infected with hMPV is significantly higher than that in children infected with other viruses (Gu et al., 2017). In this study, we found that the expression of IL-4 mRNA and protein increased with the duration of hMPV infection and was dose-dependent with viral titers.
Regulation of viral replication is critical for disease progression, and IL-4 plays different roles in the replication of different viruses depending on the type of virus, the way the virus infects, and the type and microenvironment of the host cell. Previous studies have reported that IL-4 promotes replication of mouse cytomegalovirus 68 and blocks the antiviral effect of IFN-γ by inducing STAT6 to bind to the promoter of an important viral transcriptional transactivator (Reese et al., 2014). IL-4 production partially promotes the replication of Kaposi sarcoma-associated herpes virus induced by herpes simplex virus type 1 (Qin et al., 2008). IL-4 inhibits the production of interferon in bronchial epithelial cells induced by rhinovirus (RV)-16 and increases RV-16 replication (Contoli et al., 2015).
Studies have also shown that IL-4 can inhibit the expression of the transcription factor C/EBPα in hepatocellular carcinoma Hep3B cells, blocking HBV transcription and reducing HBV replication (Lin et al., 2003). In addition, IL-4 can reduce HIV-1 replication in the human colorectal cell line HT-29 (Gauthier and Tremblay, 2010). In our study, we found that IL-4 knockdown by transfection with siRNAs targeting the IL-4 gene decreased hMPV replication in 16HBE cells. Furthermore, we demonstrated that the exogenous addition of rhIL-4 stimulated 16HBE cells to promote hMPV replication. Therefore, consistent with the aforementioned observations, we established that IL-4 expression is indeed closely associated with hMPV replication, and the underlying mechanisms warrant further study.
Although the efficacy of recombinant IL-4Rα and anti-IL-4 in asthma treatment has been disappointing (Borish et al., 2001; Hart et al., 2002), anti-IL-4Rα, which blocks both type I and type II IL-4 receptors, has yielded promising results in patients with asthma or atopic dermatitis (Beck et al., 2014; Wenzel et al., 2013). In addition, studies have shown that targeting the STAT6 transcription factor can inhibit airway inflammation, eosinophil infiltration, and fibrosis (Oh et al., 2010); moreover, inhibiting STAT6 can reduce tumor growth and spread (Delgado-Ramirez et al., 2020; Lu et al., 2018; Mendoza-Rodríguez et al., 2020). These encouraging results demonstrate the therapeutic potential of targeting the regulation of IL-4 expression and/or signaling. Although the molecular mechanisms that regulate IL-4 expression in pathological type 2 inflammation have been identified, little is known about the regulation of IL-4 in viral infections.
According to the results of this study, hMPV infection can enhance the expression of IL-4, which in turn promotes the replication of hMPV; however, the mechanism has not been well-elucidated. In previous studies, the JAK/STAT6 signaling pathway has been shown to contribute to the regulation of IL-4 expression (Furue, 2020; Wang et al., 2020; Yildiz et al., 2015), while the role of the JAK/STAT6 signaling pathway in inducing IL-4 expression during viral infection has rarely been reported. Our study sheds new light on the role of JAK/STAT6 signaling in this capacity as we discovered that hMPV infection activates the JAK/STAT6 signaling pathway (Fig. 6a–c) and IL-4 knockdown cells infected with hMPV exhibit decreased levels of JAK2 and STAT6 phosphorylation (Fig. 6d–f). These results suggest that the JAK/STAT6 signaling pathway activity is required for IL-4 expression during hMPV infection and provide a putative target for the development of novel treatments.
Taken together, our study demonstrates that IL-4 promotes hMPV replication through a mechanism that is dependent on the JAK/STAT6 signaling pathway. A clear understanding of the effect of IL-4 on hMPV replication may lead to the design of new strategies involving cytokines or their antagonists to remove the virus from the body to better serve susceptible children. As hMPV can alter many cytokine profiles and induce multiple signaling pathways, further research is needed to better understand whether other cytokines and their signaling pathways are also involved in hMPV replication. In addition, we believe that by using the corresponding IL-4 knockout mice to determine the replication capacity of hMPV, our experimental conclusions will be further substantiated. Moreover, obtaining additional clinical data from hMPV-infected patients, particularly with respect to changes in cytokine expression, would contribute to establishing the potential utility of IL-4 as a therapeutic target for hMPV.
Footnotes
Authors' Contributions
Yy.Z., L.N., and Y.Z. conceived and designed the experiments. Yy.Z. performed most experiments and wrote the article. G.W. and Y.Y. assisted with some experiments. Yy.Z. and Y.Z. conducted data analysis and revised the article.
Author Disclosure Statement
The authors declare that there are no conflicts of interest.
Funding Information
This study was supported by the Chongqing Talent Program (CQYC20210303393), the Program for Youth Innovation in Future Medicine at Chongqing Medical University, National Natural Science Foundation of China (81701997), Natural Science Foundation Project of CQ (cstc2019jcyj-msxmX0244).
Supplementary Material
Supplementary Data
Supplementary Figure S1
Supplementary Figure S2
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.