
Abstract
The constant emergence of variants of concern (VOCs) challenges the effectiveness of severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) vaccines over time. This is most concerning in clinically vulnerable groups, such as older adults. This study aimed to determine whether the novel adjuvant MF59-like adjuvant can improve cross-immunity against VOCs in aged animals. We compared the humoral and cellular immune responses of Alum and MF59-like adjuvant-formulated inactivated coronavirus disease 2019 (COVID-19) vaccines against prototype and SARS-CoV-2 variants in 18-month-old mice. Our results showed that two doses of the MF59-like adjuvant inactivated vaccines induced more robust binding and pseudo-neutralizing antibodies (Nabs) against the SARS-CoV-2 prototype and VOCs compared to the Alum-adjuvant and reduced Omicron variant escapes from Nabs in aged mice. The humoral immune responses of inactivated vaccines were much lower against VOCs than the prototype with or without adjuvants; however, T cell responses against VOCs were not affected. In addition, Alum and MF59-like adjuvanted vaccines induced Th1-biased immune responses with increased interferon-gamma and interleukin (IL)-2 secreting cells, and hardly detectable IL-4 and IL-5. Furthermore, the MF59-like adjuvant vaccine produced 1.9–2.0 times higher cross-reactive T cell responses against the SARS-CoV-2 prototype and VOCs than the Alum adjuvant. Therefore, our data have important implications for vaccine adjuvant strategies against SARS-CoV-2 VOCs in older adults.
Introduction
The coronavirus disease 2019
Aging is one of the high-risk factors associated with severe disease, ICU admission, and high mortality caused by COVID-19 (Grasselli et al., 2020; Liwsrisakun et al., 2022). Consequently, older adults are considered the first priority for vaccination. In China, as of December 14, 2022, more than 228 million people over the age of 60 have received COVID-19 vaccines, accounting for 86.6% of this population, most of whom received the Alum adjuvant-inactivated vaccines. However, it is important to note that, owing to the age-related decline of the immune system, or immunosenescence, older adults reported poor immune responses upon receiving the inactivated vaccines (Karamese and Tutuncu, 2022). Moreover, the emergence of VOCs with increased transmissibility and reduced sensitivity to vaccine-elicited antibodies has raised greater concern among older adults.
Based on real-world data, a two-dose schedule of inactivated vaccines only provided limited protection against Omicron infection in older adults during an outbreak in Shanghai, China (Wu et al., 2023). Moreover, the level of vaccine effects on disease progression in older adults was much lower compared with other age groups. Therefore, improving the immune responses of inactivated vaccines against VOCs in the older population is crucial.
Unlike the aluminum hydroxide adjuvant currently used in inactivated SARS-CoV-2 vaccines, MF59 is a squalene-based oil-in-water emulsion widely used in influenza vaccines. It can enhance cross-reactive responses for homologous and heterologous influenza strains in animal models and clinical trials (Lazarus et al., 2016; Zhao et al., 2021a). In this study, 18-month-old mice were used to compare the humoral and cellular immune responses of Alum and MF59-like combined with inactivated COVID-19 vaccine against prototype and VOCs. We aimed to determine whether MF59-like adjuvant can improve cross-immunity against SARS-CoV-2 variants in aged animals.
Materials and Methods
Inactivated SARS-CoV-2 virus
The inactivated SARS-CoV-2 virus used in this study was supplied by Sinovac Life Sciences, Beijing, China. The virus inactivation procedure is the same as CoronaVac, which was described previously (Han et al., 2021). SARS-CoV-2 (CN02 strain) was propagated in African green monkey kidney cells (WHO Vero 10–87 Cells). At the end of the incubation period, the virus was harvested and inactivated with β-propiolactone.
Mouse immunization
This study was conducted in compliance with Beijing CDC's Animal Care and Welfare Committee-approved animal use protocol (approval number: BJCDC IACUC 2022-006, date: March 2, 2022). For immunogenicity studies, we used 18-month-old, specific pathogen-free female BALB/c mice. Mice were purchased from Beijing Vital River Laboratory Animal Co., Ltd. (Beijing, China), housed in approved biosafety Level-2 facilities, and confirmed to be SARS-CoV-2 virus antibody-free using enzyme-linked immunosorbent assay (ELISA) before immunization.
All immunogens used in this study were mixed with an equal volume of AddaVax (an MF59-like adjuvant, InvivoGen, vax-adx-10) or Imject Alum (Thermo Scientific) to a final volume of 100 μL (Table 1). As illustrated in Figure 1, sixty 18-month-old BALB/c mice were randomly divided into six groups (n = 10 per group) and vaccinated using intramuscular route using 100 μL of different immune formulations at day 0 and 14, respectively (Table 1). For each dose, mice were administered 1/10 of the human immunogen dose (0.3 μg/dose/mice). Each animal in the control groups (phosphate buffer saline [PBS] group, PBS+Alum group, PBS+MF59-like group) received PBS, Alum, or MF59-like alone at day 0 and 14, respectively.

Schemes for animal immunization and sample collection. Eighteen-month-old BALB/c mice were randomized into six groups and were immunized at day 0 and 14, respectively. Serum samples were collected on days 14 and 28 for ELISA and pVNTs. Mice were sacrificed at day 28, and splenocytes were isolated for ELISPOT assay. ELISA, enzyme-linked immunosorbent assay; ELISPOT, enzyme-linked immunospot; pVNT, pseudovirus neutralization assay.
Vaccine Formulations and Study Groups
Inactivated SARS-CoV-2 virus group, immunized with Inactivated SARS-CoV-2 virus alone; Inactivated SARS-CoV-2 virus+MF59-like group, immunized with MF59-like adjuvant Inactivated SARS-CoV-2 virus; Inactivated SARS-CoV-2 virus+Alum group, immunized with Alum adjuvant Inactivated SARS-CoV-2 virus; PBS, PBS+Alum, and PBS+MF59-like group, administered PBS, Alum, or MF59-like alone, respectively; w/o, without.
PBS, phosphate buffer saline; SARS-CoV-2, severe acute respiratory syndrome coronavirus 2.
Sample collection
Blood was drawn from all animals at 14- and 28-days postimmunization. After centrifugation at 1,000 g for 30 min, sera were obtained and stored at −80°C before use. Mice from each group were euthanized at day 28, and splenocytes were isolated for enzyme-linked immunospot (ELISPOT) assay.
Enzyme-linked immunosorbent assay
Procedures of the SARS-CoV-2 RBD protein ELISA test have been described previously (Chen et al., 2022; Zhao et al., 2021b). Briefly, 96-well polystyrene microtiter plates were precoated with 2 μg/mL of recombinant SARS-CoV-2 RBD protein (2019-nCoV Wuhan-Hu-1 strain, GenBank Accession No. YP_009724390.1) (Sino Biological, Inc., Beijing, China) diluted in PBS and incubated at 4°C overnight. After blocking and then incubating with serially diluted mouse sera, bound immunoglobulin G (IgG) antibody was detected using horseradish peroxidase-conjugated anti-mouse IgG (1:2,000). Optical density values were measured at 450 nm using an ELISA plate reader (Bio-Rad, Hercules, CA). The endpoint of serum antibody titers was determined as the reciprocal of the highest antibody dilution that was 2.1-fold higher than the optical absorbance value of the PBS (Zhao et al., 2021b).
Pseudovirus neutralization assay
Neutralizing antibodies (Nabs) against Wuhan-Hu-1, Delta (B.1.617.2), and Omicron variant (B.1.1.529) were quantified using the pseudovirus neutralization assay as described previously (Xie et al., 2022).
In brief, pseudoviruses with Wuhan-Hu-1, B.1.617.2, or B.1.1.529 spike proteins were prepared by cotransfecting 293T cells with pCD/NL-BH*DDD, pLenti CMV V5-LUC Blast (w567-1), and pcDNA3.1-Wuhan-HU-1-S, pcDNA3.1-B.1.617.2-S, or pcDNA3.1-B.1.1.529-S. The cell supernatant containing the Wuhan-Hu-1, Delta, and Omicron pseudoviruses was harvested at 48 h after transfection. To determine the neutralization activity of mouse sera, threefold serial dilutions of heat-inactivated serum samples (starting at 1:10) were prepared and mixed with 50 μL of pseudovirus (64,000 TCID50/mL), and the mixtures were incubated at 37°C for 1 h and then incubated with 293T-hACE2 cells (Sino Biological, Inc.) for 48 h. After that, chemiluminescence signals were collected by PerkinElmer EnSight using luciferase substrate (PerkinElmer). Reed–Muench method was used to calculate the half-maximal inhibition dilution (ID50), and ID50 values ≥30.0 were considered positive (Xie et al., 2022).
ELISPOT assay
ELISPOT assays were performed as previously described (Zhao et al., 2022; Zhao et al., 2021b).
The peptide pools consisted of 15 amino acid peptides with an overlap of 11 amino acids spanning the SARS-CoV-2 spike (S)-protein from the Wuhan-Hu-1 or variant strains (GenScript Biotech Corporation, Nanjing, China). The expression of interferon-gamma (IFN-γ), interleukin (IL)-2, IL-4, and IL-5 by splenocytes stimulated with S peptide pools from the SARS-CoV-2 Wuhan-Hu-1 strain was measured using commercial ELISPOT Kits (Cellular Technology Limited, Ohio). The expression of IFN-γ stimulated with S peptide pools from Delta and Omicron variants was also measured using commercial kits following the manufacturer's standard protocol. Lymphocytes from the spleen were harvested from each mouse, and a single cell suspension (2 × 105 cells/well) was seeded in 96-well plates for subsequent analysis. SARS-CoV-2 peptides at 10 μg/mL were used to stimulate S-protein-specific T cell responses. The cell cultures treated with 2 μg/mL phytohemagglutinin served as positive controls (Su et al., 2021). Wells containing unstimulated cells and RPMI 1640 were used as negative controls. The spots were counted using an ELISPOT reader system (Cellular Technology Limited).
Statistical analyses
Statistical analyses were conducted with SAS 9.4 and GraphPad Prism 8.0.1. Specific binding antibodies against SARS-CoV-2 RBD were presented as geometric mean titers (GMTs). Nabs against Wuhan-Hu-1, Delta, and Omicron variants were presented as GMT and seropositivity rate. Titers below the lower limit of detection (LOD) of 1:10 were assigned a value of one half the lower LOD (1:5) for calculation of GMTs. Cellular immune responses were presented as the number of spot-forming cells (SFCs) per 1 million cells. The Wilcoxon matched-pair signed rank test was used to compare the differences between groups. Two-sided p-values of <0.05 were considered significant.
Results
SARS-CoV-2-specific binding antibodies
To examine whether the MF59-like adjuvant can improve the RBD-specific IgG to inactivated SARS-CoV-2 vaccine against prototype SARS-CoV-2 in aged mice, we immunized six groups of 18-month-old mice using different immune formulations twice, at 2-week intervals.
As shown in Figure 2, all vaccine groups produced detectable RBD-specific IgG after the first vaccination. In addition, the MF59-like adjuvanted group had a significantly higher GMT of RBD-specific IgG than the nonadjuvanted group (p = 0.0061) and Alum-adjuvanted group (p = 0.012). After the second dose, RBD-specific IgG titers of all vaccine groups increased dramatically. Among the groups, strong immune responses were evident in the MF59-like adjuvanted group, with 11.3-fold and 3.7-fold higher RBD-specific IgG antibody responses compared to the nonadjuvant and Alum-adjuvanted groups, respectively. These results demonstrate that the MF59-like adjuvant can elicit more-robust RBD-specific IgG antibody responses than Alum or nonadjuvant. Besides, none of the aged mice had any measurable RBD-specific IgG antibody response at baseline, and none in the PBS, PBS+MF59, or PBS+Alum control groups mounted a detectable RBD-specific IgG response during the follow-up.

SARS-CoV-2-specific binding antibodies. Six groups of 18-month-old BALB/c mice were immunized with different immune formulations through intramuscular injection (n = 10). Spike RBD-binding IgG antibodies against prototype SARS-CoV-2 were measured 2 weeks after the first immunization and 2 weeks after the second boost by ELISA. Asterisks indicate statistical significance: *p < 0.05, **p < 0.01. IgG, immunoglobulin G; SARS-CoV-2, severe acute respiratory syndrome coronavirus 2.
Nabs to variants
As shown in Figure 3A, before boosting, only a few immunized mice had detectable neutralizing responses against Wuhan-Hu-1, Delta, or Omicron variant. However, no animal achieved Nab seroconversion (Nabs titer = 30.0) on day 14 across all groups.

Nabs to SARS-CoV-2 prototype and VOCs. Serum samples were collected from each mouse at 14
Two weeks after boosting, the GMTs of all vaccine groups against the Wuhan-Hu-1 and Delta variants increased significantly, but not the Omicron variant (Fig. 3B). Specifically, in the nonadjuvant group, the GMTs were 22.0, 14.1, and 5.5 against the Wuhan-Hu-1, Delta, and Omicron variants, respectively. The seropositivity rate of the nonadjuvant group was 50%, 30%, and 0% against these viruses. In contrast, in the Alum-adjuvant group, the GMTs were 33.7, 22.0, and 12.5, respectively, and the seropositivity rate was 60%, 40%, and 20% against these viruses, respectively. In contrast, in the MF59-like adjuvant group, the GMTs were 39.6, 28.9, and 20.5, with seropositivity rates of 80%, 40%, and 40%, respectively.
These data show that, compared to the nonadjuvant group and Alum-adjuvant group, the MF59-adjuvant group produced 1.8-fold and 1.2-fold higher Nabs against the Wuhan-Hu-1, 2.0-fold and 1.3-fold higher Nabs against Delta variant, and 3.7-fold and 1.6-fold higher Nabs against Omicron variant, respectively. Moreover, these data also demonstrate a 4.0-fold reduction of Nabs against the Omicron variant compared with the Wuhan-Hu-1 in the nonadjuvant group, suggesting that the Omicron variant evaded Nabs in this group. Similarly, a 2.7-fold reduction in Nabs against the Omicron variant was observed compared with the Wuhan-Hu-1 in the Alum-adjuvant group. In contrast, Omicron was 1.9-fold less sensitive to neutralization than Wuhan-Hu-1 in the MF59-like adjuvant group. Overall, these results indicate that MF59-like adjuvant not only elicited a higher Nabs and seropositivity rate against the SARS-CoV-2 prototype and VOCs compared to the nonadjuvant and Alum-adjuvant but also reduced the evasion of Nabs by the Omicron variant in aged mice.
Polarization of T cell responses
Lymphocytes isolated from the spleens of the immunized mice at day 28 were stimulated with S peptide pools from the SARS-CoV-2 Wuhan-Hu-1 strain. IFN-γ, IL-2, IL-4, and IL-5 secreting cells were determined using the ELISPOT assay to assess the magnitude of Th1/Th2 polarization. As shown in Figure 4, the nonadjuvant group exhibited detectable secretion of IFN-γ (SFCs, 62.8) and IL-2 (SFCs, 23.7); however, production of IL-4 (SFCs, 2.2) and IL-5 (SFCs, 3.1) was minimal. Similar trends were observed in the Alum-adjuvant and the MF59-adjuvant groups, with significantly higher secretions of IFN-γ and IL-2 than IL-4 and IL-5. Therefore, these data indicate that, with Alum and MF59-like adjuvants or without adjuvants, inactivated SARS-CoV-2 vaccines induced Th1-biased immune responses in aged mice.
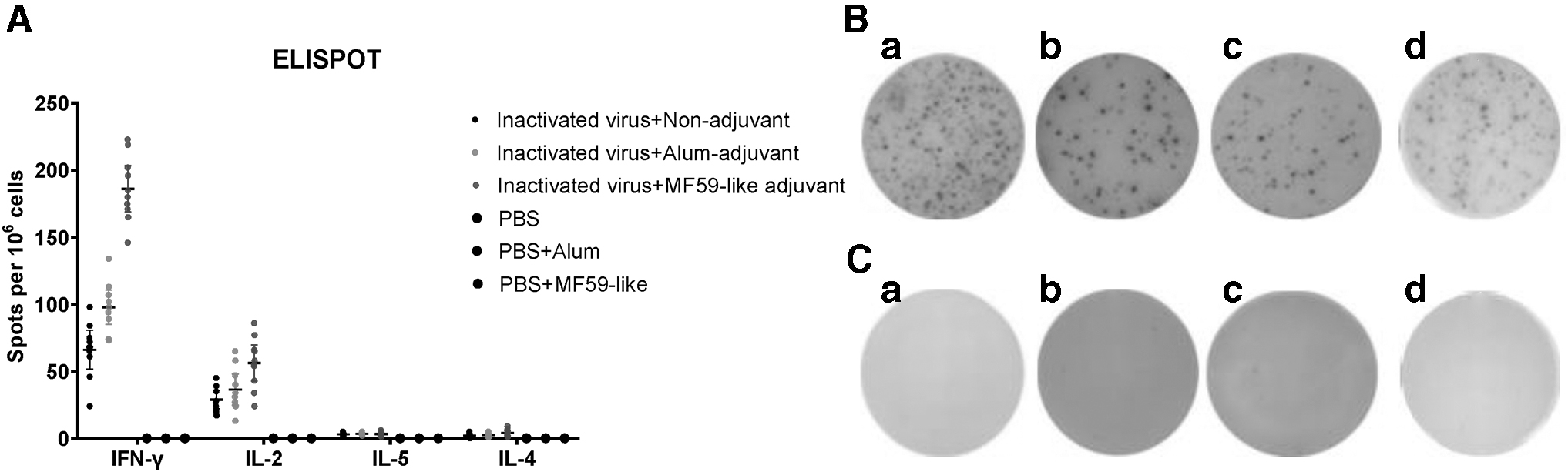
Polarization of T cell responses. Mice were sacrificed at 28 days postinoculation. Mouse splenocytes were stimulated with S peptide pools from the SARS-CoV-2 Wuhan-Hu-1 strain for 24 h, followed by IFN-γ, IL-2, IL-4, and IL-5 ELISPOT assay
Cellular immune responses to variants
Overlapping peptide pools derived from the S peptide pools of the Wuhan-Hu-1, Delta, and Omicron variants were used to stimulate lymphocytes collected from the spleens of the immunized mice at day 28. ELISPOT assays were used to estimate the amount of IFN-γ secreted by the lymphocytes.
As shown in Figure 5, all animals exhibited ELISPOT responses except for PBS, PBS+MF59, or PBS+Alum control groups. In contrast with the differences observed with antibody responses against certain variants, cellular immune responses were comparable against the SARS-CoV-2 prototype and VOCs, with no evidence of decreased responses in any group. However, it is worth noting that the number of IFN-γ-secreting cells against Wuhan-Hu-1, Delta, and Omicron variants in the MF59-like adjuvant group was 184.5, 179.1, and 187.7, respectively. These values were 2.9–3.2 times higher than that in the nonadjuvant group and 1.9–2.0 times higher than that in the Alum-adjuvant group. Therefore, our results showed that the MF59-like adjuvant can induce higher cross-immune IFN-γ responses against the SARS-CoV-2 prototype and VOCs compared to nonadjuvant and alum-adjuvant in aged mice.

T cell responses against the SARS-CoV-2 prototype and variants. Mice were sacrificed at 28 days postinoculation. The splenocytes were isolated and stimulated with S peptide pools from the Wuhan-Hu-1, Delta, and Omicron variants for 24 h, followed by IFN-γ ELISPOT assay. The results are expressed as the number of spot-forming cells per 106 cells. Asterisks indicate statistical significance: *p < 0.05, **p < 0.01, ***p < 0.001, ****p < 0.0001.
Discussion
Many older individuals have been protected from COVID-19 by the implementation of mass vaccination. However, owing to age-related immunosenescence, older individuals develop poor humoral and cellular immune responses to the Alum adjuvant-inactivated COVID-19 vaccine, which raises concern regarding the impact of SARS-CoV-2 variants (Chandrashekar et al., 2021; Yue et al., 2021). In this study, we showed that compared to the Alum-adjuvant inactivated COVID-19 vaccine, the MF59-like adjuvant vaccine induced more robust binding and Nabs against the SARS-CoV-2 prototype and VOCs and reduced the escape of the Omicron variant from Nabs. In addition, the MF59-like adjuvant-inactivated COVID-19 vaccine produced higher cross-reactive T cell responses against the SARS-CoV-2 prototype and VOCs than Alum-adjuvant in aged mice.
Nabs have been identified as a potential indicator of vaccine protection against SARS-CoV-2, as suggested by vaccine efficacy studies, preclinical studies in mice and nonhuman primates, and data from the early use of convalescent plasma in older patients (Khoury et al., 2021; Krammer, 2021). Several research groups, including ours, have shown that commercial COVID-19 vaccines, such as BNT162b2, mRNA-1273, ChAdOx1, and CoronaVac, can induce sufficient Nabs and are effective against the ancestral strain of SARS-CoV-2, thereby demonstrating high efficacy in preventing both asymptomatic and symptomatic infection. However, this situation is currently challenged by the emergence of VOCs (Ling et al., 2021; Zhao et al., 2022). SARS-CoV-2 VOCs, especially the Delta and Omicron variants, have several mutations in the S-protein and can partially evade native and vaccine-induced Nabs (Yu et al., 2021).
Using sera from a cohort aged between 70 and 89 years, who were vaccinated with two doses of BNT162b2, Newman et al. (2022) demonstrated substantially reduced neutralization ability against the Beta, Delta, and Omicron VOCs, among which the neutralizing titers against the Omicron VOC decreased >30-fold. Follow-up studies with other COVID-19 vaccines also revealed a 10-to-25-fold reduction in Nabs against the Omicron VOC after two-dose vaccinations (Zhang et al., 2022). Increasing evidence has shown that the heterologous or homologous booster dose can increase the neutralizing capacity of vaccines against most VOCs; however, the Omicron variant still exhibits obvious evasion from vaccines (Ai et al., 2022). In this study, our results indicate that two doses of the MF59-like adjuvant inactivated SARS-CoV-2 vaccines induced significantly higher Nabs against the SARS-CoV-2 prototype and VOCs than the Alum adjuvant vaccine and reduced Omicron variant escape from Nabs in aged mice. Furthermore, the MF59-like adjuvant may be a better adjuvant than Alum in reducing immune escape against VOCs from the inactivated SARS-CoV-2 vaccine in older adults.
T cells play a critical role in defending against viral infection (Zhao et al., 2022). Therefore, we determined the T cell response to SARS-CoV-2 S-protein by stimulating lymphocytes with overlapping peptide pools to the Wuhan-Hu-1, Delta, and Omicron variants using an IFN-γ ELISPOT assay. Our results showed that MF59-like adjuvant-inactivated COVID-19 vaccine induced significantly higher cross-reactive T cell responses against the SARS-CoV-2 prototype and VOCs compared to the Alum-adjuvant in aged mice. It is increasingly recognized that, although Nabs are predominantly responsible for protection against initial infection, T cells can limit disease progression when Nab titers are low (Moss, 2022). Further evidence from patients infected with COVID-19 has shown that the reduction in the number of T cells is related to poor clinical outcomes and immune pathogenesis. In contrast, adequate T cell counts are associated with rapid viral clearance and patients having mild disease symptoms or successful rehabilitation (Grifoni et al., 2020).
Moreover, because a well-balanced Th1 immune response can lead to virus clearance, immunization with MF59-like adjuvant shows promising capacities of inducing an antiviral response in the host. Therefore, our results indicate the potential effects of the MF59-like adjuvant on disease progression. In addition, many researchers have demonstrated that, compared with the traditional Alum adjuvant, the novel MF59-like adjuvant can induce a higher immune cell response by producing more inflammatory cytokines, activating and maturing more dendritic cells, and increasing antigen presentation (Ko and Kang, 2018). This is crucial because the decline in efficacy of Alum-adjuvant inactivated COVID-19 vaccine in older adults is associated with reduced excitability of cell-mediated and cytotoxic T lymphocyte responses, which are essential for viral clearance.
Interestingly, our results showed that in contrast with the differences observed with antibody responses against certain variants, T cell responses against Omicron were comparable to those against Wuhan-Hu-1 and Delta in any group. This suggests that the inactivated vaccines used in this study elicited highly conserved cellular immunity to SARS-CoV-2 strains. These findings are consistent with reports from human clinical trials (Liu et al., 2022; Riou et al., 2022; Yu et al., 2021).
Unlike humoral immune responses, T cell responses target multiple regions in the S-protein, including the conserved regions which are also present in SARS-CoV-2 VOCs. Therefore, specific mutations in SARS-CoV-2 variants may not lead to the evasion of cellular immunity. In addition to the S-protein, an advantage of inactivated COVID-19 vaccines is that they contain additional conserved SARS-CoV-2 antigens (Samaranayake et al., 2021). As changes in the antigenicity of conserved and internal structural viral proteins, which are the main focus of T cell responses, are rare in VOCs, these inactivated vaccines are less likely to be affected by antibody escape mutations in VOCs (Afkhami et al., 2022). Consequently, the inactivated COVID-19 vaccines are expected to be effective against both ancestral and variant SARS-CoV-2.
However, it is notable that there are some limitations. First, previous research suggests a strong correlation between vaccine immunogenicity and vaccine efficacy (VE), but direct data on VE, particularly in virus challenge studies, are lacking in this study. Second, our study only compared the immunogenicity of Alum and MF59-like adjuvants, and a broader range of adjuvants should be evaluated in clinical trials to identify the optimal choice. Third, we did not assess the long-term durability of humoral and cellular immune responses. These issues are being addressed by the ongoing research programs.
In conclusion, this study demonstrated that the addition of the MF59 adjuvant to the inactivated COVID-19 vaccine reduced the escape of VOCs from Nabs and increased cross-reactive T cell responses against VOCs in aged mice.
Footnotes
Authors' Contributions
Conceptualization, planning, and management: W.Z. and J.W. Laboratory experimental design: W.Z. and J.W. Laboratory execution of experiments, and data acquisition, analysis, and interpretation: S.B., Y.K., W.C., H.X., L.Z., M.L., and J.W. Drafting and revision of the article: S.B. and W.Z.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This study was supported by the Beijing Municipal Science & Technology Commission (Z211100002521014).