
Abstract
Cholesterol 25-hydroxylase (CH25H) and its product 25-hydroxycholesterol (25HC) showed antiviral effects against various viruses in vitro. CH25H expression is regulated in mice by pro-inflammatory cytokine interferons (IFNs) in mice but data on its possible correlation with IFNs in humans are still unclear. We examined gene expression of CH25H, IFN-α, and IFN-β and serum levels of 25HC in Iranian patients with mild and severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2). Fifty intensive care unit (ICU) patients and outpatients with SARS-CoV-2 and 25 healthy controls were studied. Gene expression of CH25H and relevant inflammatory cytokines was quantified in peripheral blood mononuclear cells by real-time polymerase chain reaction. The expression of CH25H and serum levels of 25HC were significantly higher in ICU patients with SARS-CoV-2. Notably, IFN-α levels increased in healthy controls. However, compared to healthy controls, IFN-β was considerably higher in outpatients. Finally, statistical analysis shows that no correlation was found between CH25H and IFN-α expression; nevertheless, a lower correlation was found with IFN-β. The data revealed that CH25H and 25HC levels increase after SARS-CoV-2 infection. In other words, decreased levels of those factors in severe patients compared with mild patients may indicate the importance of their function in controlling the progression of the disease.
Introduction
Since the emergence of severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2), infection with this virus called coronavirus disease 2019 (COVID-19) has caused millions of deaths worldwide (Mousavizadeh et al., 2022). SARS-CoV-2 predominantly affects the respiratory tract. However, studies have also found the virus in cardiac, renal, gastrointestinal, and neurological tissues, indicating the potential to damage various parts of the body (Ram-Mohan et al., 2022). Although, as of 2022, vaccination began to almost cover most of the population for prevention, the requirement for improving effective treatment and different therapeutic strategies for patients with COVID-19 infection seems to be necessary (Asano et al., 2023). Since high levels of pro-inflammatory cytokines have been found in the peripheral blood of patients with severe disease, it has been suggested that severe COVID-19 is related to antiviral immune responses and virus pathogenicity (Fung et al., 2022; Mansourabadi et al., 2023).
The first line of defense against viral pathogens is innate immune system in the body. Inflammatory cytokine interferons (IFNs) are one of the most beneficial components of innate immunity against viral infections. By inducing the expression of various interferon-stimulating genes (ISGs) such as virus inhibitory protein, IFNs-inducible (Viperin), endoplasmic reticulum (ER)-associated, bone marrow stromal antigen 2, and IFN-induced transmembrane proteins, they can trigger antiviral responses (Li et al., 2020). IFN-I family is known as an inhibitor to prevent viral replication. This family consists of two cytokines: IFN-α and IFN-β. IFN-α is produced by hematopoietic cells, while a wide range of cell types produce IFN-β. Notably, it is reported that low level of IFN-α results in increase of autoantibodies against IFN-α, which leads to affect the severity of COVID-19 (Russo et al., 2020). The antiviral activity of IFN-β is shown to be more compared with IFN-α against SARS-CoV2 infection (Radenkovic et al., 2020).
Cholesterol-25-hydroxylase (CH25H), also known as cholesterol-25-monooxygenase, is a member of the redox enzyme family and is composed of 272 amino acids. The Golgi apparatus and the ER are the cellular sites where CH25H catalyzes the formation of its enzymatic product 25-hydroxycholesterol (25HC), which acts as a corepressor of cholesterol biosynthetic enzymes which block sterol regulatory element binding protein processing (Chen et al., 2014; Wei et al., 2022). CH25H is involved in various biological processes such as lipid metabolism, inflammation, and the immune response to viral infections. Studies in mice showed that CH25H is upregulated after stimulation by Toll-like receptor ligands, indicating potential antiviral effects. Studies in mice exhibited that CH25H is an ISG, and 25HC is an antiviral agent that showed antiviral effects against various enveloped and nonenveloped viruses (Xiang et al., 2015).
An important factor closely related to the regulation of CH25H expression is the type I IFN level. Previous studies have indicated that type I IFNs increase expression through the IFN receptor and the JAK/STAT1 pathway upon infection (McDonald and Russell, 2010). In addition, there is a relationship between susceptibility to viral infections and low cholesterol in the serum and exogenous cholesterol such as 25HC which has been used in COVID-19 infections (Zhang et al., 2019).
Based on Kim et al., (2021) nanovesicles such as 25HC@DDAB may be promising candidates to modulate cytokine storm and lung tissue damage in severe COVID-19 patients. While CH25H and its enzymatic product, 25HC, have been suggested as antiviral ISG effectors, there is little knowledge about their antiviral effects in humans as currently published studies are mostly based on cell culture or animal model experiments. The current study was designed and conducted to investigate the role of CH25H and 25HC in patients with COVID-19 and evaluate the association of CH25H and 25HC with disease severity. We also studied whether CH25H is an ISG or not in humans.
Materials and Methods
Study group
From May 2022 to August 2022, 50 patients with clinical symptoms of SARS-CoV-2 were referred to the Infectious Diseases Department of Firouzgar hospital and Keyvan virology laboratory, two SARS-CoV-2 centers in Tehran, Iran, and completed the informed consent form and participated in the study. For the diagnosis of SARS-CoV-2, patients were screened with a real-time polymerase chain reaction (RT-PCR) test using a commercial kit (Pishtaz Teb, Tehran, Iran) in nasopharyngeal swab specimens. In addition, 25 healthy individuals without clinical signs of COVID-19 and with negative SARS-CoV-2 RT-PCR test were included in the study as controls. The study was approved by the Ethics Committee of Iran University of Medical sciences (ethical code: IR.IUMS.FMD.REC.1401.357).
Peripheral blood mononuclear cell isolation and total RNA extraction
In brief, 3 mL of peripheral blood was collected from each patient in an Ethylenediaminetetraacetic Acid (EDTA) tube. Ficoll (DNA Biotech, Korea) density gradient centrifugation technique was used to isolate peripheral blood mononuclear cells (PBMCs). To remove platelets, the pellet was washed thrice with cold phosphate-buffered saline (pH: 7.3 ± 0.1), then centrifuged at 2,000 rpm for 10 min at 4°C, and stored at −80°C until RNA extraction. Total RNA was extracted from PBMCs by the phenol-chloroform method using RNX Plus (Sinaclon, Iran). Seven hundred fifty microliters of RNX Plus was added to the cell lysate and then incubated for 5 min, 200 chloroform was added to pellet, centrifuged at 14,000 rpm for 15 min at 4°C. The supernatant was aspirated and centrifuged at 14,000 rpm for 10 min at 4°C in 500 μL of isopropanol. Afterward, the pellets were centrifuged at 8,000 rpm for 5 min at 4°C with 1 mL of cold 70% ethanol. Finally, after evaporated ethanol, the extracted RNA pellet was dissolved in diethyl pyrocarbonate treated water. RNA concentration was monitored by UV-vis spectroscopy (Thermo Fisher). Extracted RNA samples were kept at −80°C (29).
Complementary DNA synthesis and RNA quantification
Complementary DNAs (cDNAs) were synthesized from total extracted RNA using a cDNA Synthesis Kit (Parstus, Mashhad, Iran) as described by the manufacturer.
The expression patterns of CH25H, IFN-α, and IFN-β were quantified by RT-PCR using a Rotor-Gene Q thermal cycler (Qiagen, Hilden, Germany). Moreover, we investigated the pattern expression of transforming growth factor-beta (TGF-β) among patients with different status. The PCR mix included 0.5 pmol each of primer, 10 μL of SYBR Green PCR Master Mix (Ampliqon; Denmark), 2 μL of cDNA template, and 6 μL of nuclease-free distilled water to obtain a reaction volume of 20 μL. RT-PCR steps were initial denaturation of cDNAs for 15 min at 95°C, followed by 40 cycles of denaturation for 15 sec at 95°C, annealing for 25 sec at 60°C, and extension for 30 sec at 72°C. The β-actin housekeeping gene was used as internal control. In Table 1, the list of primer pairs used for quantification of each gene can be found. Relative expression values were calculated using the 2−ΔΔCt method.
List of Primers Used in this Study
CH25H, cholesterol 25-hydroxylase; IFN, interferon.
25HC assay and cholesterol measurement
The level of 25HC in serum samples was determined using a commercial Enzyme-linked Immunosorbent Assay (ELISA) Kit (ImuGex GmbH, Germany) for human 25HC, 25-OHC according to the manufacturer's protocol. The quantity of cholesterol levels was determined by cholesterol enzymatic-colorimetric CHOD-POD Kit (BIONIK, Iran).
Statistical analyses
Statistical analysis was performed using SPSS version 26.0 (SPSS, Inc., Chicago, IL). Demographic and clinical characteristics were presented as n (%) for definite variables and as mean ± standard deviation for age, which were analyzed with Fisher's exact test and Student's t-test, respectively. All statistical analyses were two-tailed, and p-values <0.05 were considered significant.
Results
Demographic and clinical status of the patients
Finally, 50 COVID-19 patients participated in the study, including 25 outpatients with mild infection aged 55.52 ± 16.91 years with 12 females and 13 males and 25 inpatients admitted to the intensive care unit (ICU) for critical symptoms aged 65.80 ± 11.51 years with 15 females and 10 males. The demographic characteristics of the included patients are shown in the Table 2. The mean age of the inpatients was significantly higher compared with the outpatients (p = 0.028). Twenty-five healthy control subjects who had no symptoms of COVID-19 in the past 3 months were also included. The vaccination history of the patients was recorded, and the COVID-19 patients were monitored for 60 days and their vital status was recorded. A significant difference (p = 0.002) was observed between the vital status 60 days after the onset of symptoms between outpatients and inpatients. Patients who had received vaccination in the history had significantly higher (p = 0.025) survival than nonvaccinated patients.
Demographic Characteristics of the Included Patients and Healthy Controls
Living status is considered within 60 days after symptoms.
SD, standard deviation.
However, no significant difference (p = 0.384) in vaccination status was observed between outpatients and inpatients. Comparison between deceased and surviving ICU patients indicated that vaccination was associated with lower risk of ventilatory failure (p = 0.023). The amount of total cholesterol in serum of control and patients was measured among all groups. The levels of cholesterol were higher in patients with severe disease compared to control and patients with mild infection. However, this difference was not significant (p = 0.212). All results from patient's samples including: groups (inpatients, outpatients, and control), age, and other information have been shown in Supplementary Table S1.
Assessment of CH25H and 25HC
The analysis showed a significant difference in CH25H gene expression between the groups. Expression was significantly higher in outpatients than in inpatients (p = 0.002) and controls (p < 0.001), but no significant difference was found between controls and inpatients (p = 0.171). Gene expression is shown in the Figure 1. Serum levels of 25HC indicated a similar pattern, with levels significantly higher in outpatients compared to inpatients (p = 0.015) and controls (p < 0.001), but the difference (p = 0.092) between controls and inpatients was not significant. Figure 2 shows the mean serum levels of 25HC in the different groups. Overall, there was a significant difference in CH25H (p = 0.001) gene expression and serum 25HC levels (p = 0.001) between patients and controls.
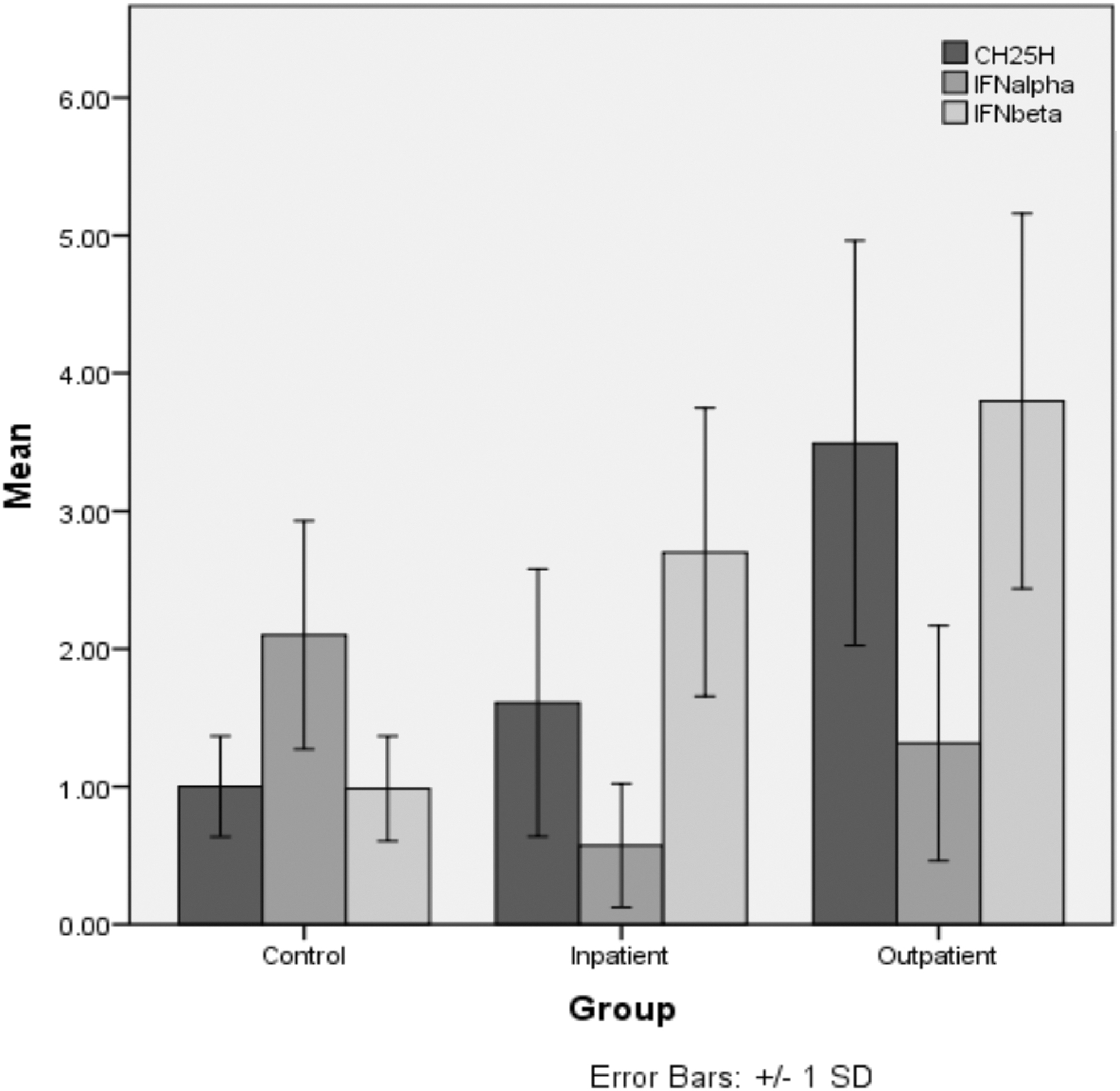
Mean fold change and standard deviation (error bars) in gene expression of CH25H, IFN-α, and IFN-β in the study groups. RT-PCR analysis of CH25H mRNA expression, IFN-α, and IFN-β is compared in control, inpatients, and outpatient group. Data are presented as mean ± SD. As can be seen a difference in CH25H gene expression between the groups is significant. Expression is significantly higher in outpatients than in inpatients (p = 0.002) and controls (p < 0.001), but no significant difference is found between controls and inpatients (p = 0.171). CH25H, cholesterol 25-hydroxylase; IFN, interferon; RT-PCR, real-time polymerase chain reaction; SD, standard deviation.
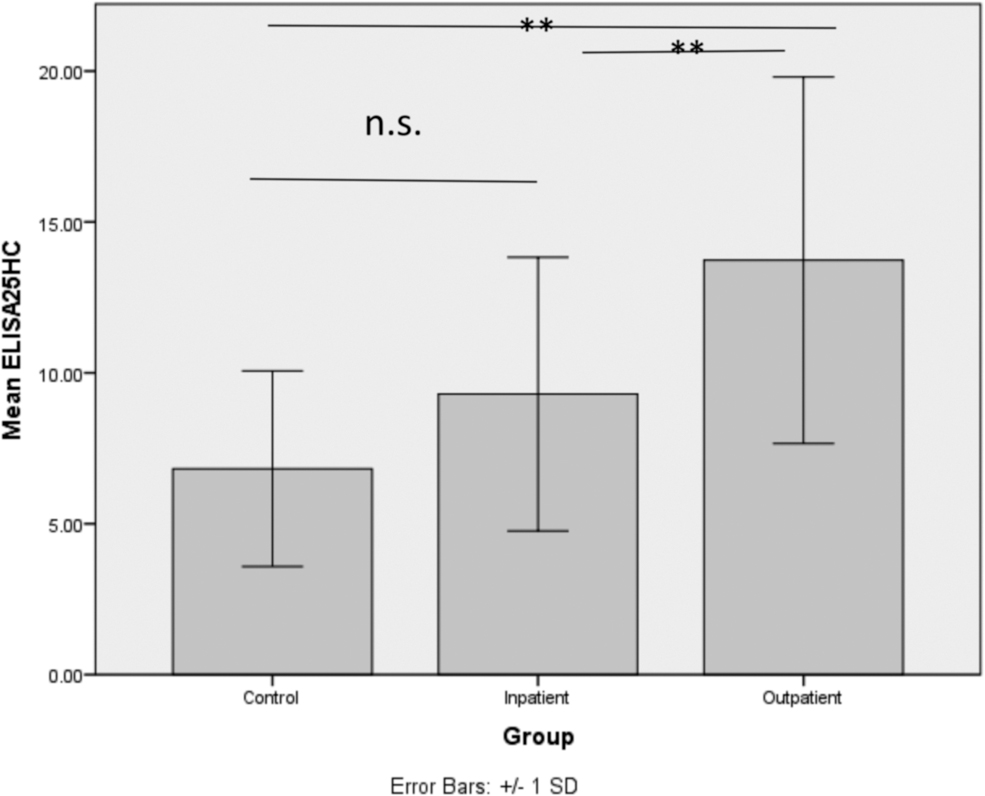
Mean and standard deviation (error bars) of an ELISA measurement of serum levels of 25HC (ng/mL) in the study groups. Serum levels of 25HC indicated a similar pattern, with levels significantly higher in outpatients compared to inpatients (p = 0.015) and controls (**p < 0.001), but the difference (p = 0.092) between controls and inpatients was not significant (n.s.). There was a significant difference in CH25H (p = 0.001) gene expression and serum 25HC levels (**p = 0.001) between patients and controls. 25HC, 25-hydroxycholesterol; ELISA, enzyme-linked immunosorbent assay.
CH25H expression (p = 0.001) and plasma levels of 25HC (p < 0.001) were significantly elevated in surviving patients compared to the deceased patients. Conversely, in patients admitted to the ICU, CH25H expression levels (p = 0.023) and serum 25HC (p = 0.010) levels were significantly lower in the hospitalized who required ventilation.
Cytokine assays
IFN-α was significantly (p = 0.010) higher in outpatients than in inpatients and significantly elevated in controls than in outpatients (p = 0.014) and inpatients (p < 0.001). However, IFN-β was significantly (p < 0.001) increased in outpatients and inpatients but not in controls (p = 0.012), but there was no significant difference between inpatients and outpatients (p = 0.146). Overall, there was a significant difference in IFN-α (p < 0.001) and IFN-β (p < 0.001) between those groups. Gene expression of IFNs is shown in Figure 1.
The gene expression of these two IFNs was compared; no significant difference was observed between surviving and deceased patients for IFN-α (p = 0.293) and IFN-β (p = 0.912).
The analysis showed that there was a correlation between IFN-β and CH25H gene expression (p = 0.008), although not a strong one (Spearman correlation coefficient of 0.353). In contrast, no correlation was found between the expression patterns of IFN-α and CH25H (p = 0.381).
In addition, our data related to the gene expression of TGF-β did not show meaningful results, and we excluded those data from our article.
We wondered whether or not, within the groups, there are correlations between IFN and CH25H mRNA levels. We compared the mean of every variable such as CH25H, IFN-α, and IFN-β in each group and what we observed was that the mean of IFN-α in control group was higher than the mean of CH25H and IFN-β, while CH25H and IFN-β did not show significant difference. The mean of IFN-α in hospitalized patients was lower than the mean of CH25H and IFN-β, while the mean of IFN-β was higher than the mean of CH25H. The mean of IFN-α in patients with mild infection was higher than the mean of IFN-β but was not significantly higher than CH25H.
Discussion
Clinical reports of COVID-19 have found that the severity of COVID-19 disease is related to dysregulation of innate immune responses such as the activity of type I IFNs (Van der Sluis et al., 2022). Type I IFNs exert their antiviral effects by binding to the heterodimeric receptor complex of IFNAR1 and IFNAR2 and stimulating the receptor-associated tyrosine kinases TYK2 and JAK1 signaling pathways, which phosphorylate STAT1 and STAT2. Subsequently, phosphorylated STAT1 and STAT2, in addition to interferon regulatory factor 9, form a trimeric complex named IFN-stimulated gene factor 3, which can enter the nucleus and bind to IFN-stimulated response elements to increase transcription of ISGs (Kim and Shin, 2021). Previous studies have shown that SARS-CoV2 in both in vitro and in vivo is susceptible to this type of IFNs, and therefore, IFN type I can be a promising treatment to control COVID-19 (Radenkovic et al., 2020).
Cholesterol is an important component of the cell membrane and contributes to various cellular activities. Homeostatic pathways and immune responses involved in the control of cholesterol metabolism are mutually regulated (Yvan-Charvet et al., 2019). CH25H is proposed as an ISG that utilizes host cholesterol metabolism for the antiviral immune response by catalyzing the oxidation of cholesterols to 25HC, which acts against viral infections through several mechanisms, including: (1) modifying metabolite biosynthesis to disrupt the viral life cycle; (2) impeding viral replication by attaching to viral protein targets; and (3) regulating the inflammatory response. In mice, the open reading frame (ORF) of CH25H is located downstream of the IFN active site and IFN response elements, and CH25H gene expression increases in response to IFN stimulation (Zhao et al., 2020).
A study in primary human hepatocytes reported that CH25H is not induced after treatment with IFN-α and IFN-γ but is induced by innate immune system adaptation molecules, including MDA5, MAVS, IRF3, and NF-κB (Xiang et al., 2015). However, others showed no association between serum 25HC levels and IFN-γ in hospitalized COVID-19 patients (Asano et al., 2023). In contrast, CH25H was found to be induced by type I IFNs in vitro in animal models in some other studies (Anggakusuma et al., 2015; Park and Scott, 2010). Previous studies demonstrated that immune response in COVID-19 patients with severe status is controlled by TGF-β, as well as IL-21 and IFNs, which trigger switching antibodies to IgA1 and IgG1. While, later on, TGF-β and IL-21 are predominant and lead cells to produce antibody class IgA2 (Ferreira-Gomes et al., 2021).
In our study, we did not observe any significant results related to the status of patients with COVID-19 and the level of TGF-β (data did not show). In this study, we investigated the expression level of CH25H and affected cytokines, as well as the serum level of 25HC among patients in three different groups, including control, inpatients, and outpatients.
Based on our analysis, CH25H and 25HC were higher in COVID-19 outpatients than in healthy controls and hospitalized patients, indicating that SARS-CoV-2 infection is capable of triggering this innate immune response. However, both values were significantly higher in mildly ill patients than in those admitted to the ICU. These results were in contrast to those of Marcello et al. (2020). Serum 25HC levels were found to be elevated in symptomatic pauci/a but not in moderate or severe COVID-19 patients. However, our data were consistent with others (Asano et al., 2023). One explanation for CH25H reduction in patients with severe infections could be the effect of nosocomial drug therapy that reduces the induction of CH25H expression in macrophages. This may be the reason for the slight increase in 25HC levels in hospitalized patients.
In addition, surviving ICU patients had higher CH25H expression and higher serum 25HC levels. In addition, there is evidence that decreased expression of CH25H and decreased production of 25HC are associated with a higher likelihood of mechanical ventilation. Taken together, this evidence suggests that they may play an important role in controlling the spread of SARS-CoV-2 infection. We observed that initial infection with bovine parainfluenza virus type 3 increased CH25H expression. However, this infection caused a decrease in CH25H (Lv et al., 2019). Dong et al. (2018) found that porcine reproductive and respiratory syndrome virus nonstructural proteins (nsp) 1β and nsp11 are the CH25H gene product through the lysosomal pathway in the HEK 293FT cell line.
Therefore, we recommend in vitro studies of possible effects of the SARS-CoV-2 proteins on CH25H. The effect of dietary cholesterol on the expression of CH25H and thus 25HC has been investigated in several studies. Synthesis of 25HC increases very rapidly after a high-cholesterol diet (Russo et al., 2020).
Cholesterol plays a role in SARS-CoV2 virus entry and is important in postattachment processes (Radenkovic et al., 2020). Darrington found that LDL cholesterol, triglycerides, and total cholesterol increased and HDL cholesterol decreased in COVID-19 survivors compared to healthy controls who did not test positive for COVID-19 (Durrington, 2023). For this reason, the study measured total cholesterol levels. We observed different cholesterol levels in the control group and the inpatient and outpatient groups. Patients with mild infections had higher cholesterol levels, whereas controls and patients with severe infections had similar cholesterol levels. Although this difference was not significant, the higher levels of CH25H and, consequently, 25HC may be due to elevated cholesterol levels in patients with severe infections. Of note, patients with mild infections had uncontrolled diets and were on a high-cholesterol diet, which may have resulted in elevated serum cholesterol levels. Critically ill patients presenting to the ICU may not be able to receive normal nutrition. Our controls did not have elevated cholesterol levels compared to hospitalized patients.
This may be due to age group, which may affect cholesterol consumption. According to the literature, extracellular cholesterol induces an immune response into an inflammatory situation. In contrast, endogenous cholesterol plays a role in immune cell differentiation, motility, and function. It has been proven that a diet high in cholesterol can adversely affect inflammatory processes. In contrast, low dietary cholesterol levels reduce excess cholesterol biosynthesis and downregulate inflammatory activation. Studies in mice have found that 25HC can alter B cell responses in animals and humans fed a high-fat diet. Therefore, the exact contribution of different cholesterol sources to the production of 25CH in humans is still unclear, and the exact mechanism needs further investigation (Trindade et al., 2021). 25HC production is influenced by dietary cholesterol levels, but it is unclear whether the source of 25HC is exogenous or endogenous cholesterol. Therefore, the association between decreased 25HC levels and increased total cholesterol levels in the outpatient group should be investigated.
However, it is also important to understand why 25HC levels are high in the outpatient group but total cholesterol levels are low and whether endogenous or exogenous cholesterol affects 25HC levels in different mechanisms. As previously mentioned, mouse CH25H is affected by type I IFNs (Park and Scott, 2010), but debate regarding the ISG nature of CH25H in humans is still ongoing. In this study, we also analyzed the gene expression of type I IFNs. Analysis revealed different expression patterns for IFN-α and IFN-β. IFN-α was more highly expressed in healthy individuals than in patients, consistent with the results of Contoli et al. (2021). They found lower IFN-α level in COVID-19 patients than in controls and also agreed with a cohort study of patients with severe COVID-19 infection that suggested an association between decreased type I IFN expression and complicated condition of COVID-19 infection (Durrington, 2023). We also found that IFN-α was significantly more highly expressed in patients with mild infection.
As IFN-α exhibits a dose-dependent inhibitory effect on SARS-CoV-1 (Ströher et al., 2004), this may explain the association between its expression and disease severity. In particular, low IFN-α expression aggravates the disease caused by SARS-CoV-2. Furthermore, its lack of activity contributes to the severity of COVID-19. IFNAR1 deficiency has been reported to be associated with severe COVID-19 (Marcello et al., 2020). Similarly, autoantibodies against IFN-α may predispose patients to the severe forms of the disease (Bastard et al., 2020). In addition, we hypothesized that high IFN-α expression by itself might exacerbate infection as the SARS-CoV-2 receptor angiotensin-converting enzyme 2 is upregulated by IFN-α (Ziegler et al., 2020).
Given previous data, serum IFN type I levels are elevated in patients who develop hypoxemia and respiratory failure, especially during the early stages of COVID-19 (7 days after onset of clinical symptoms). Another study examined IFN-α levels within 5 days after symptom onset and showed that IFN-α and IFN-β levels were low (Mansourabadi et al., 2023). Other studies suggested that there is mRNA upregulation by both IFNs, but not by IFN-γ in adults aged 16–65 years and in the elderly aged ≥60 years (Asano et al., 2023). They showed that IFN-β may predominate over the systemic immune system in nasopharyngeal samples, but it is still unclear why IFN-α levels elevated during the early stages of SARS-CoV-2 infection (Russo et al., 2020). Based on the results, the low IFN-α levels in the severely ill patient in our results could be attributed to the disease stage at which IFN-α levels were measured in nasopharyngeal specimens.
Thus, the patient sampling stage is important and represents one of the limitations of the current study, while one of the limitations of our study is that we only assessed IFN at one time point of infection, whereas changes in IFN over time in the hospitalized group should be considered. Another limitation is the failure to consider the potential for treatment and the impact of secondary bacterial infections in hospitalized patients. IFN-β was more strongly expressed in patients than in controls and, in contrast to IFN-α, showed a weaker correlation with the CH25H gene. Similar to Berri et al.'s results, patients with mild symptoms had higher IFN-β levels. Patients admitted to the ICU were found to have significantly reduced IFN-β levels (Berri et al., 2023).
Because the SARS-CoV-2 nsp1, nsp3, nsp12, nsp13, nsp14, ORF3, ORF6, and M proteins require activation of the IFN-β promoter, higher IFN-β levels may promote further SARS-CoV-2 infection through increased ACE-2 expression (Nakazono et al., 2021), which may reduce IFN-β levels (Lei et al., 2020). This cycle may explain the reason for decreased IFN-β expression in patients with severe COVID-19. Since IFN-β correlates with CH25H, we hypothesize that this cycle of events may affect CH25H expression and, ultimately, plasma 25HC levels as well. In vitro studies showed that SARS-CoV-2 was more sensitive to IFN-β than to IFN-α.
Overall, consistent with results highlighting the critical role of IFN-β in the immune response to SARS-CoV-2 (Radenkovic et al., 2020), we observed different expression patterns of CH25H and IFN-β in patients, which may explain the effects of CH25H and IFN-β on patient status. Consistent with other studies, we propose that serum levels of 25HC as a biomarker, along with other inflammatory factors affecting CH25H production, may help investigate the severity of COVID-19.
Conclusion
In summary, CH25H and 25HC levels were higher in COVID-19 patients than in healthy controls. Moreover, both values were significantly higher in patients with mild infections than in those admitted to the ICU.
In addition, the surviving ICU patients had higher CH25H expression and serum 25HC levels, indicating the importance of CH25H in preventing severe outcomes. To investigate the potential ISG properties of CH25H in humans, we analyzed the expression level of IFN type I. Our analysis revealed different expression patterns for IFN-α and IFN-β, with no correlation between CH25H and IFN-α, except for a low correlation between CH25H and IFN-β.
Footnotes
Acknowledgment
The authors express their gratitude to the respected participant in this study.
Authors' Contributions
M.R.: Performed Experiments/Reviewing and Editing, M.H.R.: Performed Sample Preparation-Experiments/Conceptualization, H.K.: Performed Reviewing, F.N.: Performed Experiment, S.S.: Performed Sample Preparation and Demographic Information, L.M.: Correspondence/Performed Writing-Original draft and Conceptualization.
Authors' Confirmation Statement
All authors are from the Iran University of Medical Sciences (Tehran, Iran), where education and research are the primary functions.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
The study was financially supported by Iran University of Medical Sciences (Grant Number: 23488).
Supplementary Material
Supplementary Table S1
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.