
Abstract
Vitamin D exerts its antiviral effect through vitamin D receptor (VDR)/retinoid X receptor-mediated host immunomodulation. Besides the downregulation of VDR expression, its polymorphism was also observed among hepatitis B virus (HBV)-positive patients. To understand the possible link between VDR polymorphism and its altered expression during HBV infection and disease progression, VDR Apa-I [rs7975232 (C>A)] single nucleotide polymorphism (SNP) was analyzed in a case–control manner. VDR Apa-I (rs7975232, C>A) polymorphism was studied using 340 HBV patients and 102 healthy controls. Genotype analysis and gene expression study was performed using restriction fragment length polymorphism and quantitative polymerase chain reaction, respectively. Statistical analysis was performed using SPSS (IBM) considering p-value <0.05 as significant for comparing the differences between the groups. Significant mean difference in VDR expression was observed between HBV-positive patients (1.6 ± 0.94) and controls (0.69 ± 0.73). Furthermore, the mean fold change of Healthy control with CC genotype (1.92 ± 0.99) was found to be marginally significant compared with mutant genotype (CA/AA) (1.08 ± 0.43/0.59 ± 0.56, p = 0.045). In HBV+ patients, the mean fold change in the CC genotype was 0.88 ± 0.38, which exhibits a significant mean difference upon comparison with other genotypes (0.52 ± 0.49, 0.113 ± 0.34; p = 0.018, p = 0.048). However, the fold change value does not differ between CA and AA genotypes. Further comparative analysis of VDR expression between the control and case also exhibits significant differences (p = 0.001) among allelic variants. Observed genotype distribution frequency exhibits a significant association with disease type. The mutant genotype was found to be significantly associated with HBV infection and disease progression, (odds ratio = 0.730, 95% confidence interval = 0.462–1.152, p = 0.06). VDR SNP rs7975232 (C>A) may affect VDR expression by controlling several other variables and suggest that deviation from wild-type genotype (CC) is associated with downregulation of expression, which in turn involved in host immunomodulation in favor of HBV infection and disease progression.
Introduction
Hepatitis B virus (HBV) can cause a serious viral infection that primarily affects the liver. The virus is transmitted through contact with infected bodily fluids, including blood, semen, and vaginal secretions. HBV infection can lead to acute or chronic hepatitis (CH), liver cirrhosis, and liver cancer. According to WHO Report, it is estimated that around 296 million people are living with chronic HBV infection worldwide (Hsu et al., 2023). Vitamin D receptor (VDR) encodes the VDR protein, which plays a crucial role in the metabolism of vitamin D. Vitamin D and its metabolites have been shown to have immunomodulatory effects and the VDR is an important transmodulator of immune function (Gotlieb et al., 2018). In addition, VDR activation has been shown to inhibit HBV replication in vitro and in vivo, suggesting a direct antiviral effect of vitamin D (Ahluwalia et al., 2021; Gotlieb et al., 2018). Whereas, altered expression of VDR has been observed in patients with HBV infection (Hoan et al., 2016; Yang et al., 2022). Several studies have reported that VDR expression is reduced in the liver tissue of patients with CH B (Yang et al., 2022).
The reduced VDR expression in HBV infection is thought to play a role in the pathogenesis of the disease. Furthermore, vitamin D deficiency has been associated with an increased risk of HBV infection and a more severe disease course in patients with CH B. A study conducted in China found that vitamin D deficiency was more common in patients with CH B than in healthy controls and that vitamin D supplementation improved liver function in these patients.
VDR gene comprises 8 protein-coding exons and 6 untranslated exons along with intervening intron sequences and makes up to 75 kb sequence of chromosome 12 (Li et al., 2015; Miyamoto et al., 1997). Exons 2 and 3 are necessary for DNA binding and Exons 7, 8, and 9 are involved in the intracellular binding of Vitamin D and altogether form the VDR structural protein (Hughes et al., 1988).
Past research revealed different allelic variants of the VDR gene with significant characteristics, such as Bsm-I (rs1544410, C>T), Tru-9 (rs757343, G>A), and Apa-I (rs7975232, C>A) in intron 8, Taq-I (rs731236, A>G) in exon 9, and Fok-I (rs10735810, C>T) in exon 2. These single nucleotide polymorphisms (SNPs) have been reported to have an association with various important diseases (Uitterlinden et al., 2004). VDR Apa-I polymorphism, which involves a change from cytosine to adenine (C>A), is linked with lumbar disc degeneration (Yuan et al., 2010), while another study suggested the possible correlation between the Apa-I polymorphism and the susceptibility of HBV infection, as well as the potential clinical outcomes of such an infection (Hoan et al., 2019).
A past study conducted in Taiwan found that individuals with the A allele of the Apa-I polymorphism had a higher risk of developing cirrhosis and hepatocellular carcinoma (HCC) in patients with chronic HBV infection (Huang et al., 2010). Whereas, a similar study on the Chinese population revealed that the rs2228570 SNP in the VDR gene was associated with an increased risk of chronic HBV infection and liver cirrhosis in the Chinese population (Peng et al., 2014). However, the relationship between differential VDR expressions with its allelic variants during HBV infection is still not well understood. This study therefore focuses to understand the role of VDR Apa-I polymorphism and VDR gene expression with the risk of HBV infection and disease progression.
Materials and Methods
Study design and specimen collection
This is a cross-sectional study, which includes 340 HBV-infected patients attending the outpatient department of the Department of Gastroenterology, Gauhati Medical College and Hospital within 3 years. Depending on the type of clinical incarnation, patients were further stratified into three distinct clinical subgroups viz. Acute viral hepatitis (AVH, N = 205), CH (CH, N = 84), and HCC (HCC, N = 51). All the patients exhibit HBsAg +ve status, elevated liver enzyme aspartate aminotransferase (AST)/alanine transaminase (ALT), nausea, and vomiting as the common characteristic of HBV-related liver disease. Additionally, elevated serum Anti-HBe and HBeAg-Anti HBe immune complex besides persistent HBsAg positivity for 6 months and beyond were the discriminating characteristics for CH from other types. HCC patients were confirmed based on various computed tomography scanning approaches and histopathological grading of liver tissue. Individuals from all age groups irrespective of gender were included in the study. Whereas patients with other than HBV-related liver disease, with an autoimmune disorder, and HIV-infected individuals were excluded from the study.
Besides the HBV-negative status, a similar exclusion criterion was also followed while recruiting 102 healthy controls in the study. The samples were collected after obtaining proper informed consent from the individuals. The study was approved by the institutional Ethics Committee of Gauhati Medical College and Hospital, Guwahati-14, Assam (vide no MC/190/2007/Pt-II/DEC-2020/43). Approximately 5 mL of blood samples were collected aseptically from each of the individuals and divided into two parts. One part is in a clot activator vial for serum separation and the other is in EDTA viral for DNA/RNA extraction.
Detection of VDR allelic variants at Apa-I locus (rs7975232)
Isolated DNA from blood samples was subjected to VDR polymorphism analysis through polymerase chain reaction (PCR)–restriction fragment length polymorphism following quantification of extracted nucleic acid. For amplification of the Apa-I locus forward primer 5′-GCTTCTGGATCATCTTGGCA-3′ and reverse primer 5′-ATGGACAGAGCATGGACAGG-3′ were used, which will produce a 402 bp amplicon from VDR gene. PCR amplification of the target region was carried out in a 20 μL reaction mixture containing 1 U Taq polymerase (Thermo Scientific), 2 μL 10 × Taq Buffer, 0.4 mM dNTP mixture, ∼200 ng extracted DNA, and 1.6 mM MgCl2. After mixing all the components, the thermal cycler was set for an initial denaturation at 95°C for 5 min followed by 35 cycles of denaturation at 95°C for 30 sec, annealing at optimum temperature, and extension at 72°C for 45 sec with a final extension step at 72°C for 5 min. The PCR amplicons were subjected to restriction digestion using Apa-I enzyme by overnight incubation at 37°C. In the presence of wild-type genotype (CC) at rs7975232 Apa-I digestion produces fragments of length 238 and 164 bp.
Expression analysis of VDR gene
Total RNA was extracted using a commercial kit (QIAamp RNA Blood Mini Kit, QIA-GEN, Germany) following the manufacturer's protocol, and 1 μg of RNA was converted to cDNA using a cDNA Synthesis Kit (Verso cDNA Synthesis Kit; Thermo Scientific). Expression analysis of the VDR gene was performed in ABI QuantStudio3 Real-time PCR detection system using the SYBR green (RR420A TaKaRa) method following the manufacturer's instruction. The relative expression level of the gene was estimated by taking the expression level of the housekeeping gene β-actin as a reference. The 2−ΔΔCt method was used to manually estimate the expression level of VDR (Livak and Schmittgen, 2001).
Statistical analysis
Statistical analysis was performed using SPSS version 21.0 (IBM) for the Windows platform (IBM Corp., Armonk, NY). All the categorical variables are presented as numbers or ratios and the continuous variables as mean ± standard deviation. A chi-square test was performed to find the differences in nominal data among the different groups. One-way analysis of variance was performed to compare the mean difference in continuous variables. Chromatogram obtained after Sanger sequencing was visualized and aligned against the human VDR reference sequence from NCBI (NG_008731.1 C>A) using Mega X (Kumar et al., 2018). Gene-Calc web-based platform was used (
Results
Clinicopathological characteristics of the study population
Clinicopathological characteristics of the study population are described in Table 1. Average age does not differ significantly (p = 0.137) between the healthy control and HBV+ group, however, upon comparison within the HBV+ group based on the degree of disease severity, mean age was found to differ significantly (p = 0.001). The occurrence of the disease was not found to be associated with gender (Male: Female). Significant reduction in platelet count was observed among HBV+ patients (100.46 ± 40.9) upon comparison with healthy control (232.07 ± 60.97) and a similar pattern was also observed with advanced stages of the disease (AHV, CH, and HCC; p < 0.05). Elevation of the liver enzyme (AST/ALT) was significant among the positive patient group (83.25 ± 27.9/89.71 ± 36.86) upon comparison with healthy control (27.48 ± 10.18/35.92 ± 10.51; p = 0.001). Serum albumin levels exhibit an overall significant reduction with HBV infection (3.98 ± 0.81) and the pattern was also observed with the advancement of the disease. However, HBV viral load does not differ significantly (p > 0.05) among the positive patient group upon stratification based on the degree of disease severity.
Comparison of Clinicopathological Characteristics Between Hepatitis B Virus+ Patients and Healthy Control
Bold values represents p value <0.05, which exhibit significance of the mean difference at 95% level of confidence.
p1: compares AVH and CH; p2: compares AVH and HCC; p3: compares CH and HCC.
ALT, alanine transaminase; AST, aspartate aminotransferase; AVH, acute viral hepatitis; CH, chronic hepatitis; HBV, hepatitis B virus; HCC, hepatocellular carcinoma.
Analysis of VDR gene expression in HBV+ patients and healthy controls
Analysis of relative VDR gene expression (Table 2) exhibits an ∼2.3 times higher fold change value among the healthy controls (1.6 ± 0.94) in comparison to the HBV+ patients (0.69 ± 0.73) and the mean fold change difference was statistically significant (p = 0.001). Further analysis of expression within the HBV+ patients stratified based on disease severity, exhibited mean fold change of AVH patients as 0.77 ± 0.08, CH as 0.693 ± 0.53, and that of HCC patients as 0.35 ± 0.09. The reduction in mean fold change does not differ significantly between acute and chronic patients (p = 0.409), however comparison with HCC patients, the reduction in VDR gene expression was found to be highly significant for both acute and chronic HBV patients (p = 0.001) (Figs. 1 and 2).

Expression of VDR gene in HBV-positive cases and healthy control (mean ± SD). The box plot has been generated using SPSS, and the * represents extreme outliers. HBV, hepatitis B virus; SD, standard deviation; VDR, vitamin D receptor.
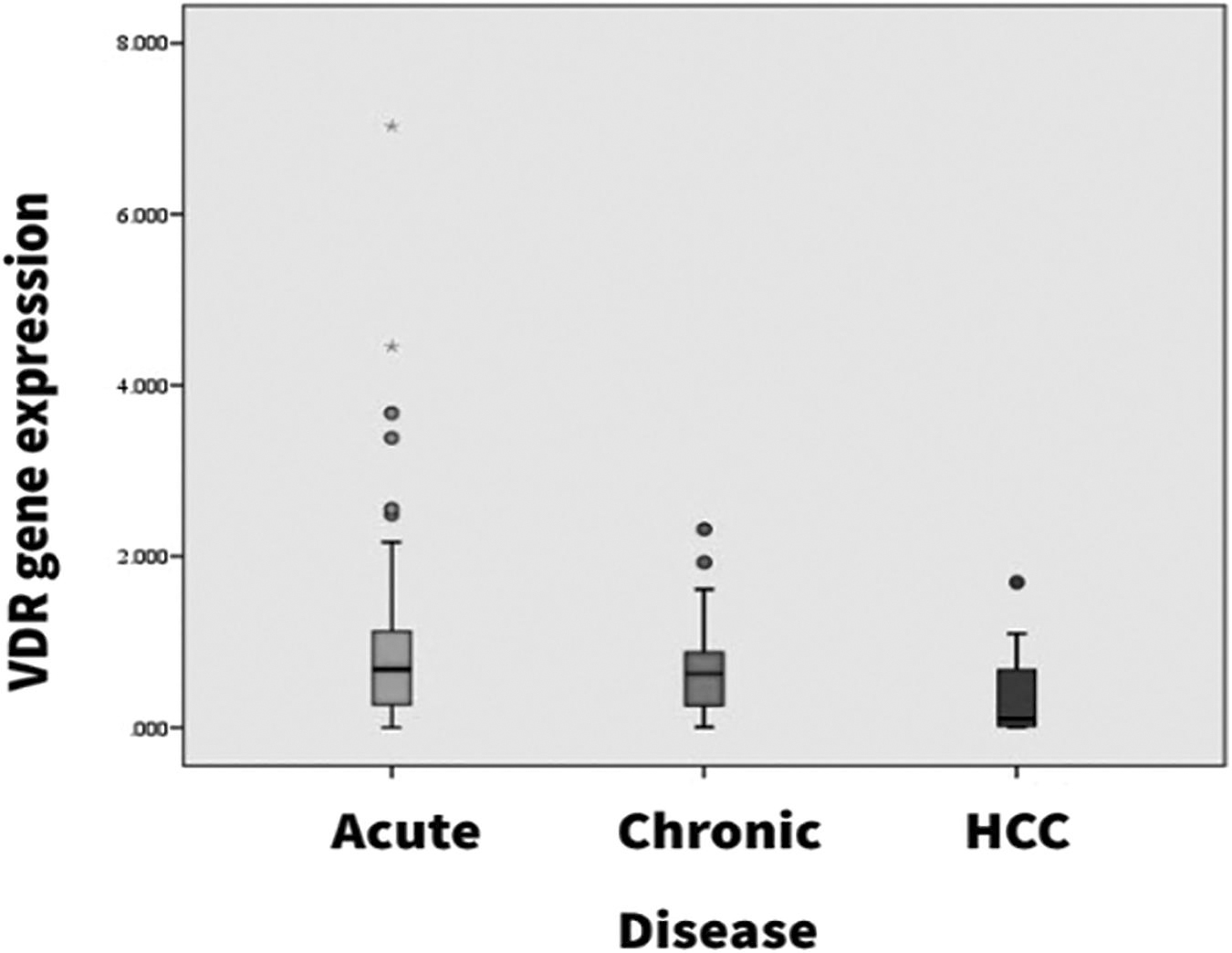
Differential expression of VDR gene in types of HBV-related liver disease within the study population (mean ± SD). The box plot has been generated using SPSS, and the * represents extreme outliers.
Comparison of Mean Vitamin D Receptor Expression Between Healthy Control and Hepatitis B Virus+ Patients and its Association with Types of Hepatitis B Virus-Related Liver Disease
Bold values represents p value <0.05, which exhibit significance of the mean difference at 95% level of confidence.
p1: compares AVH and CH; p2: compares AVH and HCC; p3: compares CH and HCC.
Indicating p value <0.05, which is statistically significant at 95% level of confidence.
CI, confidence interval; SD, standard deviation; VDR, vitamin D receptor.
VDR SNP Apa-I (rs7975232) genotype distribution frequency
The genotype distribution pattern obtained was found to be in Hardy–Weinberg equilibrium. Wild-type genotype (CC) was the dominant genotype among both healthy control (63.7%) and HBV+ patients (56.17%) meanwhile the frequency of rare homozygous genotype (AA) was the least. The frequency of AA genotype among HBV+ patients was 7.94% and that among healthy control was 4.9% (odds ratio [OR] = 0.706, 95% confidence interval [CI] = 0.252–1.979). The heterozygous (CA) genotype distribution frequency among the HBV+ group was 35.88% and among healthy control 31.3% (OR = 0.544, 95% CI = 0.201–1.472). Considering wild-type genotypes as a reference, the genotype distribution frequency of heterozygous (CA) and rare homozygous (AA) genotypes was found to be statistically significant (p = 0.001). In the dominant model of genotype distribution, mutant genotype (CA/AA) was found in 43.82% of HBV+ cases, whereas 36.2% of the healthy control possess the same mutant genotype (CA/AA).
Upon comparing with wild-type genotype (CC), the distribution pattern was found to be marginally significant (OR = 0.730, 95% CI = 0.462–1.152, p = 0.06). In the recessive model of genotype distribution, the rare homozygous genotype (AA) upon comparison with CA/CC genotype exhibits a nonsignificant association with HBV infection (OR = 0.598, 95% CI = 0.224–1.594, p = 0.304) (Table 3).
Genotype Distribution Pattern of Vitamin D Receptor Single Nucleotide Polymorphism rs7975232 (C>A) and its Association with Hepatitis B Virus Infection and Types of Hepatitis B Virus-Related Liver Disease
Bold values represents p value <0.05, which exhibit significance of the mean difference at 95% level of confidence.
Indicating p value <0.05, which is statistically significant at 95% level of confidence.
p < 0.05 was considered significant.
OR, odds ratio.
Further analysis of the genotype distribution pattern among the HBV+ patients, stratified based on disease severity (AVH, CH, and HCC), the frequency of AA genotype was found to be highest among HCC cases (11.7%) in comparison to AVH (8.7%) and CH (3.5%). The distribution pattern exhibits significant association upon comparison with the types of disease (OR = 0.972, 95% CI = 0.36–2.64, p = 0.004 and OR = 2.78, 95% CI = 0.65–11.8, p = 0.038, respectively). In the dominant model of genotype distribution analysis, the mutant genotype (CA/AA) was also found to be significantly associated with HCC cases upon comparison with AVH (OR = 0.438, 95% CI = 0.27–0.84, p = 0.014) and CH cases (OR = 0.613, 95% CI = 0.29–1.28, p = 0.019). However, the recessive model of genotype distribution pattern did not exhibit any significant association with types of liver disease.
Association of VDR expression with rs7975232 in HBV-related liver disease and its types
The VDR gene expression in healthy control and HBV+ patients can be seen in Figure 3. An overall reduction in mean fold change value has been observed among the HBV+ group, which is statistically significant upon comparison with Apa-I (rs7975232) genotype-stratified healthy controls. Within the HBV+ patients with wild-type genotype, the reduction in overall mean fold change differs significantly (p = 0.001) upon comparing AVH (1.01 ± 0.99) and CH (0.94 ± 0.52) patients with HCC (0.44 ± 0.47) patients. Patients with the CA genotype exhibit significant mean differences only among AVH and CH groups (p = 0.011). Furthermore, patients with rare homozygous genotype (AA) exhibit significant mean fold change differences across the types of HBV-related liver disease (Table 4).
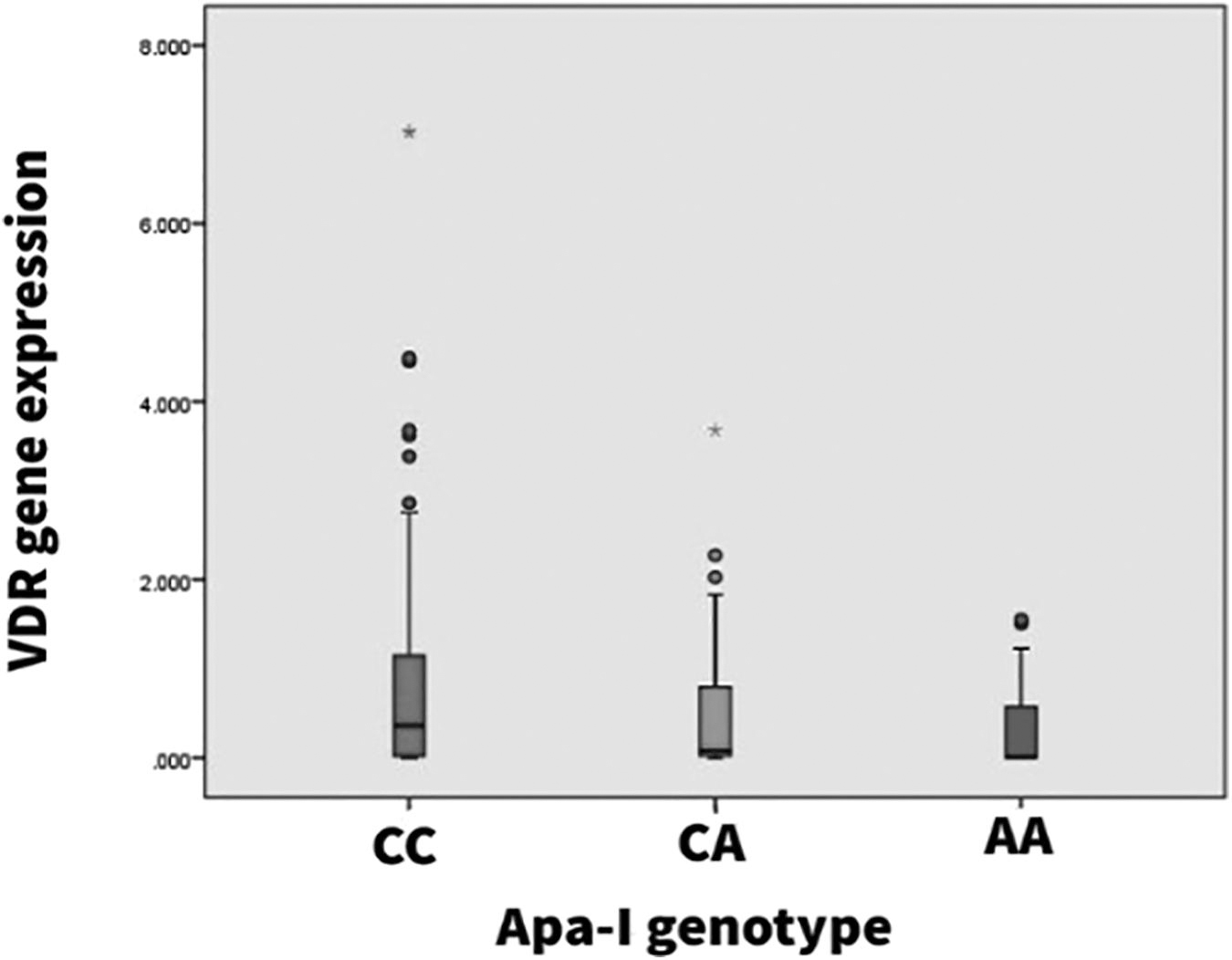
Differential expression of VDR gene with Apa-I allelic variants rs7975232 (C>A) (mean ± SD) among HBV-infected patients. The box plot has been generated using SPSS, and the * represents extreme outliers.
Association of Vitamin D Receptor Gene Expression with Single Nucleotide Polymorphism rs7975232 (C>A) in Hepatitis B Virus-Related Liver Disease and its Types
Bold values represents p value <0.05, which exhibit significance of the mean difference at 95% level of confidence.
Indicating p value <0.05, which is statistically significant at 95% level of confidence.
p < 0.05 was considered significant.
Analysis of VDR gene expression within individual study groups across all the observed Apa-I (rs7975232) genotypes also exhibits a similar reduction pattern in mean fold change value as the genotype deviates from the wild type. Even among the healthy control group, the mean fold change in wild-type genotype (1.92 ± 0.99) was found to be marginally significant upon comparison to the mutant genotype (CA/AA) (1.08 ± 0.43/0.59 ± 0.56, p = 0.045). In the HBV+ patient group, the mean fold change in the CC genotype was 0.88 ± 0.38, which exhibits a significant mean difference upon comparison with other genotypes (0.52 ± 0.49, 0.113 ± 0.34; p = 0.018, p = 0.048) (Table 5). However, the fold change value does not differ significantly between CA and AA genotypes. Furthermore, analysis of the mean fold change difference was found to be significant across all the disease types upon the comparison of wild-type genotype with the mutated genotype (Figs. 3 and 4).
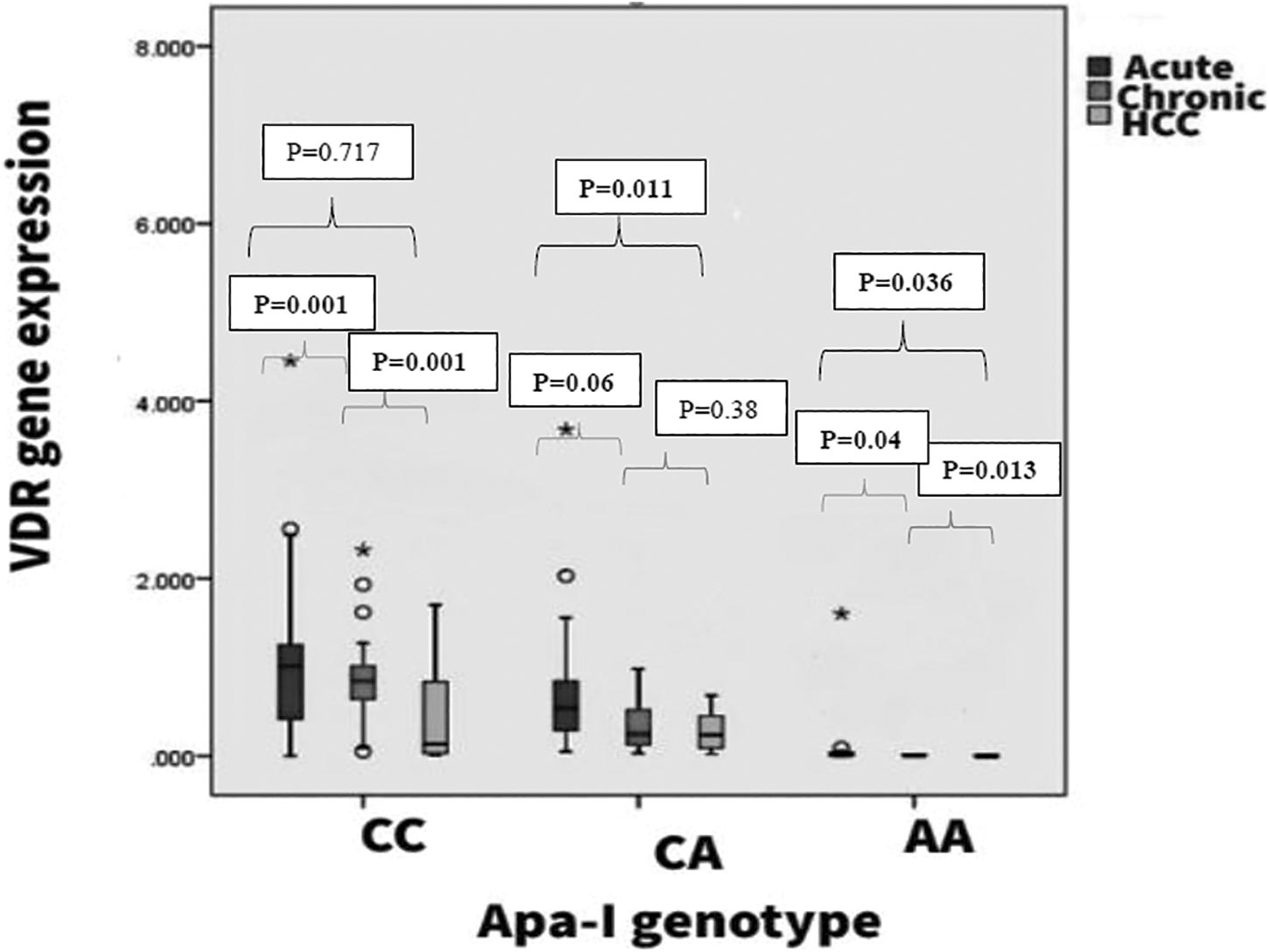
Alteration in mean expression of VDR with types of HBV-related liver disease stratified based on Apa-I genotype (rs7975232). The box plot has been generated using SPSS, and the * represents extreme outliers.
Comparison of Mean Vitamin D Receptor Expression with rs7975232 (C/A) Allelic Variants Among Control and Hepatitis B Virus+ Group
Bold values represents p value <0.05, which exhibit significance of the mean difference at 95% level of confidence.
Factors associated with HBV-related liver disease and its types by logistic regression analysis
Multinomial logistic regression analysis showed AST (OR = 1.318, 95% CI = 1.131–1.536, p = 0.001), ALT (OR = 1.253, 95% CI = 1.072–1.405, p = 0.005), platelet count (OR = 0.940, 95% CI = 0.920–0.962, p = 0.001), altered VDR gene expression (OR = 1.250, 95% CI = 0.869–1.836, p = 0.001), and VDR genotype at rs7975232 (CA/AA) (OR = 0.577, 95% CI = 0.33–0.98, p = 0.049) as independent disease predictors. Further analysis among disease groups considering AVH as a reference group, the increasing age of individuals (OR = 1.118, 95% CI = 1.084–1.154, p = 0.001) apart from the above-mentioned factors acts as independent predictors for disease progression also (Table 6).
Association of Clinicopathological Factors and Types of Hepatitis B Virus-Related Liver Disease Among Hepatitis B Virus-Positive Patients in Multivariate Regression Analysis
Bold values represents p value <0.05, which exhibit significance of the mean difference at 95% level of confidence.
SNP, single nucleotide polymorphism.
Discussion
The antiviral role of 1, 25-dihydroxy vitamin D3 involves VDR/retinoid X receptor mediated host immunomodulation and thus VDR acts as the sole transducer for regulating the expression of nearly 3% of human genes (Yang et al., 2022). As such, the alteration of VDR expression itself plays an important role during viral infection. A negative association between hepatic VDR expression and hepatic inflammation cum fibrosis among patients chronically infected with HBV has already been reported by (Yang et al., 2022). Another study reported significant downregulation of VDR expression in the HBV-transfected HepG2 cell line, which prevents vitamin D from inhibiting viral transcription and translation (Gotlieb et al., 2018). Downregulation of VDR makes the receptor unable to provide efficient signaling for the expression of downstream effector molecules, which maintains the normal calcitriol level (Miao and Goltzman, 2021). Increased calcitriol levels may reduce VDR competency, apart from the suppression of macrophage activity and the kappa-β pathway (Mangin et al., 2014). HBV uses this molecular mechanism to evade the host immune system.
Significant reduction in VDR mRNA level among HBV+ patients thus indicates hindrance in the activation of innate immune cells as well as inflammatory cytokines (Maruthai et al., 2022). In this study, VDR gene expression was reduced by approximately two to three times among HBV+ patients to that of healthy controls, which is statistically significant and is consistent with previously reported findings. Furthermore, the reduction in mean fold change was in direct association with an increasing degree of disease severity. Thus, altered VDR expression may itself act as an immunomodulator in favor of HBV infection and disease progression or it may be a result of HBV infection.
Recent studies suggest the influential effect of VDR polymorphism on altered VDR expression in addition to many other host factors (Hitman et al., 1998; Jurutka et al., 2000; Panda et al., 2019). VDR polymorphism, particularly Apa-I, Taq-I, and Fok-I, imparts reduced mRNA stability and thereby hinders intracellular VDR protein levels, which will affect the Th1–Th2 cytokine balance important for providing an innate antiviral response (Santos et al., 2018). In this study, VDR Apa-I polymorphism (rs7975232) was observed to be associated with HBV+ patients, and noted that CA (35.88%) and AA (7.94%) genotype was significantly (p = 0.001) more prevalent in comparison to healthy control (31.3% and 4.9%, respectively). The present study results suggest probable Apa-I polymorphism association with HBV infection and disease severity. Previous studies also suggest the significant association of the Apa-I CA/AA genotype with increased serum bilirubin level, elevated liver enzyme, and HBV DNA among chronic HBV patients from Vietnam (Hoan et al., 2019). In our study, within the HBV+ patients, the frequency of the AA genotype was highest in HCC patients.
The frequency of the CA genotype was highest in AVH cases and the frequency decreased with increasing degree of disease severity, which indicates a probable association of VDR Apa-I polymorphic deviation from wild-type genotype and the type of HBV-related liver disease.
We further observed an overall downregulation of VDR expression among HBV+ patients in comparison to healthy controls across all the genotypes for rs7975232. Within the HBV patients, the VDR fold change decreases with deviation from wild-type genotype showing the lowest fold change value with the AA genotype. The difference in mean fold change between observed genotypes was statistically significant. Furthermore, a significant mean fold change difference was also observed with changing genotypes for the said VDR SNP in different types of liver diseases (AVH, CH, and HCC). This indicates a probable involvement of VDR SNP in the alteration of VDR m-RNA expression mainly by hindering mRNA stability resulting in host immunomodulation favoring HBV infection and disease progression.
Intronic SNP plays an important role in maintaining mRNA stability (Akdeli et al., 2014). The studied SNP rs7975232 (Apa-I, C>A) located within intron 8 at the 3′ UTR region of the VDR gene has also been reported to exhibit an association with its transcript stability (Zhang et al., 2020) apart from other SNPs (Fang et al., 2005). As such, the formation of Apa-I allelic variants due to SNP resulting in downregulation of VDR expression may attenuate the VDR and its downstream function, and thereby enhance susceptibility to viral infection (Areeshi et al., 2017).
Overall, our study showed a significant mean fold change difference of VDR among HBV+ patients with varied degrees of liver disease. A significantly higher expression of the VDR gene was observed in the CC genotype upon comparison to CA/AA genotype. Patients with the AA genotype exhibit a very low level of VDR expression (0.113 ± 0.34), whereas the CA genotype exhibits a moderate expression (0.52 ± 0.49) and as such increases the risk of HBV infection and disease progression. A similar pattern of downregulation of VDR gene expression was also observed upon segregation of the positive patients based on the type of liver disease. The observation indicates the impact of rs7975232 polymorphism on VDR expression controlling several other variables and suggests that deviation from wild-type genotype (CC) is associated with the downregulation of expression.
Further verification is advocated to determine whether the studied SNP plays a direct role in regulating the expression of the VDR, affects the stability of its transcripts, or is simply in disequilibrium with a VDR activity-regulating SNP. This could be achieved by studying a possible larger sample size to ensure accuracy and reliability.
As such, VDR Apa-I polymorphism (rs7975232) may act as a prognostic marker for HBV infection and disease types due to its association with altered VDR expression, which in turn modulates the host antiviral response machinery in favor of HBV infection and disease progression.
Conclusion
VDR gene expression is associated with HBV infection and with types of HBV-related liver disease. A significant reduction in mean expression was observed with increasing disease severity in comparison to healthy controls. Further analysis of VDR SNP rs7975232 (C>A) exhibits significant mean expression difference of VDR with other than wild-type allelic variants (CA/AA). The allelic variants were also found to be associated with HBV infection and degree of disease severity. Indeed, it can be concluded that VDR intronic SNP rs7975232 (C>A) may play an important downregulatory role in its expression, which in turn is involved in host immunomodulation in favor of HBV infection and disease progression.
In the present study, the VDR SNP [rs7975232 (C>A)] analysis exhibits significant results, where we observed a substantial difference in mean expression between the wild-type allelic variant and mutant allelic variants. These allelic variants were also found to be associated with HBV infection and the degree of disease severity. The expression of the VDR gene is altered in response to HBV infection and correlates with the severity of liver disease. The findings suggested that the VDR intronic SNP rs7975232 (C>A) may play a crucial role in downregulating its expression, which, in turn, could be involved in host immunomodulation that favors HBV infection and contributes to disease progression. In conclusion, the study found that as the severity of the disease increased, there was a significant reduction in the mean expression of the VDR gene compared with healthy individuals. However, we highly advocate for these findings to analyze and have a deeper understanding of the mechanism through which VDR and its allelic variation affect the development of liver disease.
Authors' Contributions
M.J.K1. and S.K1. wrote the article. M.J.K1. and P.P.D1. designed the work plan, including methodology and primers to be used. M.J.K1, J.B.1,2, A.J.D.,1 and K.D1. performed all the molecular wet laboratory work at L.M.V.O., Department of Bioengineering and Technology, Gauhati University. M.J.K.1, G.H.,1 and S.K1. prepared the tables and figures. B.N.C.2, M.G.I.1, and S.M.1 performed language editing and arrangement of the article. All authors reviewed the article.
Consent to Participate
Informed consent was obtained from all individual participants included in the study either in self or from attendants.
Ethics Approval
The study has been ethically approved by the institutional Ethics Committee of the Gauhati Medical College and Hospital, Guwahati, Assam-781032 vide no MC/190/2007/Pt-II/DEC-2020/43 and by the IEC of Gauhati University, Guwahati, Assam-781014.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
No funding was received for this article.