
Abstract
Hepatitis E virus (HEV) exhibits tropism toward hepatocytes and thus affects the liver; however, HEV may also affect other tissues, including the heart, kidneys, intestines, testicles, and central nervous system. To date, the pathophysiological links between HEV infection and extrahepatic manifestations have not yet been established. Considering that HEV infects multiple types of cells, the direct effects of virus replication in peripheral tissues represent a plausible explanation for extrahepatic manifestations. In addition, since the immune response is crucial in the development of the disease, the immune characteristics of affected tissues should be revisited to identify commonalities explaining the effects of the virus. This review summarizes the most recent advances in understanding the virus biology and immune-privileged status of specific tissues as major elements for HEV replication in diverse organs. These discoveries may open avenues to explain the multiple extrahepatic manifestations associated with HEV infection and ultimately to design effective strategies for infection control.
Introduction
With an estimated 20 million cases annually, hepatitis E virus (HEV) is a causative agent of endemic and epidemic liver disease worldwide and is consequently an important public health concern, especially in immunocompromised and pregnant populations (Tian et al., 2022). Beyond the liver, HEV can infect other tissues, including the heart, kidneys, intestines, testicles, and central nervous system, resulting in multiple extrahepatic manifestations.
HEV is a single-stranded, positive-sense RNA virus composed of 7.2 kb of capped and polyadenylated RNA (Nagashima et al., 2017) that encodes three main open reading frames (ORFs): ORF1 encodes nonstructural proteins that are important for virus replication (Nagashima et al., 2017), ORF2 is translated into a protein that forms the viral capsid, and ORF3 encodes a small protein described as being important for the egress of virions from infected cells (Glitscher and Hildt, 2021).
This virus belongs to the Paslahepevirus genus within the Hepeviridae family. The Paslahepevirus balyani species affects humans and comprises eight genotypes (gt). Two genotypes, gt1 and gt2, represent a special category, as they are restricted to infecting humans (Chapuy-Regaud et al., 2017), while genotypes 3, 4, and 7 infect both humans and diverse animal species. Gt5, gt6, and gt8 have been identified in a wide variety of wild and domestic animals, including pigs, wild boars, camels, and rabbits; and gt7 has been identified in camels. Zoonotic abilities have been demonstrated in gt3, gt4, and gt7, with transmission through pigs being the most studied (Kamani et al., 2021).
The transmission of HEV gt3 and gt4 is commonly zoonotic in developed countries, whereas enteric transmission is frequent for gt1 and gt2 in developing regions. All genotypes can be transmitted via the fecal–oral route through contaminated water and food, from mother to fetus, from person to person, and through blood transfusions (Fig. 1A) (Bi et al., 2020). Most HEV contacts result in asymptomatic infections and cause self-limited diseases, and in developed countries, acute HEV infection contributes to 5–15% of acute liver failure (Sayed et al., 2021b). Currently, the mortality rate for acute infection of the virus is 0.2–4%, but this will depend on risk groups such as pediatric and pregnant women or those patients with a history of liver disease in whom mortality is higher (Webb and Dalton, 2019).
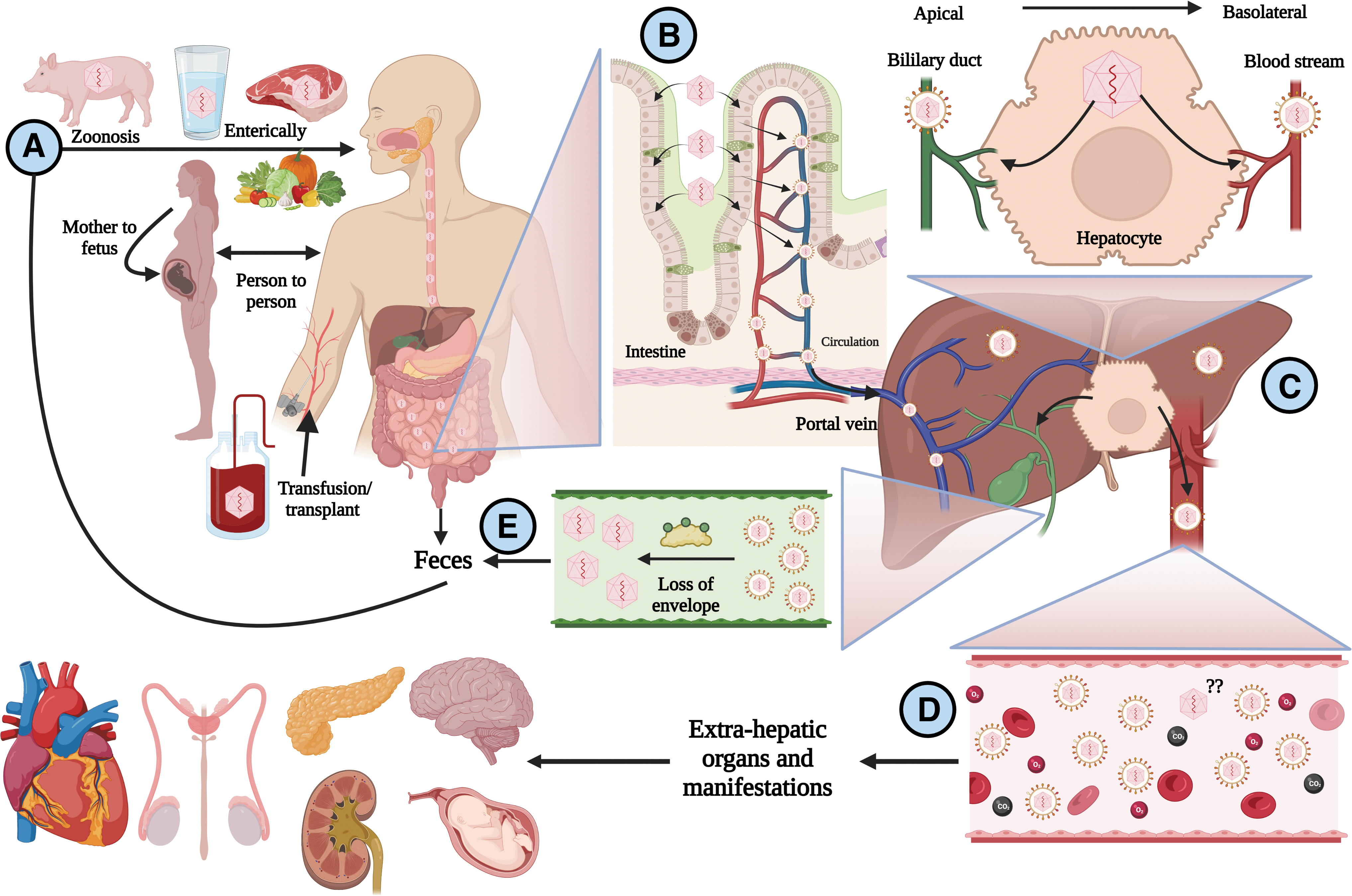
Life cycle of HEV.
In immunocompromised patients, gt3 and gt4 HEV infections may progress to chronic hepatitis and cirrhosis. It is accepted that liver damage and progression of the disease resulting from infection are also influenced by the immune response (Damiris et al., 2022). Moreover, acute liver failure during pregnancy attributed to gt1 and gt2 seems to also be associated with the host's immune conditions (Pérez-Gracia et al., 2017). However, how a specific immune response against the virus affects the variability in the course of the infection has not yet been determined in detail.
In the last two decades, reports of extrahepatic manifestations linked to HEV infection have been increasing. However, the pathophysiological mechanisms behind their development remain to be elucidated. Herein, we summarize the most recent advances in the understanding of HEV biology and the penetration of immune-privileged barriers by virions. These discoveries might explain the presence of extrahepatic manifestations during infection.
Naked Versus Quasienveloped HEV Infection
HEV exists in two infectious conformations: naked (nonenveloped virions, found in bile and feces) and quasienveloped (host membrane-cloaked virions found in bloodstream). These forms have different ways to infect cells. HEV has been proposed to first replicate in intestinal cells and to be subsequently released in its quasienveloped form (q-HEV) into the portal bloodstream, from which it can infect the liver (Fig. 1B) (Marion et al., 2020). When q-HEV virions infect hepatocytes and RNA is duplicated, the virions that are released at the basolateral side of hepatocytes will retain their envelope and then will be delivered through the bloodstream until they find a target organ to infect.
In contrast, when released at the apical side of hepatocytes, the q-HEV particles will travel through the biliary conduct, and the bile salts will dissolve the quasimembranes, resulting in the excretion of naked virions (n-HEV) in the stool (Fig. 1C). Both naked and quasienveloped virions are infectious, but it is accepted that quasienveloped virions have less efficient and slower cellular uptake than naked virions (Yin et al., 2016). The receptor for n-HEV is still unknown, but several host factors, including heparan sulfate proteoglycans, glucose-regulated protein 78, asialoglycoprotein receptor, ATP synthase subunit 5β, and integrin α3, have been described to be involved in the cell attachment or entry of n-HEV.
In addition, studies using mass spectrometry have shown that 31 other proteins are involved in the entry process (Kalia et al., 2009). Apparently, the cell attachment of the quasienveloped virions is not as specific as that of the naked virions; this may be an explanation for the penetration of immunologically privileged sites.
Once the viral genetic material is released into the cell, the structural proteins are immediately translated. The complete process depends on the formation of the viral capsid, where ORF2 proteins package the genetic material of the virus to assemble new virions (Glitscher and Hildt, 2021; Kenney and Meng, 2019). Once the virus has been released from the infected cells through the exocytic GTPase Rab27-dependent pathway (Nagashima et al., 2014), diverse mechanisms allow viruses to evade the immune response, including the quasienvelope to enable the virus to avoid neutralization by anti-HEV antibodies (Fig. 2). This mechanism may allow virus replication, resulting in systemic manifestations.
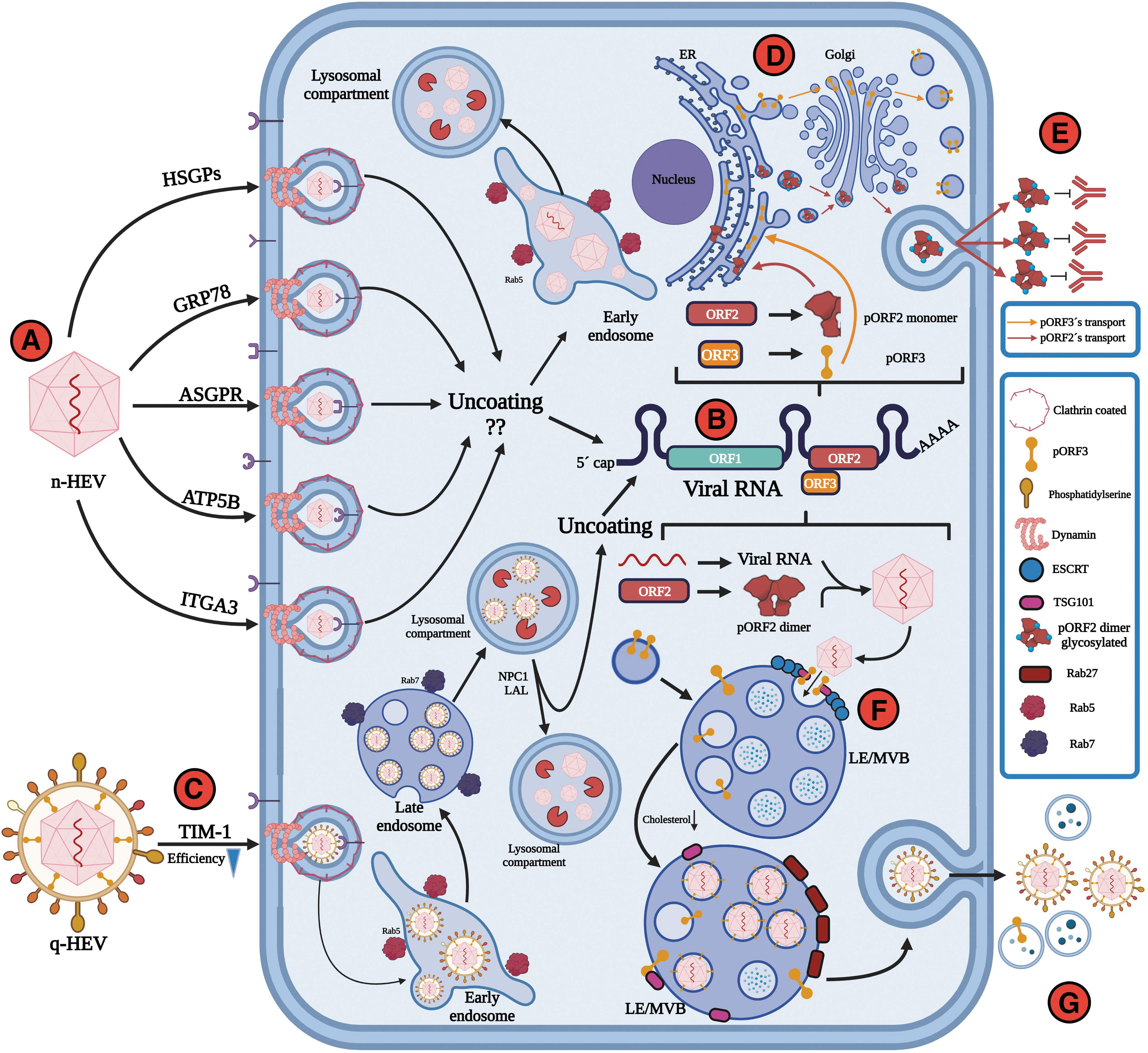
HEV entry and release of naked versus quasienveloped virus.
The Immunological Mechanisms of HEV-Associated Liver Disease
As mentioned before, viral genotypes are crucial in the course of infection, and the host immune response is also important and has been mainly studied in the setting of liver injury. Several lymphoid cells have been identified to contribute either to liver disease progression or to achieving sustained virologic response. From the histologic liver analyses of patients with acute HEV infection, neutrophils have been identified as the predominant population in inflammatory cell infiltrates (Peron et al., 2007). Moreover, studies with HEV-infected patients have shown a diminished presence of natural killer (NK) cells in the peripheral blood, whereas activation was strongly increased, indicating a possible migration to affected hepatic tissue, as shown by higher NK cell counts in liver biopsies (Prabhu et al., 2011; Srivastava et al., 2008). Importantly, the adaptive T cell response in HEV infection differs widely regardless of the course of infection.
An acute HEV infection is associated with elevated T cell frequencies. Diverse studies have shown an increased proportion of CD4+ and CD8+ T cell populations (Husain et al., 2011; Srivastava et al., 2007; Trehanpati et al., 2011). In this context, they are more activated and produce increased quantities of interferon-γ (IFN-γ) (Brown et al., 2016) as well as interleukin (IL)-10 (Wu et al., 2020b). Furthermore, an enhanced response and an elevated frequency of regulatory CD4+FoxP3+ T cells have been reported in acutely infected patients (Tripathy et al., 2012).
In patients with acute liver failure, an increased frequency of CD4+ T cells and Th2 cytokines, with a concomitant decrease in IFN-γ production, has been reported (Srivastava et al., 2011). It is important to consider that most of the reported findings come from peripheral lymphoid cells and do not necessarily reflect the ongoing situation in the infected liver. Nevertheless, few studies have examined postmortem liver biopsies of patients with liver failure due to HEV infection. These studies reveal a predominant CD8+ T cell-infiltrating population in the absence of CD4+FoxP3+ regulatory T cells (Agrawal et al., 2012; Prabhu et al., 2011). In contrast, in chronically HEV-infected patients, attenuated CD4+ and CD8+ T cell responses and decreased lymphocyte counts (Kamar et al., 2008) have been reported, and these changes are normalized after viral clearance (Suneetha et al., 2012).
Taken together, these results suggest a balanced regulation by pro- and anti-inflammatory cytokines in uncomplicated HEV infection and an exhausted T cell phenotype related to chronic disease (Fig. 3). However, how all these immune components underlie the development of extrahepatic manifestations associated with the virus has not been determined.
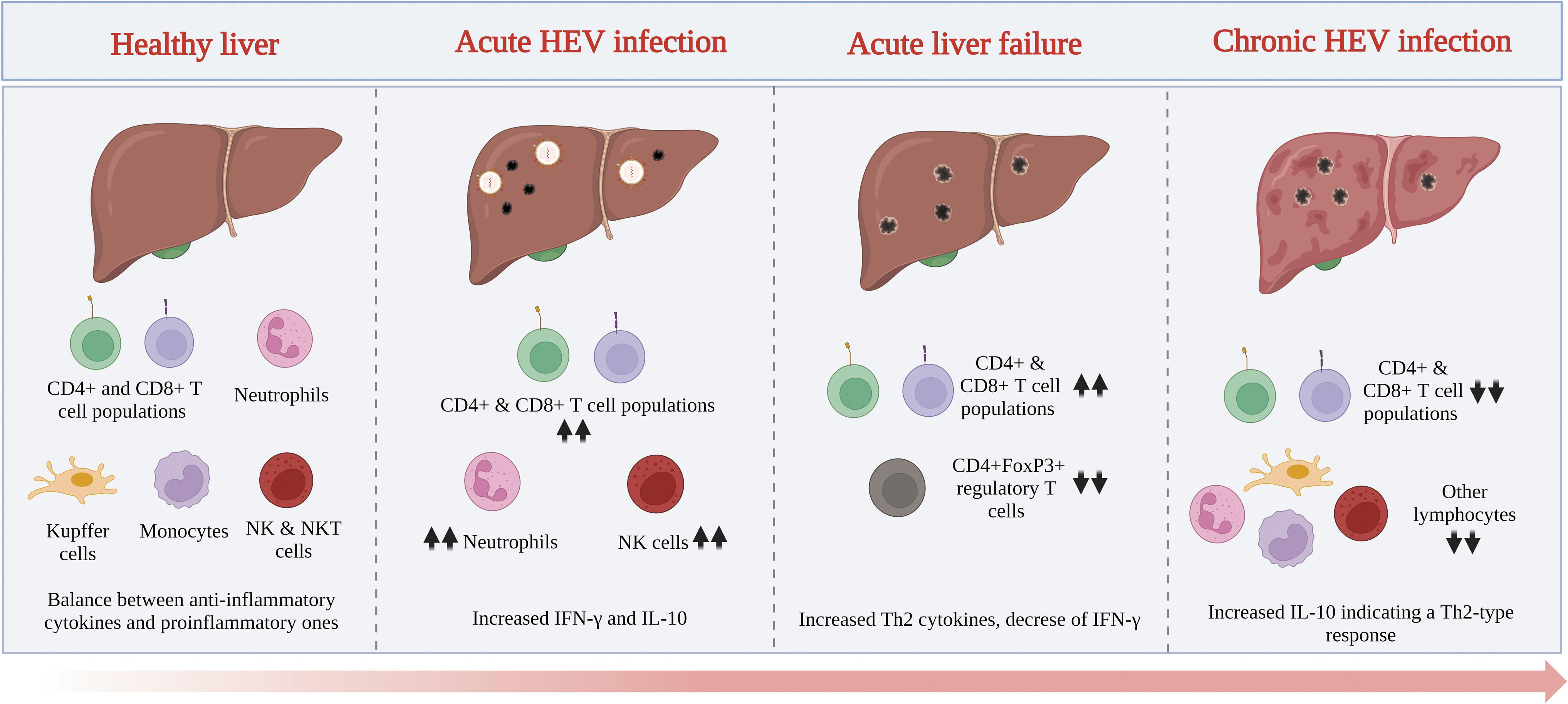
Generalities of the immune response in HEV-associated liver disease.
Importantly, in recent years, the relationship between gut dysbiosis and liver disease has become an important research topic. There is evidence of some degree of gut microbiota disorder in the occurrence and development of liver disease (Albhaisi et al., 2020; Chopyk and Grakoui, 2020; Lv et al., 2021; Rao et al., 2020; Wang et al., 2021), and viral hepatitis is linked with gut dysbiosis, particularly hepatitis C and B virus infections (Milosevic et al., 2021). Although there is scarce information regarding HEV infection, a study has shown that Proteobacteria, Gammaproteobacteria, and Enterobacteriaceae are abundant in acute HEV gt4-infected patients. In addition, a positive correlation between an increase in Gammaproteobacteria and serum alanine transaminase and total bilirubin levels in these patients has been reported (Wu et al., 2020c).
These findings are remarkable not only because the gut microbiota influences the immune response (Pickard et al., 2017) but also because the state of gut dysbiosis in several immune-privileged sites (Cook and Mansuy-Aubert, 2022; Liu et al., 2021; Santacroce et al., 2022) is particularly involved in HEV infection, as reviewed below. In addition, gut dysbiosis induces pathological manifestations (Chen et al., 2020; Rao et al., 2020), such as those associated with HEV.
HEV Infects Multiple Cells
Despite hepatocytes, several groups of cells are prone to be infected by HEV; as first described, information related to the life cycle points to the fact that viral particles infect the host through the epithelial cells of the intestines, where early replication begins. This has been demonstrated in swine models through immunohistochemical experiments in which the negative-sense RNA of the virus has been detected in the gastrointestinal tract, indicating viral replication (Williams et al., 2001). Indeed, a study with Caco2 cells (a human intestinal cell line) demonstrated that the ORF3 protein accumulates on the cell surface, which is evidence that virions are released through the apical surface of the membrane into the portal vein, targeting the liver (Emerson et al., 2010). This has recently been confirmed through the infection of primary intestinal cells and polarized enterocytes with gt1 and gt3, resulting in virus replication inside enterocytes and the intestinal mucosa.
Moreover, confocal microscopy studies have revealed that pORF2 in infected intestinal cells colocalizes with the trans-Golgi, a network involved in vesicle trafficking and HEV release, confirming that HEV can infect the intestines before reaching the liver. Furthermore, ribavirin does not completely inhibit the replication of the virus in enterocytes; it blocks the release of HEV virions at the basolateral side but not at the apical side. These data suggest that an HEV intestinal reservoir could contribute to HEV persistence under ribavirin therapy, thus contributing to the spread of the virus (Marion et al., 2020) and possibly to extrahepatic manifestations.
Peripheral blood mononuclear cells (PBMCs) are another group of HEV-infected cells. Evidence obtained from human PBMCs isolated during the acute phase of HEV infection confirms these data. Although viral positive-sense RNA was detected in most of the samples, negative-sense RNA was also detected in some samples, supporting virus replication, which is consistent with the detection of pORF2 inside PBMCs from these patients (Sayed et al., 2021a). Moreover, HEV replication has been reported in vitro in primary human-derived monocytes and macrophages isolated from healthy volunteers infected with gt1 and gt3 (Sayed et al., 2020).
HEV infection and replication, particularly with gt3 in neuronal-derived cell lines, may explain, in part, the neurological manifestations observed in HEV-infected patients. In fact, the oligodendrocyte cell line M03.13 shows viral RNA replication levels similar to hepatic infected cells; these cells are important for the development of myelin and are related to the Guillain‒Barré syndrome, which is commonly associated with HEV infection (Drave et al., 2016).
New evidence supports that gt1 and gt3 can replicate in primary human endometrial stromal cells (PHESCs) from healthy nonpregnant women, confirming a site of extrahepatic replication and PHESCs as possible viral reservoirs for vertical transmission during pregnancy (El-Mokhtar et al., 2020a). The placenta is another important tissue during vertical transmission, and findings from placental-derived cell lines (JEG-3) support the replication and a complete viral life cycle of gt1 and gt3 with an efficiency similar to that of human liver cells (Ratho et al., 2022). Other cell types in the kidneys, heart, pancreas, and reproductive organ tissues are also prone to HEV infection (Fig. 4).
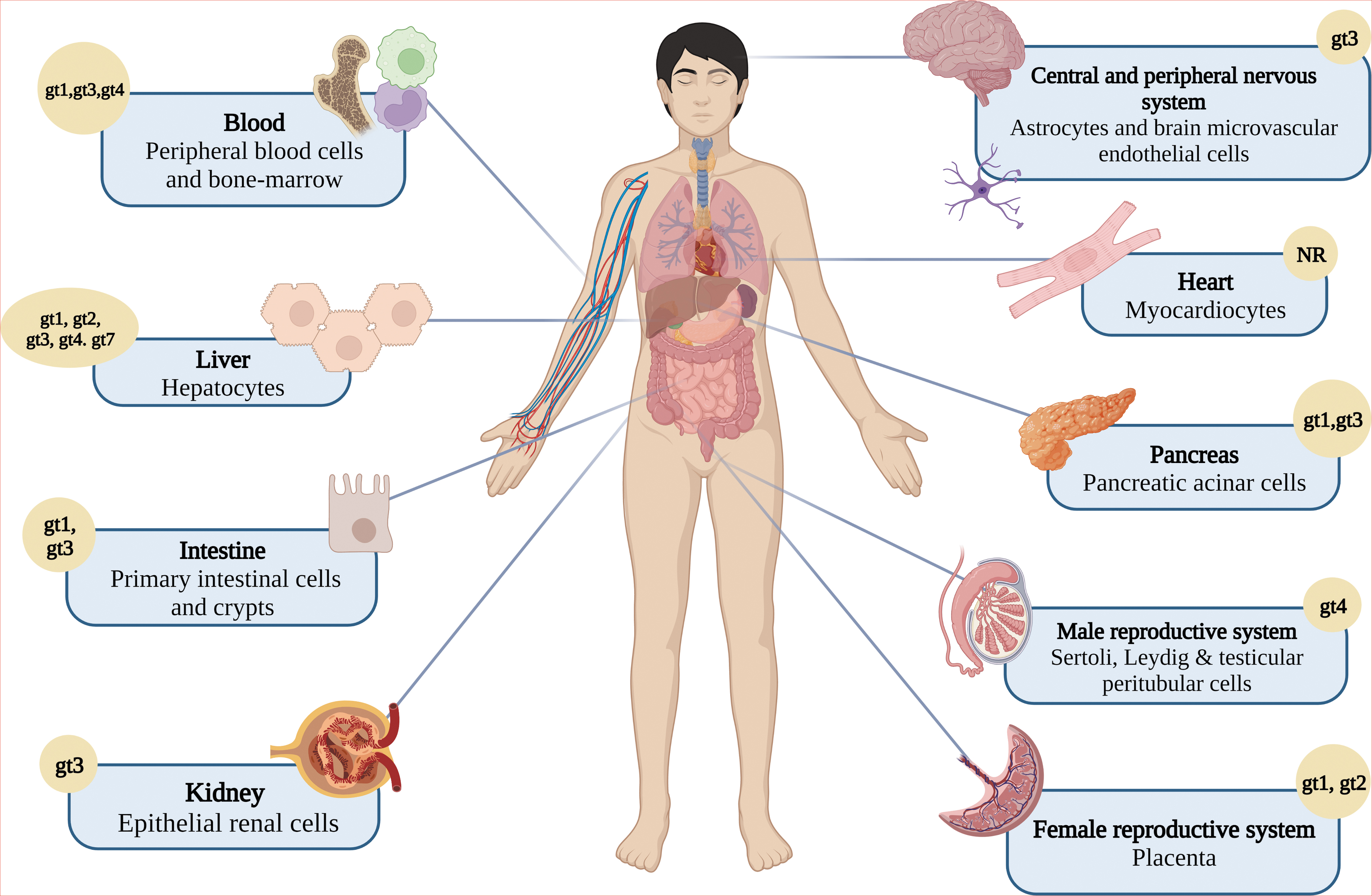
HEV affects multiple organs and systems. Beyond the liver, HEV infection results in extrahepatic manifestations related to the infection of diverse cell types. gt, genotype. Created with BioRender.com
HEV: A Systemic Disease?
The broad distribution of HEV in distinct tissues supports the notion that infection might be considered a systemic disease. These manifestations have mainly been reported in Europe and Asia, as illustrated in Table 1 (Abravanel et al., 2018; Colson et al., 2008; Dalton et al., 2017; Del Bello et al., 2015; Deroux et al., 2014; Fourquet et al., 2010; Fraga et al., 2018; Fukae et al., 2016; Jha et al., 2012; Kamar et al., 2012; Kamar et al., 2011; Kamar et al., 2005; Marion et al., 2018; Masood et al., 2014; Mengel et al., 2016; Montpellier et al., 2018; Perrin et al., 2015; Rehman et al., 2022; Ripellino et al., 2020; Sarkar et al., 2015; Singh and Gangappa, 2007; Sood, 2000; Stevens et al., 2017; Taton et al., 2013; Thapa et al., 2009; van den Berg et al., 2014; Verschuuren et al., 1997; Vikrant and Kumar, 2013; Wallace et al., 2020; Woolson et al., 2014; Yazaki et al., 2015).
Extrahepatic Manifestations in Hepatitis E Virus
In six patients.
In about 1/3 of the patients.
Available for 32.
Seven cases of NR.
ALT, alanine aminotransferase; GBS, Guillain-Barré syndrome; gt, genotype; MFS, Miller Fisher syndrome; NR, not reported; PTS, Parsonage–Turner syndrome or asymmetric neuralgic amyotrophy also known as brachial neuritis; U.K., the United Kingdom.
Patterns of infection have also been identified, such as the presentation of these extrahepatic manifestations in both immunocompetent patients and immunosuppressed patients, in patients with chronic infection, and in patients with acute infection (Jha et al., 2021). Moreover, the severity of extrahepatic manifestations does not necessarily correlate with the severity of HEV infection, and even asymptomatic HEV infection may trigger and cause systemic disorders. Genotypes 1, 3, and 4 have been associated with extrahepatic manifestations, with gt3 being the most frequently found (Kamar and Pischke, 2019; Wu et al., 2021).
According to a systemic review (Rawla et al., 2020), neurological manifestations are the most common (reported in 55% of a total of 324 articles), followed by hematological manifestations (35%); however, renal, endocrine, gastrointestinal, muscular, autoimmune, and male reproductive system manifestations have also been reported (Wu et al., 2021).
Neurological complications
Neurological complications are currently the most reported extrahepatic manifestations with an extensive relationship with gt3. The most common neurological complication is the Guillain‒Barré syndrome, with a prevalence between 5% and 11% in patients with acute HEV infection, followed by neuralgic amyotrophy, with a prevalence of 10.6% in patients with acute infection; both Guillain‒Barré syndrome and neuralgic amyotrophy directly impact the nerve roots and plexus (Jha et al., 2021). In several reports, patients with neurological complications present increased hepatic enzyme levels, which suggests previous or ongoing liver involvement; however, there are isolated cases of asymptomatic patients or even patients without liver enzyme alterations in laboratory tests. Neurological manifestations have been found to be significantly more common in immunocompetent patients, suggesting that a stable immune system is needed to generate a response to the virus, but at the same time tissue injury (Lhomme et al., 2021).
Moreover, neurotropism by the virus could take place in the compartmentalization of HEV quasienveloped species in the cerebrospinal fluid, which arises in a chronic infection, as it seems that the virus requires prolonged replication in the brain to obtain neurotropic variants, giving rise to lesions in the tissue (Abravanel et al., 2021).
Despite the absence of physiopathogenic mechanisms to explain the Guillain‒Barré syndrome in HEV infection, a relationship between axonal degeneration caused by the virus or the host response, which would give rise to the syndrome, characterized as a demyelinating disease, has been suggested; this coincides with experiments performed in a model of rabbits infected by HEV, where axonal degeneration was observed in the white matter of the spinal cord, as well as necrosis in axons, Purkinje cells, neuronophagia, nodules in microglia, and hemorrhages in the dorsal area, which could contribute to other neurological disorders associated with the infection (Tian et al., 2019).
Renal complications
HEV seroprevalence has increased significantly in recent years in hemodialyzed patients, which is important due to the identification of more significant extrahepatic manifestations in the kidneys of HEV-infected patients than in those of hepatitis A virus (HAV)-infected patients (Brehm et al., 2021). A relationship between the generation of glomerulonephritis (GN) and HEV infection, especially in patients with cryoglobulinemia, has been documented (Guinault et al., 2016; Pischke et al., 2014). Indeed, HEV is a nephrotoxic agent, generating cases of GN in greater numbers than HAV-infected individuals. There is also a significant decrease in the glomerular filtration rate in liver and kidney transplant recipients during the acute and chronic phases of HEV infection (Fousekis et al., 2020; Pischke et al., 2017).
Most renal complications are commonly linked to HEV gt3, except for isolated cases of thrombocytopenia and GN in patients with acute HEV gt1 infection, in whom the infection corrupts amino acid metabolism, thus increasing
Moreover, although no evidence of an association between GN and HEV gt3 previous exposure has been reported (Pischke et al., 2022), an augmentation of bilirubin and AST levels in anti-HEV IgG patients when compared with HEV-seropositive healthy individuals has been recently reported in a region where HEV gt1 prevails (El-Mokhtar et al., 2023); this supports the notion of a link between both conditions and underscores a potential role of genotypes in the development of HEV-related GN.
Hematological complications
Anemia, hematopoietic syndrome, pure red blood cell hypoplasia, and monoclonal gammopathy of undetermined significance are the most common hematological disorders related to HEV infection, but in patients infected with HEV gt1, gt3, and gt4, thrombocytopenia and cryoglobulinemia are the most reported hematological complications (Wu et al., 2021). The relationship of these manifestations to infection by the virus is increasingly strong due to evidence of virus replication in bone marrow and human PBMCs, which may contribute to infection recurrence, chronicity, and especially distribution to extrahepatic regions; in addition, blood products have been identified as important sources of the transmission of HEV infection (Sayed et al., 2020; Sayed et al., 2021a).
Gastrointestinal complications
Acute pancreatitis is the most reported gastrointestinal manifestation of HEV infection, and according to a systematic review, 18% of HEV patients develop severe pancreatitis (Haffar et al., 2015). Pancreatitis is related to HEV gt1 and gt3, and although the underlying mechanisms are unknown, it is proposed that infection induces direct inflammation and destruction of pancreatic acinar cells, as well as the release of lysosomal enzymes from inflamed hepatocytes through the portal vein and, in conjunction with the action of trypsin, which is obtained by the activation of trypsinogen into the circulation, causes pancreatic damage that progresses to acute pancreatitis (Sudhamshu et al., 2011). Intrapancreatic hemorrhage or disseminated intravascular coagulation are manifestations rarely observed (Rawla et al., 2020; Wu et al., 2021).
Other autoimmune manifestations
There are isolated reports of miscellaneous disorders, such as autoimmune thyroiditis, pleural effusion, acute graft dysfunction, acute polyarthritis, myocarditis (Premkumar et al., 2015), thyrotoxicosis (Hui et al., 2003), Henoch–Schönlein purpura (Thapa et al., 2010), cutaneous necrotizing small-vessel vasculitis and cutaneous T cell lymphoproliferative disorder (Mallet et al., 2017), but there are still limitations in the hypotheses proposed and the findings observed in HEV-infected patients, and so, as with the other manifestations, more in-depth research is needed (Fousekis et al., 2020; Rawla et al., 2020).
Immune Privilege and Its Relation to Extrahepatic Manifestations
Diverse study findings have emerged to explain HEV extrahepatic manifestations. The direct effects of virus replication in peripheral tissues represent plausible explanations (Kupke and Werner, 2021; Velavan et al., 2021). In addition, considering that the immune response is crucial in the development of the disease, the commonalities of immune characteristics of affected tissues may support the HEV effects.
In the 1940s, the biologist Peter Medawar first described the term immune privilege, referring to the absence of immune responses to minimize the likelihood of inflammation and systemic mechanisms, particularly at sites such as the brain, testicles, placenta, and eyes, which have sophisticated anatomical barriers (Male, 2021). This has become relevant due to the ability of HEV to penetrate these barriers, specifically the blood–brain barrier (BBB), hematotesticular barrier, and placenta.
BBB and HEV
The BBB, which consists of an epithelial layer with tight junctions, impedes the entry of immune cells and inflammatory mediators into the brain; in addition, the scarcity of dendritic cells in the brain reduces adaptive immunity to antigens (Abbas et al., 2015). This becomes relevant due to the numerous conditions mentioned above, such as infection by a virus that can cause dysfunctional processing in the central nervous system through the activation of limited barrier responses that under conditions of inflammation generate changes in the neurovascular unit and that compromise the immune-privileged status of the tissue (Muldoon et al., 2013).
It has been demonstrated, through an in vitro experimental model of the BBB, that HEV is able to enter the barrier in any of its conformations (nonenveloped and quasienveloped), which could be an essential finding in the search for answers about HEV infection. The potential mechanism of BBB invasion by HEV was demonstrated by inoculating human brain endothelial cells, which cover the barrier in vivo, and human astrocytes, causing a productive viral infection directly through these cells. Likewise, HEV has been found to produce mild histological changes in some tissues, such as meningitis, gliosis, and perivascular inflammation, and to induce high levels of proinflammatory cytokines (tumor necrosis factor-α, IL-18) in an animal model (Tian et al., 2022).
Blood–testicle barrier and HEV
In direct functional analogy of the central nervous system and BBB, the testicles pose a sophisticated structure barrier, the blood–testicle barrier (BTB), to safeguard the contents of the testicles and the process of spermatogenesis, including the Sertoli cells that line the outer layer of the seminiferous tubules, as well as the androgen-rich environment with its inflammatory influence on macrophages. The BTB is composed of Leydig cells (LCs), Sertoli cells, and testicular peritubular cells (TPCs), which produce transforming growth factor-β that contributes to limiting the immune response in the region in the case of proinflammatory situations. It should be noted that this type of involvement related to viral infection is still being studied because despite the finding of HEV infection in infertile men and murine studies on testicular involvement, there are groups of researchers who deny a relationship.
The controversy is based on the variability of the results obtained by distinct groups. Little or no detection of RNA-HEV was found in semen from HEV gt1-infected patients who presented with one or more symptoms of acute hepatitis, and no significant differences in reproductive hormone levels (follicle stimulating hormone, luteinizing hormone) were observed. Similarly, no significant differences were found in semen quality or quantity (El-Mokhtar et al., 2021). Moreover, no HEV RNA was detected in 79 semen samples from infertile men in a European population (Horvatits et al., 2020).
On the contrary, the presence of HEV antigens in BALB/c SPF mice, specifically in LCs and TPCs, has been demonstrated, which reaffirms the replication of the virus in the early stages of infection and the alteration of sperm quality (necrozoospermia); in addition, this finding confirms the capacity of the virus to cross the BTB, altering the underlying mechanisms for its preservation, which is probably the reason the virus is attracted to this tissue, that is, the depleted immune responses (Situ et al., 2020). Furthermore, the notion of the male reproductive system as a niche of HEV persistence has been supported by using an experimental HEV gt3-infected pig model where HEV genetic variants in the semen were distinct when compared with the variants in the stool and serum (Horvatits et al., 2021).
Therefore, damage to the testicular barrier cannot yet be ruled out due to the limitation of study models and variables to be considered, such as genetic variants and their infective capacity and host factors (immunocompetence vs. immunosuppression).
Placental tissue: impact of HEV infection during pregnancy
The placenta is an immune privileged organ where an important relationship with HEV infection is observed; viral in situ replication has been recently found, and the relationship of hepatitis E with high mortality in pregnant women (5–22%) has been documented (Bigna et al., 2020; Bose et al., 2014; Ratho et al., 2022). During pregnancy, there is a meticulous balance to maintain strong defenses against microorganisms that can infect the mother and the fetus, as well as a tolerance of the semiallogenic fetus. This is possible due to the suppression of responses to fetal antigens and a meticulously selected immune microenvironment composed mainly of maternal NK cells.
According to a comparison study between pregnant women with HEV-induced fulminant hepatic failure and nonfulminant and healthy pregnant women as controls, it was found that in women with fulminant liver disease, lymphocytes, dendritic cells, and macrophages have decreased phagocytic capacity and low TLR3 and TLR9, which prevents IFN production (Sehgal et al., 2015).
Despite the unknowns surrounding this virus and the candidate mechanisms for the high susceptibility to adverse outcomes due to infection during pregnancy, there is research suggesting an interaction between estrogen and progesterone receptor depletion, as well as the maintenance of the antigenic fetus in the mother through the suppression of T cell-mediated immunity, in HEV infection with high viral load. These same hormonal and immunological factors favor harmful events for the fetus and the mother, such as low birth weight, maternal death, and prematurity <36 weeks (Bigna et al., 2020).
It has been demonstrated that HEV gt1 has significantly higher replication than gt3 in the maternal decidua, placenta, and placental stromal cells, and such efficacy is thought to be the main cause of HEV-associated complications mentioned above; in contrast, gt3 has a mild-to-moderate and self-limited course (El-Mokhtar et al., 2020a). Finally, vertical transmission from mother to fetus could be justified by direct viral infection of stromal cells derived from the decidua and placenta, which has been previously demonstrated, meaning that the placenta could be a reservoir and an endogenous source of transmission (El-Mokhtar et al., 2020a; Wu et al., 2020a).
Remarks
Using membrane hijacking endows HEV with the possibility of dissemination inside and outside of its host; this is evidenced by its effect on health beyond the consequences on the liver initially associated with this virus. Likewise, due to the immune-privileged status of specific tissues, HEV can replicate in diverse organs and promote systemic disorders. Therefore, in addition to viral genotypes and the different routes of transmission, the characteristics of the immunological environment of affected tissues could be a landmark in the development of extrahepatic manifestations (Fig. 5).
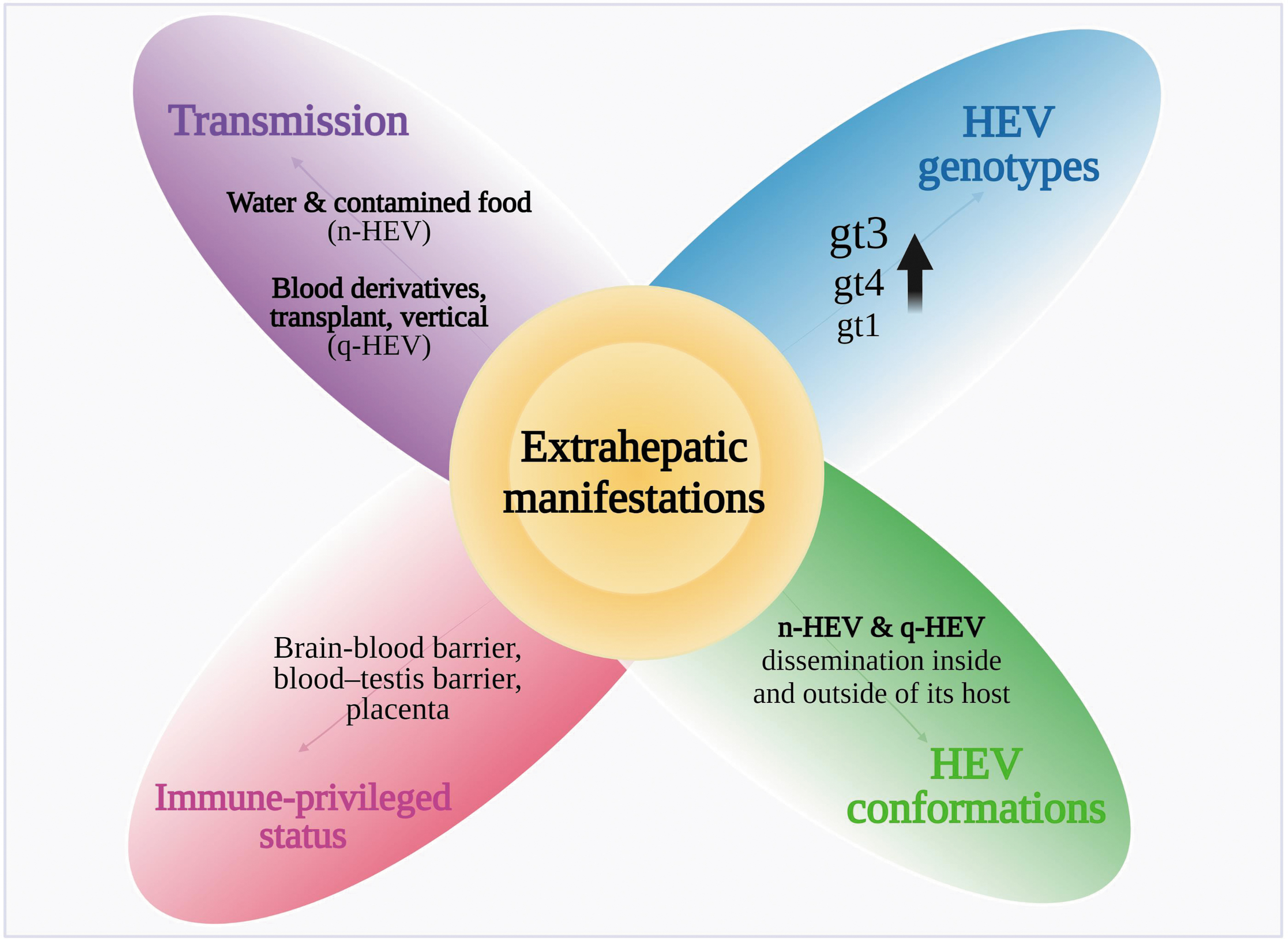
Key elements from virus biology and host response implicated in the development of extrahepatic manifestations. Created with BioRender.com
Hepatitis E is not part of the differential diagnosis associated with the wide diversity of extrahepatic manifestations related to the virus. Therefore, the existing tools used for HEV diagnosis in the setting of liver disease, including serologic tests, HEV RNA in feces and serum samples (Zhao and Wang, 2023) and the detection of viral antigens in urine (Stahl et al., 2023), should be implemented during the development of extrahepatic symptoms. Furthermore, the search for specific biomarkers associated with the infection, including the study of immune components (cytokines, for example), ideally in noninvasive tests, that allows the delineation of infectious status is needed in this scenario.
Similarly, we currently have in vivo (Liu et al., 2023; Sayed et al., 2019) and in vitro (Pellerin et al., 2021; Schemmerer et al., 2016) models for the study of the pathogenesis associated with HEV infection. These models have been dedicated mainly to the study of hepatic alterations, while their use during extrahepatic symptoms is still limited. Recent advances in the development of in vivo models to study the congenital transmission of the virus (Yadav and Kenney, 2023) will be useful to identify the immune elements related to the penetration and persistence of HEV in the placenta. In addition, in vivo models have allowed progress in the study of neurological alterations. Improving these models, which should recapitulate hepatitis E pathology and, importantly, sites of infection, and adapting cell culture models for specific extrahepatic manifestations are needed to advance the description of the mechanisms associated with these events.
There is no approved drug for the treatment of HEV infection. Ribavirin and IFN-α treatment is recommended in patients with chronic HEV, and ritonavir in conjunction with ribavirin has been shown to block virus internalization and promote HEV clearing in vitro (Primadharsini et al., 2020). However, the benefit of this management in the setting of extrahepatic manifestations has not been demonstrated.
Understanding the immunopathogenesis and the mechanisms of HEV propagation to diverse tissues will provide clues for the appropriate design of diagnosis and therapies aimed at fighting this infectious agent.
Footnotes
Authors' Contributions
J.O.-C.: writing—original draft and writing—review and editing; C.M.: writing—original draft and writing—review and editing; G.D.P.: conceptualization; E.L.: writing—review and editing; G.C.: writing—review and editing; N.A.F.: conceptualization, writing—original draft, supervision, funding acquisition, and writing—review and editing.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by grant IA201422 from Programa de Apoyo a Proyectos de Investigación e Innovación Tecnológica (PAPIIT), Dirección de Asuntos del Personal Académico (DGAPA), and Universidad Nacional Autónoma de México (UNAM), Mexico.