
Abstract
Hantaan virus (HTNV) is prevalent in Eurasia. It causes hemorrhagic fever with renal syndrome (HFRS). Long noncoding RNAs (lncRNAs) play key roles in regulating innate immunity. Among these, lncRNA negative regulator of interferon response (NRIR) was reported as an inhibitor of several interferon (IFN)-stimulated genes. Our results showed that: NRIR expression was upregulated by HTNV infection in a type I IFN-dependent manner. The expression of NRIR in CD14+ monocytes from HFRS patients in acute phase was significantly higher than that in convalescent phase and healthy controls. HTNV infection in some HTNV-compatible cells was promoted by NRIR. NRIR negatively regulated innate immunity, especially IFITM3 expression. Localized in the nucleus, NRIR bound with HNRNPC, and knockdown of HNRNPC significantly weakened the effect of NRIR in promoting HTNV infection and restored IFITM3 expression. These results indicated that NRIR regulates the innate immune response against HTNV infection possibly through its interaction with HNRNPC and its influence on IFITM3.
Introduction
Hantavirus, currently classified as belonging to the order Bunyavirales, is an enveloped negative-stranded RNA virus maintained and transmitted mainly through rodents (Kell et al., 2020). Hantavirus causes both hemorrhagic fevers with renal syndrome (HFRS) and hantavirus cardiopulmonary syndrome. HFRS, which is caused by the “Old World” hantaviruses, including Hantaan virus (HTNV), Seoul virus, Dobrava virus, and Puumala virus, is widely prevalent in Eurasia (Jiang et al., 2016). China has long been a high-prevalence area for HFRS (Yang et al., 2022). To date, no specific treatment has been developed for HFRS, and our understanding of HTNV–host interaction remains limited.
HTNV infects many types of host cells, among which endothelial cells and monocytes are considered to play essential roles in the pathogenesis of HFRS (Li et al., 2021; Lu et al., 2020; Matthys and Mackow, 2012; Raftery et al., 2020). Pattern recognition receptors (PRRs), including TLR3, TLR4, and RIG-I, trigger innate immune response against HTNV infection (Handke et al., 2009; Kell et al., 2020; Yu et al., 2012). Activated monocytes and macrophages cause cytokine storm, which in turn leads to vascular leakage and other manifestations. The induction of type I interferon (IFN) by HTNV and the subsequent expression of interferon-stimulated genes (ISGs), such as IFITM3, have been reported to regulate HTNV infection (Kim et al., 2007; Xu-Yang et al., 2016). Hantavirus also has ways to escape from innate immunity (Zhang et al., 2021).
Long noncoding RNAs (lncRNAs) are defined as RNAs longer than 200 nucleotides with no protein-coding capability. Many lncRNAs are closely related to innate immunity, including the IFN response (Vierbuchen and Fitzgerald, 2021). Among these, lncRNA negative regulator of interferon response (NRIR) was characterized as an IFN-responsive transcript that negatively regulates several ISGs in HCV infection (Kambara et al., 2014). NRIR expression was also altered in COVID-19 patients (Enguita et al., 2022). The induction of CXCL10 and CXCL11 was reduced in NRIR-knockdown monocytes from patients with systemic sclerosis (Mariotti et al., 2019). Some NRIR-regulated ISGs, such as IFITM1, have anti-HTNV potential (Xu-Yang et al., 2016). But the role of NRIR in HTNV infection and its mechanism is still unclear.
Our study revealed the importance of NRIR in HTNV infection as a feedback inhibitor of IFN. NRIR could be induced by HTNV in a type I IFN-dependent manner and it promoted HTNV infection in different cell lines, while NRIR knockdown suppressed HTNV infection in human umbilical vein endothelial cells (HUVECs). NRIR suppressed the expression of anti-HTNV ISG IFITM3. Mechanistically, NRIR binds to HNRNPC, HNRNPC knockdown reduced the promoting effect of NRIR to HTNV infection and restored IFITM3 expression. We also found the changes of NRIR expression in patients' CD14+ monocytes with the progression of HFRS.
Materials and Methods
Ethics statement and clinical sample collection
This study was conducted following the ethical standards laid down in the Declaration of Helsinki and its later amendments. We also followed Chinese government regulations. The Institutional Ethics Committee of Tangdu Hospital approved this study. Before inclusion, all participants were informed of the study objectives and signed written informed consent. Ten milliliters of venous blood samples with EDTA anticoagulation were obtained from nine patients with HTNV-caused HFRS (in their respective acute and convalescent phases) as well as 11 healthy controls. Peripheral blood mononuclear cells were isolated by density gradient centrifugation using Lymphoprep (Stemcell Technologies, Vancouver, Canada). CD14+ monocytes were positively selected using human CD14 MicroBeads (Miltenyi Biotec, Bergisch Gladbach, Germany) following the manufacturer's protocols.
Cell lines and virus
HUVECs within passage 6 (ScienCell Research Laboratories) were cultured in endothelial cell medium (ScienCell). THP-1, Huh7.5, and Vero E6 cells were maintained in our laboratory (Li et al., 2021; Ma et al., 2017; Xu-Yang et al., 2016) and tested negative for mycoplasma. All cells were cultured at 37°C with 5% CO2. The HTNV 76–118 strain was maintained in our laboratory and propagated in Vero E6 cells. Cells were infected with HTNV at indicated multiplicities of infection (MOI). Anifrolumab (MedChemExpress, HY-P99168) was used to treat cells at 20 μg/mL with HTNV infection.
Cell transfection
NRIR expression plasmid, HTNV NP expression plasmid with c-myc tag and HNRNPC shRNA plasmid, as well as their relative controls were transfected using Effectene Transfection Reagent (Qiagen) following the manufacturer's protocol. After transfection, cells were incubated for 24 h before further analysis. siRNA transfections were performed using the HiPerFect Transfection Reagent (Qiagen) following the manufacturer's protocol. The final concentration of siRNA was 10 nM in HUVECs, 5 nM in Huh7.5 cells, and 100 nM in THP-1 cells. Commercialized siRNAs were used with undisclosed sequences (Supplementary Table S1). Due to the low basal expression of NRIR, we treated the cells with IFN-α2a (1,000 U/mL) for 6 h following siRNA transfection.
Quantitative real-time polymerase chain reaction
Total RNA was extracted using the Total RNA Kit I (Omega Bio-Tek; R6834), and cDNA was synthesized using the PrimeScript RT Reagent Kit (TaKaRa Biotechnology; RR047A). Quantitative real-time polymerase chain reaction (RT-qPCR) was performed using TB Green Premix Ex Taq II (TaKaRa Biotechnology; RR820A) on Roche LightCycler 480 system with β-actin as reference. Primer sequences are in Supplementary Table S2.
Western blotting
The cells were harvested using Radio Immunoprecipitation Assay buffer containing protease and phosphatase inhibitors (Sangon Biotech) on ice. The protein samples were quantified using the BCA Protein Assay Kit (Thermo Fisher Scientific; 23225). Equal amounts of proteins samples (20 μg/lane) were separated on SDS-polyacrylamide gels and transferred onto polyvinylidene fluoride membranes (Millipore, MA). After blocking with 5% bovine serum albumin, the membranes were incubated with primary antibodies (Supplementary Table S3) following the manufacturer's instructions. Horseradish peroxidase-conjugated secondary antibodies (Proteintech; SA00001-1/-2) were used and the blots were developed on Bio-Rad ChemiDoc MP (analyzed by ImageJ).
Fluorescence in situ hybridization
Fluorescence in situ hybridization (FISH) was performed using a FISH Kit and NRIR Probe Mix (C10910; RiboBio, Guangzhou, China) following the manufacturer's protocol (detailed in Supplementary Data) and observed on Olympus FV1000 confocal microscope.
Transcriptome sequencing
The transcriptome sequencing analysis was performed by Shanghai NovelBio Bio-Pharm Technology. Differentially expressed genes with fold change >1.5 or <0.667, p-value <0.05, and false discovery rate (FDR) <0.05 were subjected to Gene Ontology (GO) analysis and Kyoto Encyclopedia of Genes and Genomes (KEGG) pathway analysis. FPKM data were used in the heatmaps.
lncRNAs–protein interaction proteomics and RNA immunoprecipitation
Identification of NRIR-binding proteins by mass spectrometry (chromatin isolation by RNA purification-mass spectrometry [ChIRP-MS]) was performed and analyzed by KangChen Bio-tech, Shanghai, China using THP-1 cells. The propensity of the interactions between different fragments of NRIR (nt) and HNRNPC (aa) was predicted using catRAPID omics V2.1. The binding of NRIR to HNRNPC were verified by RNA immunoprecipitation (RNA IP) in THP-1 and HUVEC cells treated with 1,000 U/mL IFN-α2a using the RIP Kit (Merck-millipore; 17-700). See antibodies in Supplementary Table S1.
Statistical analysis
Data were analyzed using GraphPad Prism 7 software and showed as mean ± standard error of the mean. The two-tailed unpaired-samples t test was used to compare two groups. For multiple comparisons, one-way analysis of variance was performed. The difference was considered to be significant when the p values were <0.05 (*), <0.01 (**), or <0.001 (***).
Results
NRIR expression was induced by HTNV infection in a type I IFN-dependent manner
All three cells could be infected with HTNV (Fig. 1A–C, MOI = 1). A significant upregulation of NRIR expression was observed after HTNV infection in HUVECs and THP-1 (Fig. 1D). The fold changes were comparable to IFN-α2a stimulation (Fig. 1G). We did not find changes in NRIR expression in Huh7.5 cells (Fig. 1D). HTNV NP expression in HUVEC and THP-1 cells also induced NRIR expression to a less extent (Fig. 1E). The induction of NRIR by HTNV infection was blocked by Anifrolumab (a type I IFN receptor antagonist, Supplementary Fig. S1). NRIR expression in CD14+ monocytes from HFRS patients in acute phase was higher than that in healthy controls and patients in the convalescent phase (Fig. 1F). ISGs, including IFITM3, IFIT1, and MX1 were also upregulated by HTNV infection in HUVEC and THP-1 cells (Supplementary Fig. S2).
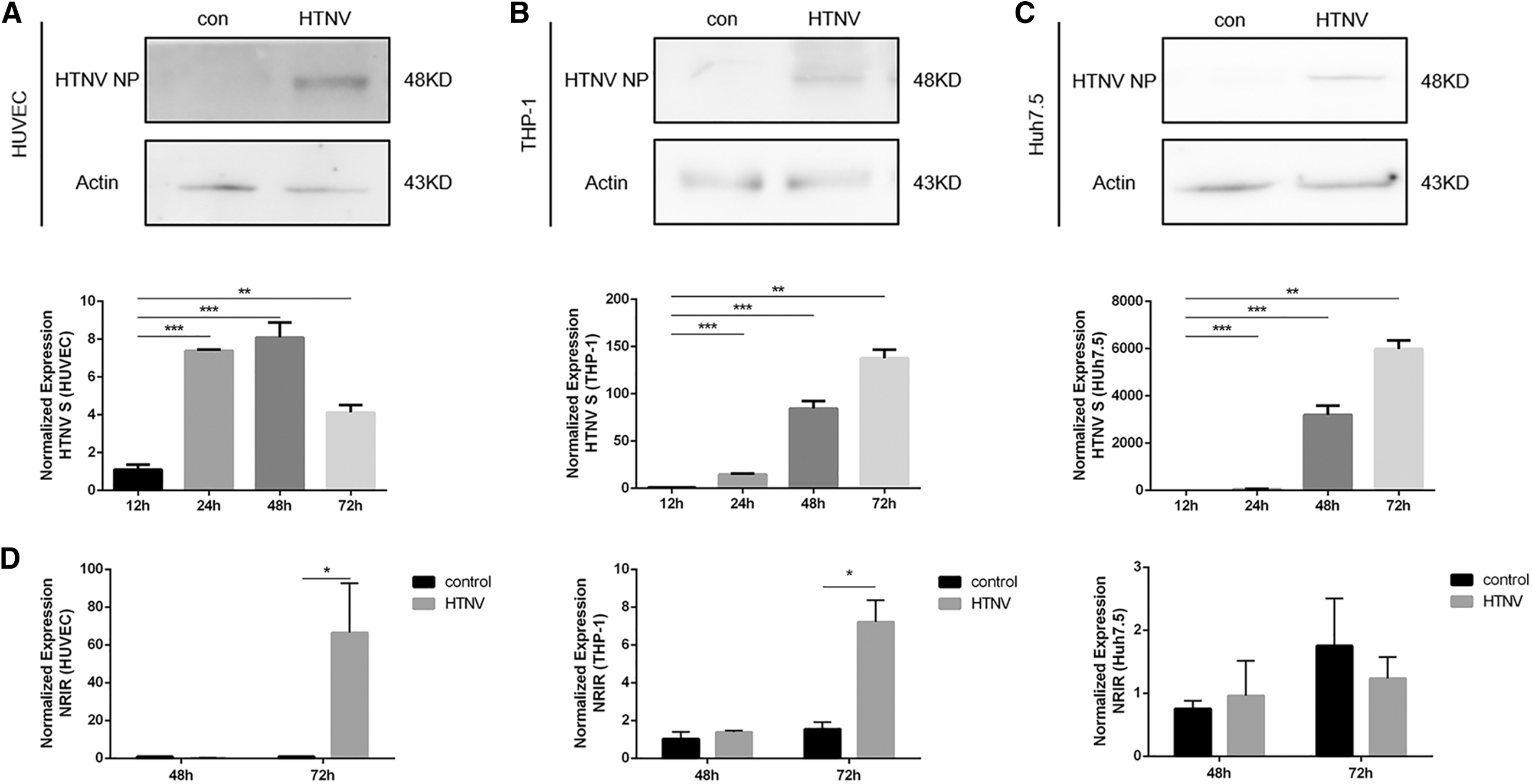
NRIR expression was induced by HTNV infection.

NRIR facilitated HTNV infection
For NRIR overexpression, we transfected pcDNA3.1-NRIR and control plasmids into HUVECs, THP-1, or Huh7.5 cells for 24 h (Fig. 2A) and challenged them with HTNV (MOI = 1). The overexpression of NRIR significantly promoted HTNV infection in all three cells (Fig. 2B and Supplementary Fig. S3A). In the knockdown experiments, the effects of four different siRNAs were tested in THP-1 cells, with siRNA4 selected as the most efficient and applied in HUVECs and Huh 7.5 cells (Fig. 2C, D). Three hours after siRNA transfection, cells were treated with IFN-α2a (1,000 U/mL) for 6 h and then challenged with HTNV (MOI = 1). We found HTNV infection was attenuated after NRIR knockdown only in HUVECs (Fig. 2E and Supplementary Fig. S3B).
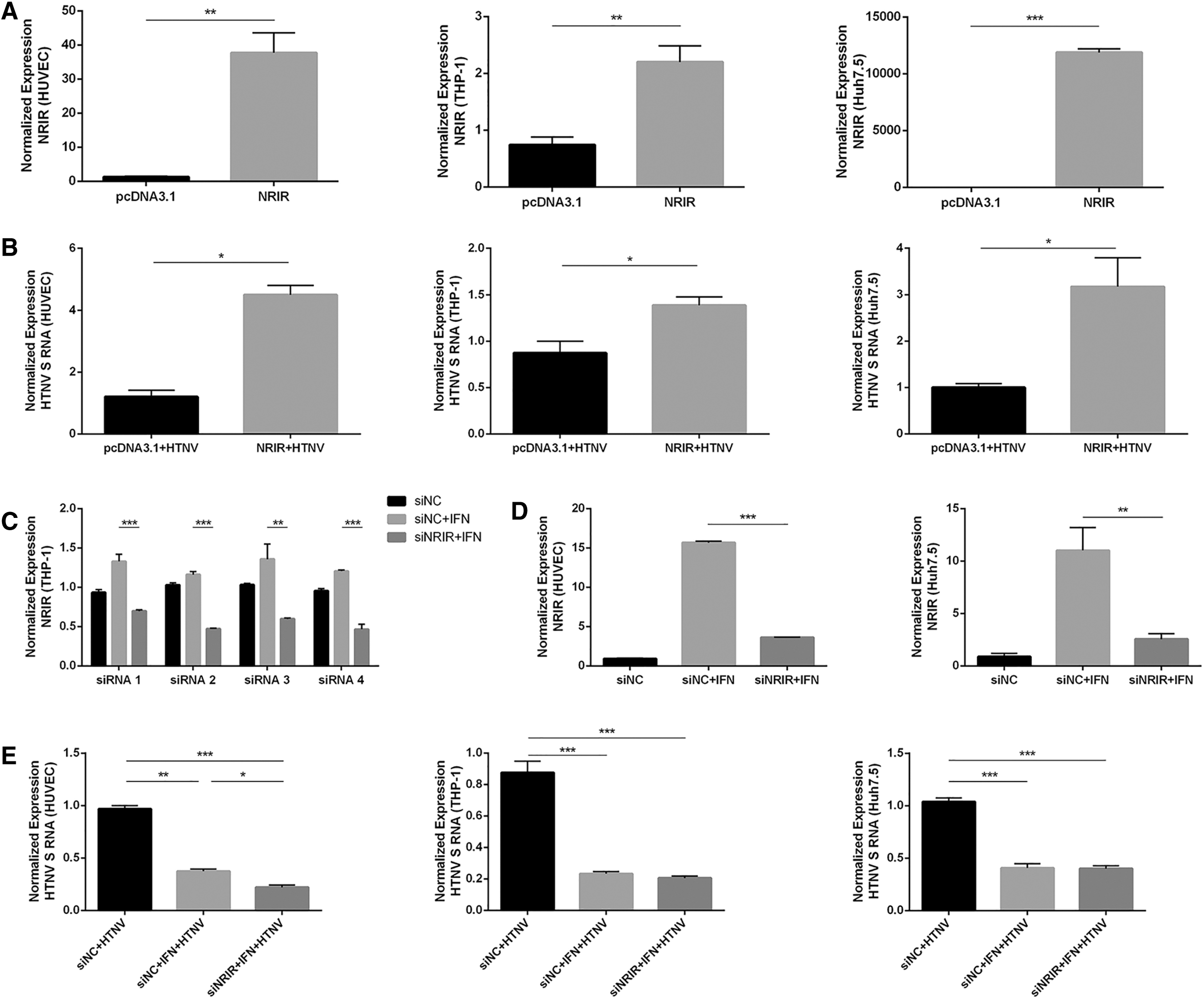
NRIR facilitated HTNV infection.
NRIR suppressed the expression of HTNV-related ISGs in HUVECs
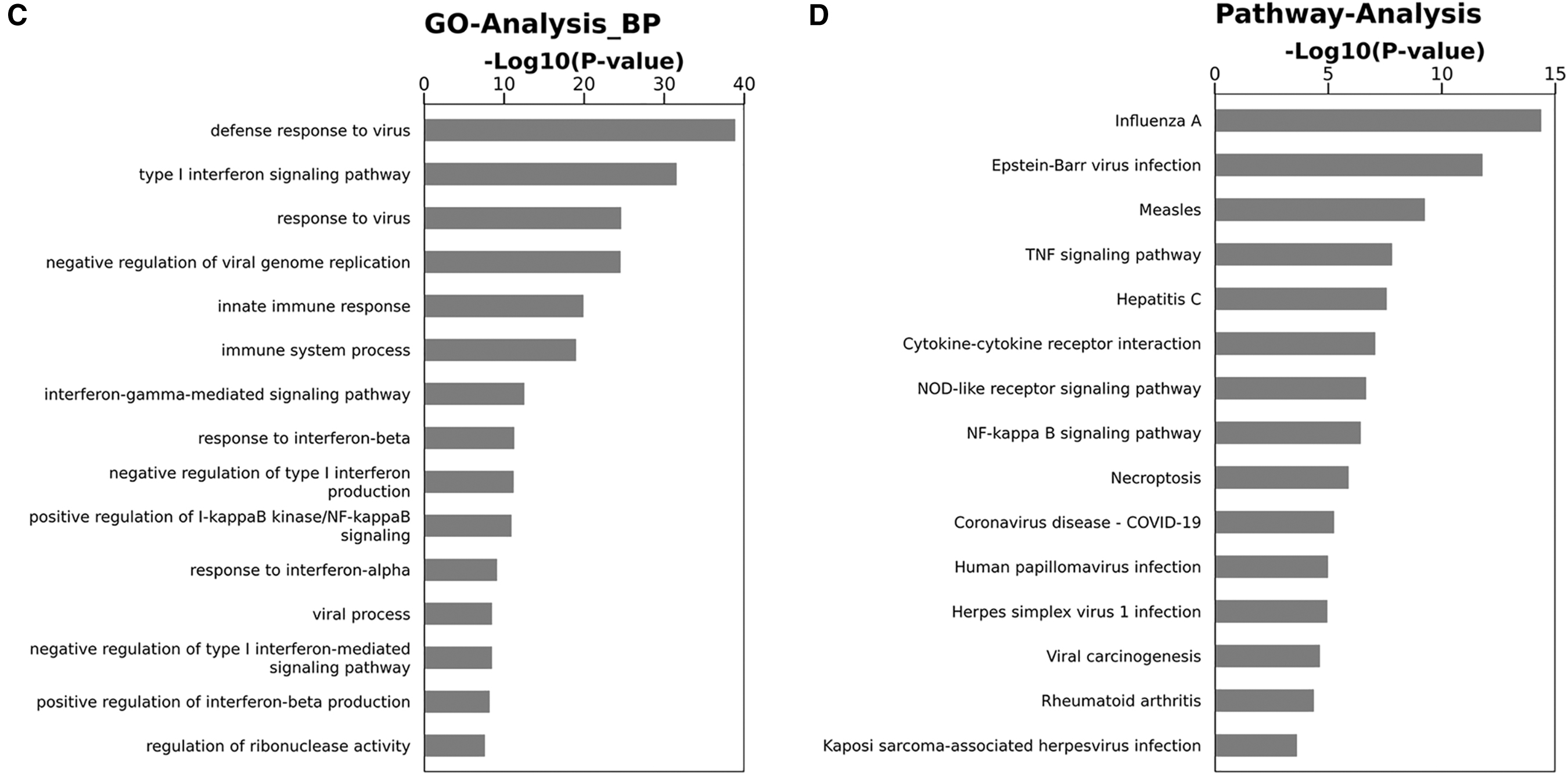
Transcriptome sequencing in HUVECs showed that 361 genes were downregulated after NRIR overexpression, including IFITM3, MX1, and many other innate immune molecules (Fig. 3A). Several ISGs, including IFITM3, IFITM1, IFIT1, ISG15, MX1, and OAS2 were tested using RT-qPCR (Fig. 3B and Supplementary Fig. S4A). The GO analysis showed that the downregulated genes were significantly enriched in GO terms “defense response to a virus,” “type I interferon signaling pathway,” and “response to a virus” (Fig. 3C). The KEGG analysis showed a considerable enrichment of downregulated genes in “influenza A” pathway (Fig. 3D). Importantly, the expression of IFITM3, which has anti-HTNV effect, were downregulated by NRIR overexpression in both basal and HTNV infection conditions in HUVECs (Fig. 3B). At the same time, IFITM3 expression was induced by IFN-α2a stimulation and was further increased when NRIR was knocked down in both basal and HTNV infection conditions in HUVECs (Fig. 3E and Supplementary Fig. S4B).
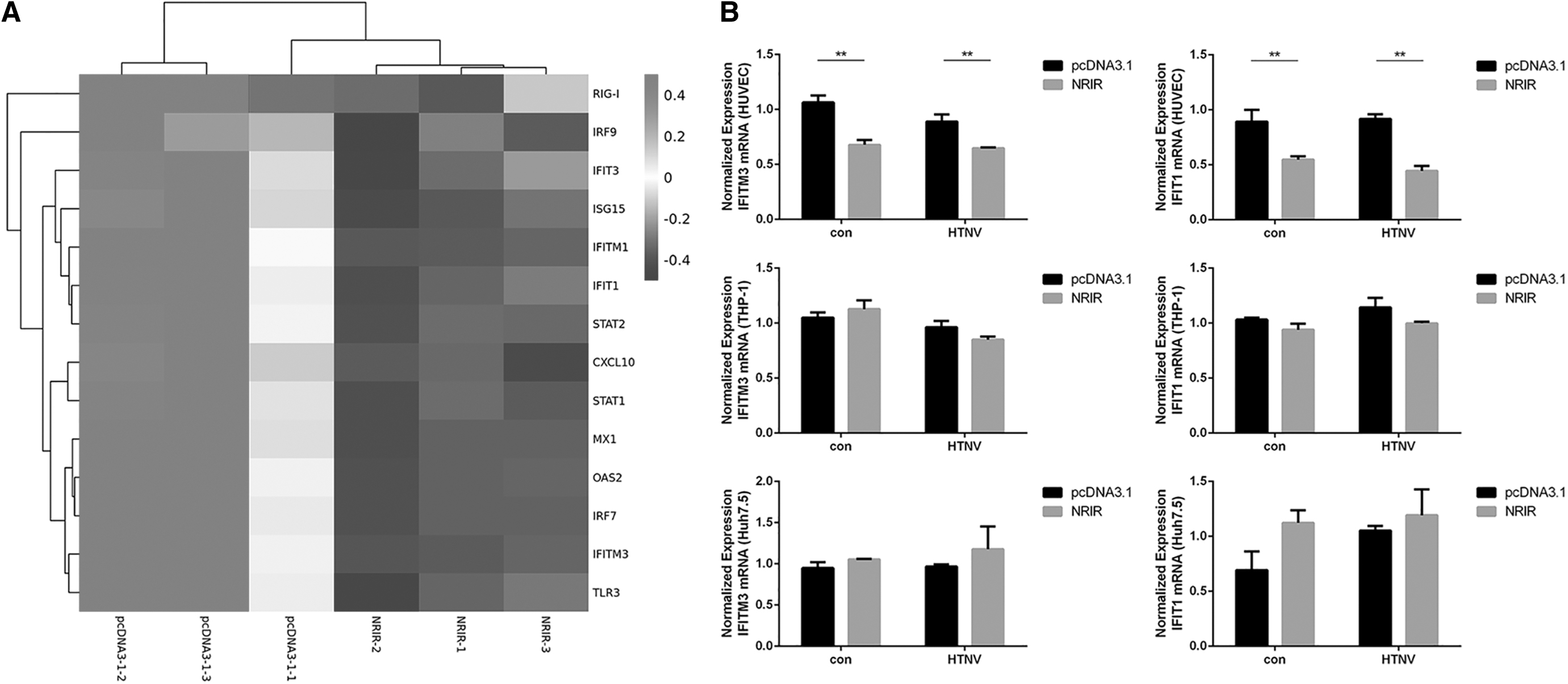
NRIR suppressed the expression of HTNV-related ISGs in HUVECs.

Subcellular localization of NRIR and its binding proteins
The subcellular localization of NRIR was examined using FISH in HTNV-infected HUVECs (MOI = 1, 72 h postinfection, Fig. 4A). NRIR was detected in the nucleus, similar to the distribution of U6. ChIRP-MS, in which the NRIR-binding proteins were identified through mass spectrometry, was performed in IFN-α2a-treated (1,000 U/mL, 12 h) THP-1 cells.
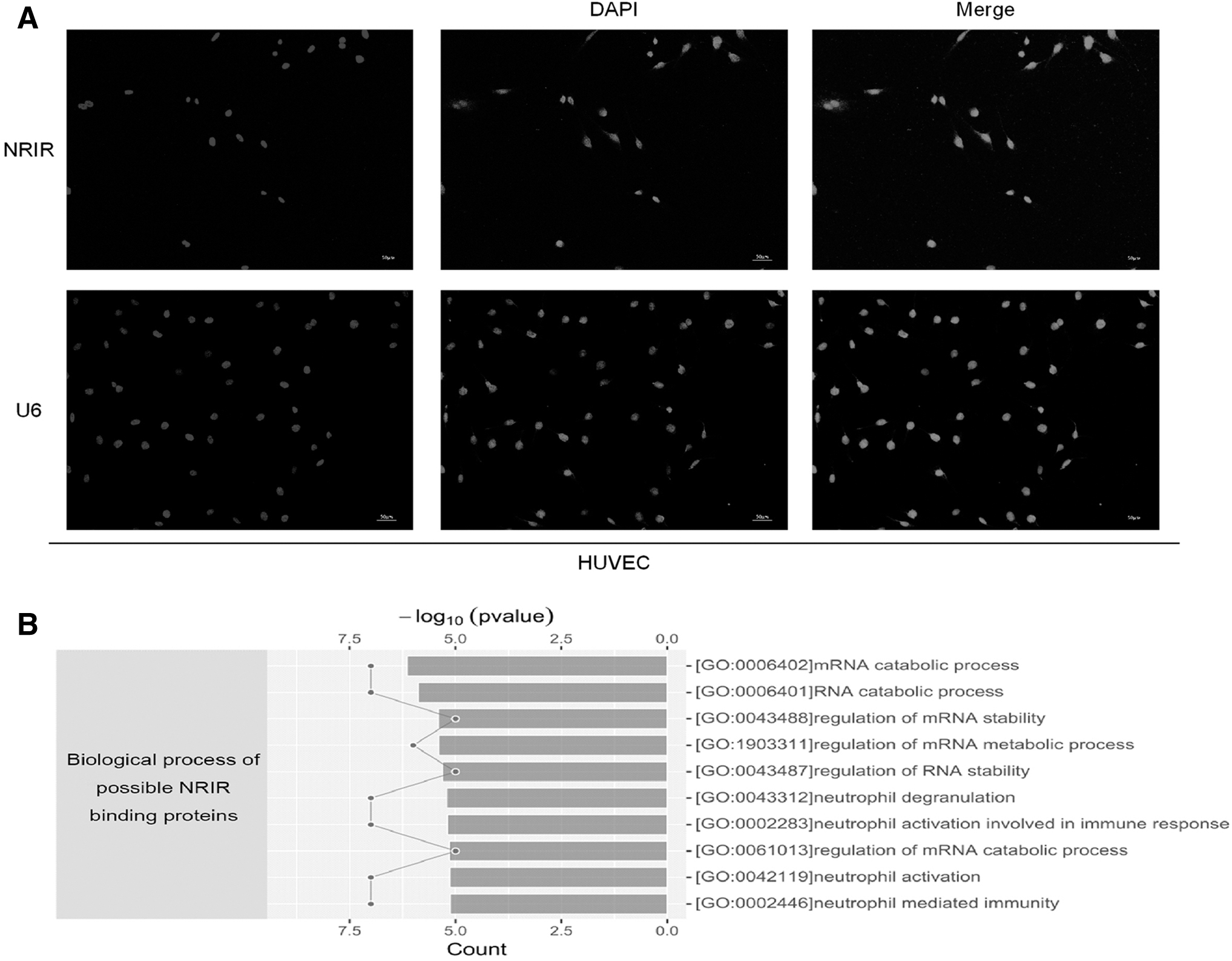
Subcellular localization of NRIR and identification of NRIR-binding proteins.
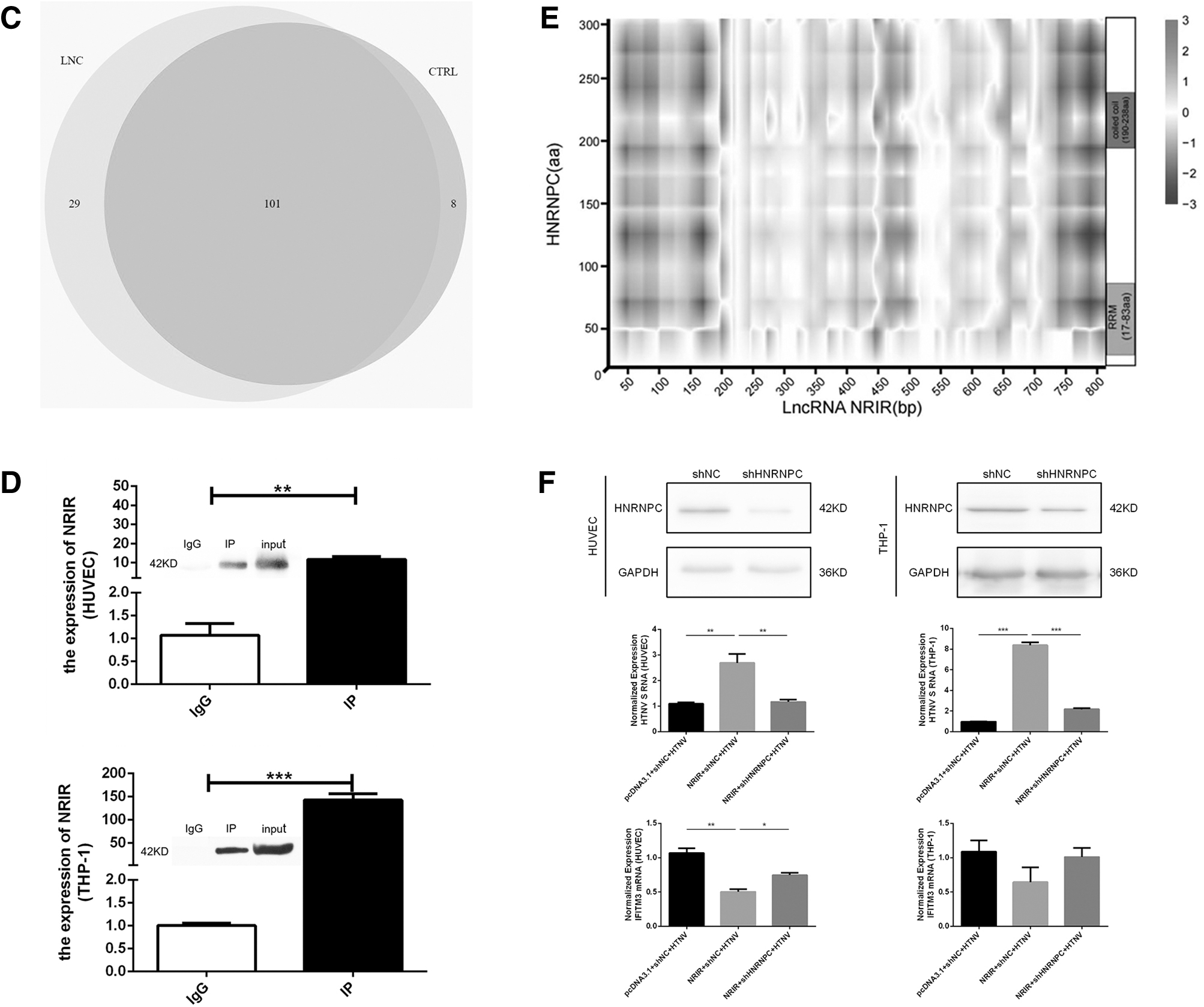
We identified 29 significantly enriched proteins (fold change >1.5, FDR <0.01, and unique peptide ≥2) in the NRIR probe group compared with the control group (Fig. 4C and Supplementary Table S4). Many enriched proteins were closely associated with mRNA and RNA-related biological processes (Fig. 4B). Among the 29 proteins, HNRNPC had the highest fold change between groups using the NRIR and control probes. We further confirmed the binding of NRIR to HNRNPC using RNA IP followed by RT-qPCR in THP-1 and HUVEC cells treated with 1,000 U/mL of IFN-α2a for 12 h (Fig. 4D). We also evaluated the propensity of the interaction between NRIR and HNRNPC using catRAPID omics. Possible bindings were predicted between two domains (RNA recognition motif and coiled-coil) of HNRNPC and specific fragments on NRIR (Fig. 4E, red parts).
To demonstrate the impact of NRIR-HNRNPC binding on HTNV infection, we overexpressed NRIR in HUVECs and THP-1 cells and knocked down HNRNPC using shRNA plasmids (Fig. 4F). On infection with HTNV (MOI = 1, 12 h), we found reduced levels of HTNV S in the NRIR overexpression and HNRNPC knockdown group compared with the NRIR overexpression and shRNA NC group in both HUVECs and THP-1 (Fig. 4F). At the same time, the expression of IFITM3 showed opposite trends with HTNV S in each comparison in HUVECs (Fig. 4F). In addition, we did not find significant effect of NRIR knockdown on the expression of HNRNPC, IFN-α2, and IFN-β (Supplementary Fig. S5A).
Discussion
lncRNAs have been shown to exert vital innate immune modulatory effects. However, a potential link between specific lncRNAs and the presence of type I IFN signature remains unclear in HTNV infection. To date, only lncRNA NEAT1, which exerts anti-HTNV effects, was intensively studied (Ma et al., 2017; Yang et al., 2022). NRIR was first characterized as an IFN-induced negative regulator of some ISGs. It was also upregulated in liver samples from HCV-infected patients (Kambara et al., 2014) and had clinical relevance in COVID-19 (Enguita et al., 2022) as well as systemic sclerosis (Mariotti et al., 2019). In the present study, we investigated the significance of NRIR as a regulator of the innate immune response against HTNV infection.
In vivo, HTNV primarily targets vascular endothelial cells and contributes to the increased vascular permeability and coagulation disorders. Hence, HUVEC is the most common model to explore innate immunity against HTNV. The THP-1 cell is used to mimic HTNV infection to monocytes. Additionally, Huh7.5 cells were reported to be HTNV permissive (Lee et al., 2011) and were used in the discovery of NRIR. In our study, HTNV infection induced NRIR expression in HUVECs and THP-1 cells but not in Huh7.5 cells, possibly due to the absence of RIG-I, which is important for IFN response against HTNV (Kell et al., 2020). In addition, the induction of NRIR by HTNV infection in HUVEC and THP-1 cells can be blocked by type I IFN receptor inhibitor Anifrolumab, further indicating the dependence of NRIR expression on the type I IFN. Subsequently, we found that HTNV NP could induce NRIR expression and it has been reported to preventing autophagosome formation (Wang et al., 2019) and contributing to miR-146a expression (Chen et al., 2017).
However, only HTNV infection could induce NRIR with comparable fold change to IFN treatment, indicating that the replication process of HTNV might be necessary for high NRIR expression. We also proved that NRIR expression in CD14+ monocytes were significantly higher in HFRS patients in acute phase compared with healthy controls and patients in convalescent phase. It is worth noting that NRIR was the only molecule with altered expression among lncRNAs and ISGs tested in CD14+ monocytes in this study (Supplementary Fig. S5B, C), suggesting its potential to be a clinical biomarker.
We altered the expression of NRIR in cells and infected them with HTNV. The overexpression of NRIR promoted HTNV infection to all three cells, which was consistent with our previous findings (Xu-Yang et al., 2016). The knockdown of NRIR before IFN-α2a treatment and HTNV infection showed the importance of NRIR among the numerous IFN-induced genes. We found that only the knockdown of NRIR in HUVECs resulted in a significant reduction of HTNV infection, which was in accordance with the findings of NRIR overexpression experiments. Some reasons may explain this phenomenon. (1) HUVEC has a more complete innate immune function compared with tumor cell line THP-1 and Huh7.5 (RIG-I-deficient). (2) The remaining NRIR after RNAi was active. (3) NRIR functions could be cell specific. Some authors found that the knockdown of NRIR reduced the expression of some ISGs in primary human monocytes, in contrast to the upregulation of ISGs in hepatocytes (Kambara et al., 2014; Mariotti et al., 2019).
lncRNAs employ diverse mechanisms to regulate the immune response. Nuclear lncRNAs may regulate transcription or mRNA processing (Suarez et al., 2020). For example, the levels of a nuclear lncRNA LUCAT1 elevated in inflammatory disease and restrain inflammation by regulating the splicing and stability of NR4A2 (Vierbuchen et al., 2023). We found that NRIR overexpression significantly inhibited the expression of many IFN-related genes in HUVECs, among which the inhibitory effects on IFITM3, which has anti-HTNV effect, was significant under basal and HTNV infection conditions. Some HTNV-related genes, such as ISGs and PRRs, were also downregulated after NRIR overexpression but IFN-α/β were unaffected. In HUVECs, when stimulated with equal amounts of IFN-α2a, IFITM3 expression increased in siNRIR+IFN group compared with siNC+IFN group, which proved the inhibitory effect of NRIR on ISGs from another perspective.
The nuclear localization of NRIR and its impact on the transcription of target genes were consistent with an epigenetic or transcriptional regulatory function. We found 29 possible NRIR-binding proteins by ChIRP-MS. The GO analysis revealed the relationship of these proteins with mRNA metabolism. Among these proteins, HNRNPC was the most promising candidate, and the binding between NRIR and HNRNPC was proved by RNA IP. The knockdown of HNRNPC significantly weakened the effect of NRIR in promoting HTNV infection in HUVECs and THP-1 cells and restored the IFITM3 expression inhibited by NRIR in HUVECs. As an RNA-binding protein regulating RNA splicing, sequence-unspecific RNA exportation, RNA expression, stability, 3′ end processing, and translation (Tang et al., 2022), HNRNPC has been related to the accumulation of endogenous double-stranded RNA, thereby activating the RIG-I pathway (Wu et al., 2018). HNRNPC was also involved in influenza virus infection (Tang et al., 2022) and DENV replication (Dechtawewat et al., 2015).
However, from this study and our previous RNA microarray (GSE86282), the expression of HNRNPC was not affected by HTNV infection and NRIR expression/knockdown, which implicated that NRIR may exert its effect by affecting the distribution or conformation of HNRNPC rather than its expression level. This deduction was in accordance with some previous researches on HNRNPC–lncRNA interactions (Cai et al., 2022; Yang et al., 2013). It is also possible that the NRIR–HNRNPC interaction was part of a larger multimolecular complex that regulates the innate immune response. The targets of NRIR in THP-1 may be different from that in HUVECs since IFITM3 expression was unchanged in THP-1 after NRIR overexpression and HNRNPC knockdown.
In conclusion, our study preliminarily revealed the role of NRIR in HTNV infection and provided new clues for further research on the immune regulatory mechanism of NRIR.
Footnotes
Acknowledgments
The authors acknowledge Prof. Fanglin Zhang and Yingfeng Lei from the Department of Microbiology, School of Basic Medicine, Air Force Medical University, Xi'an, China, for providing the expression plasmids of HTNV proteins and antibodies.
Authors' Contributions
Conceptualization, methodology, supervision: X.Z.,
Ethics Approval
The Institutional Ethics Committee of Tangdu Hospital approved this study. Before inclusion, all participants were informed of the study objectives and signed written informed consent.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This study was supported by the National Natural Science Foundation of China (81901619) and the Shaanxi Provincial Key Research and Development Program (2023-ZDLSF-32).
Supplementary Material
Supplementary Data
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.