
Abstract
Nasopharyngeal carcinoma (NPC) is a specific human malignancy with unique geographic distribution and genetic backgrounds. Although early treatment with radio-chemotherapy has been proven effective for NPC therapy, its therapeutic efficacy substantially diminishes in the late stages of this malignancy. In the tumor microenvironment of NPC, PD-L1 has been demonstrated as a critical factor in impairing T cell activation. As an etiological role for NPC development, it is found that Epstein-Barr virus (EBV) latent proteins upregulated PD-L1 expression. However, whether EBV lytic protein affects PD-L1 expression remains unclear. In this study, through monitoring the mRNA expression pattern of lytic genes and PD-L1 in EBV-positive NPC cell line NA, EBV immediately-early gene BRLF1(Rta) was found to have the potential for PD-L1 activation. Furthermore, we identified that Rta expression enhanced PD-L1 expression in mRNA and protein levels through quantitative real-time polymerase chain reaction and western blotting analysis. The luciferase reporter assay revealed that Rta expression enhanced PD-L1 promoter activity. We also demonstrated that Rta-induced PD-L1 expressions could impair interleukin 2 secretion of T cells, and this mechanism may be through ERK activation. These results displayed the importance of EBV Rta in PD-L1 expression in NPC and may give an alternative target for NPC therapy.
Introduction
Nasopharyngeal carcinoma (NPC) is a human malignancy arising from the postnasal cavity, prevalent in east southern Asia, China, and Taiwan. Chemotherapy combined with radiotherapy is a mainstream process to treat NPC. The early treatment has an efficient 5-year survival rate (80–95%) (Parkin et al., 2005). However, the treatment efficiency is poor in the late stage of NPC. Until now, advanced NPC is a significant cause of motility in NPC.
The etiology of NPC includes Epstein-Barr virus (EBV), dietary and genetic factors (Hildesheim and Levine, 1993; McDermott et al., 2001). EBV is a ubiquitous herpesvirus around the world, which is classified as gamma herpesvirus.
EBV is highly associated with human malignancies, including NPC and Burkitt's lymphoma. EBV has two life cycle stages: the latent and lytic cycles. Nine latent proteins (EBNA1–6, LMP1, LMP2A, and 2B) and two small RNAs (EBER1 and EBER2) are expressed at the latent stage. Upon activation, EBV enters the lytic phase and continuously expresses the immediate-early, early, and late genes, followed by viral particle releases (Rickinson and Kieff, 2001; Young and Rickinson, 2004).
The tumor microenvironment (TME) is the complex region around the tumor cells, which is now known to play a crucial role in tumor growth and progression. Surrounding with the TME, immune checkpoint (IC) molecules are frequently upregulated. Although upregulation of ICs could impair the antitumor activity of immune cells, the blockade of IC signaling to recover the anti-tumor activity of the immune system has become a novel therapeutic strategy recently. Among them, PD-1/PD-L1 signaling is the most famous IC pathway, triggered by the engagement with PD-1 and its ligands PD-L1 and PD-L2. Tumor cells overexpress PD-L1 to induce T cell exhaustion through PD-1/PD-L1 pathways. In NPC tissues, PD-1 and PD-L1 were detected as 37.4% and 95% positive rates, respectively, with no relevance to age, sex, and even NPC stages (Hsu et al., 2010).
Moreover, the expression of PD-L1 was detected in 89% of NPC tissues by IHC staining (Wotman et al., 2020). Of interest, IC molecules can help viruses and cancer cells avoid immune clearances in virus-associated malignancies. Many viruses were found to increase PD-L1 expression in human malignant cells (Channappanavar et al., 2012; Host et al., 2017; Huang et al., 2017b; Jones et al., 2019; Li and Zhang, 2021; Tu et al., 2010). For EBV, the upregulation of PD-L1 has been reported in EBV-associated malignancies, including Hodgkin's lymphoma (Carbone et al., 2018), Burkitt's lymphoma (Granai et al., 2020), NPC (Lv et al., 2019), gastric carcinoma (Morales-Sanchez and Fuentes-Panana, 2017), and so on. Several EBV latent proteins have been shown to induce PD-L1 expression.
Latent membrane protein 1 (LMP-1) was first identified to be able to activate PD-L1 expression through the nuclear factor kappa B (NF-κB) pathway in NPC cells (Bi et al., 2016; Fang et al., 2014). Moreover, EBNA2 induced PD-L1 expression in diffuse large B cell lymphomas (Anastasiadou et al., 2019) and Burkitt's lymphoma cell lines (Yanagi et al., 2022). Recently, it was found that EBV lytic cycle can also elicit PD-L1 expression (Yanagi et al., 2022). However, the question of which EBV lytic protein can induce PD-L1 expression remains unknown.
BRLF1 (Rta) is one of the immediate-early genes in the EBV lytic cycle. It encodes 605 amino acids, which is a transactivator to activate several lytic protein expressions and then promote the EBV lytic progression (Rickinson and Kieff, 2001). Rta activates the transcription of EBV lytic genes by directly binding to a Rta-responsive element, a GC-rich sequence, or indirect induction of signaling pathways, including phosphatidylinositol 3-kinase (PI3K; Darr et al., 2001), p38 MAPK, and JNK pathways (Adamson et al., 2000). In addition to activating EBV lytic cycle, Rta was reported to arrest the cell cycle in G1 phase (Chen et al., 2009), cause cell senescence (Chen et al., 2009), induce genomic instability (Huang et al., 2017a), and inhibit inflammatory response resulting from viral infection (Long et al., 2021).
In this study, we identified that Rta can activate PD-L1 expression. Moreover, this activation could decrease the T cell–secreting interleukin 2 (IL-2), implying that the EBV lytic cycle contributes to the exhaustion of T cells in the EBV-positive NPC cells. This finding will give a novel insight in immune evasion of EBV.
Materials and Methods
Reagents and antibodies
TPA (12-O-tetradecanoyl-phorbol-1,3-acetate), SB (sodium butyrate), PHA (phytohemagglutinin), and U0126 were purchased from Sigma-Aldrich Co. Rta antibodies used in this study was generated by our laboratory (Wu et al., 2016a; Wu et al., 2016b). Anti-PD-L1, anti-phospho-Erk1/2, anti-Erk1/2, and anti-β-actin antibodies were purchased from Cell Signaling Technology. Atezolizumab (Tecentriq) is a PD-L1 inhibitor purchased from Genentech, Inc.
Cell lines
TW01 is an NPC cell line derived from a Taiwanese NPC patient (Lin et al., 1990). NA is an EBV-positive NPC cell line derived from Akata strain EBV infecting TW01 cells. Both NPC cell lines were incubated in Dulbecco's modified Eagle's medium containing 10% fetal calf serum. Jurkat is a T cell line derived from the peripheral blood of a T cell leukemia patient (ATCC), which was incubated in RPMI-1640 containing 10% fetal calf serum.
EBV induction
EBV-positive NPC cell line NA (1 × 105) was seeded in a 24-well plate for 24 h. TPA (40 ng/mL) and SB (3 mM) were added to induce EBV reactivation. After treatment for the indicated time, the RNA or protein of each sample was extracted for further analysis.
RNA extraction and quantitative real-time polymerase chain reaction analysis
RNA extraction and reverse transcription are using commercial kits (Qiagen). Quantitative polymerase chain reaction (qPCR) was performed according to the manufacturer's instructions (Kapa Biosystems Fast qPCR kit). Indicated primers are listed in Table 1.
List of Primers Used for Amplification Quantitative Real-Time Polymerase Chain Reaction
Ectopic Rta expression and luciferase reporter assay
All plasmid transfection was performed by Lipofectamine 2000 (Invitrogen, Carlsbad, CA) according to the manufacturer's instructions. For ectopic Rta expression, empty vector or/and Rta-expressing plasmid were mixed with Lipofectamine 2000 in Opti-MEM (Invitrogen) for 20 min. The mixture was added to prepared cells in a six-well plate (2 × 106 cells/well). After 3 h of incubation, interferon γ (IFN-γ) or U0126 was added if needed. Following 24 h incubation, the cell lysates were harvested for western analysis. For luciferase reporter assay, the empty vector or Rta-expressing plasmid was mixed with pGL3 or pGL3-PD-L1 reporter in Opti-MEM containing Lipofectamine 2000 for 20 min. The mixture was added to the prepared cells in a 24-well plate (2 × 105 cells/well). After 3 h of incubation, IFN-γ was added if needed. Following 24 h incubation, the cells were lysed by HEPES buffer (0.1 M HEPES, pH 7.8, 1% Triton X-100, 1 mM CaCl2, and 1 mM MgCl2). The lysates and luciferase assay reagent II (Promega, Madison, WI) were coincubated with 1:1 volume ratio for 10 min. The luciferase activity was measured using a luminescence detector (Packard). The value of each sample was adjusted by β-actin expression to control the variation of each test (data not shown).
IL-2 secretion by Jurkat T cells
Jurkat T cells were treated with 50 ng/mL TPA and 1 μg/mL PHA for 24 h. Meanwhile, TW01 cells were transfected with vector or Rta-expressing plasmids for 3 h. Activated Jurkat T cells were cocultured with treated TW01 cells at a ratio of 5:1 for a further 48 h. IL-2 secretion in culture supernatants was detected by a human IL-2 enzyme-linked immunosorbent assay (ELISA) kit (ABclonal) following the manufacturer's instruction. The optical density values were measured by a microplate reader (Infinite M200, Tecan, Switzerland) at 450 nm with the reference wavelength at 630 nm. The value of standard deviation was obtained from three independent experiments.
Results
EBV reactivation increased PD-L1 expression in a time-course analysis of mRNA level
To determine whether EBV reactivation affected PD-L1 expression, we designed a time-course analysis to monitor the mRNA expression in the NPC cells. EBV-positive NPC cell line NA and its parental cell line TW01 were treated with TPA (40 ng/mL) + SB (3 mM) for 24 h. The mRNA expressions of various EBV lytic genes, including immediately-early genes Zta and Rta, early genes EAD and DNase, late gene gp350, and PD-L1, were detected at the indicated time points for RT-qPCR analysis. At the first 12 h, Zta and Rta were expressed slightly, followed by EAD and DNase expression at 15 h after EBV induction (Fig. 1, left panel). In the meantime, PD-L1 was also expressed at 12 h after induction, followed by the increasing expression for a further 15–24 h (Fig. 1, top of left panel). The EBV late gene gp350 was limited expressed in the first-day induction (Fig. 1, left panel), consistent with other studies (Wu et al., 2022; Yip et al., 2018). In the results of TW01 cell lines, PD-L1 was slightly expressed with TPA + SB treatment (Fig. 1, top of right panel). Moreover, there was no detection of Zta and Rta expressions with chemical induction in TW01 cells (Fig. 1, right panel). In addition, we determined the protein expression of Zta, Rta, and PD-L1 in NA cells and C666-1 cell lines. The results revealed similar phenomena to that of mRNA expression in NA cells (Supplementary Fig. S1). In addition, we checked the proportion of the cells coexpressed with Rta and PD-L1 through flow cytometry. The result revealed that coexpression of Rta and PD-L1 reached 46.3% of total cells in NA cells 24 h postinduction (Supplementary Fig. S2).
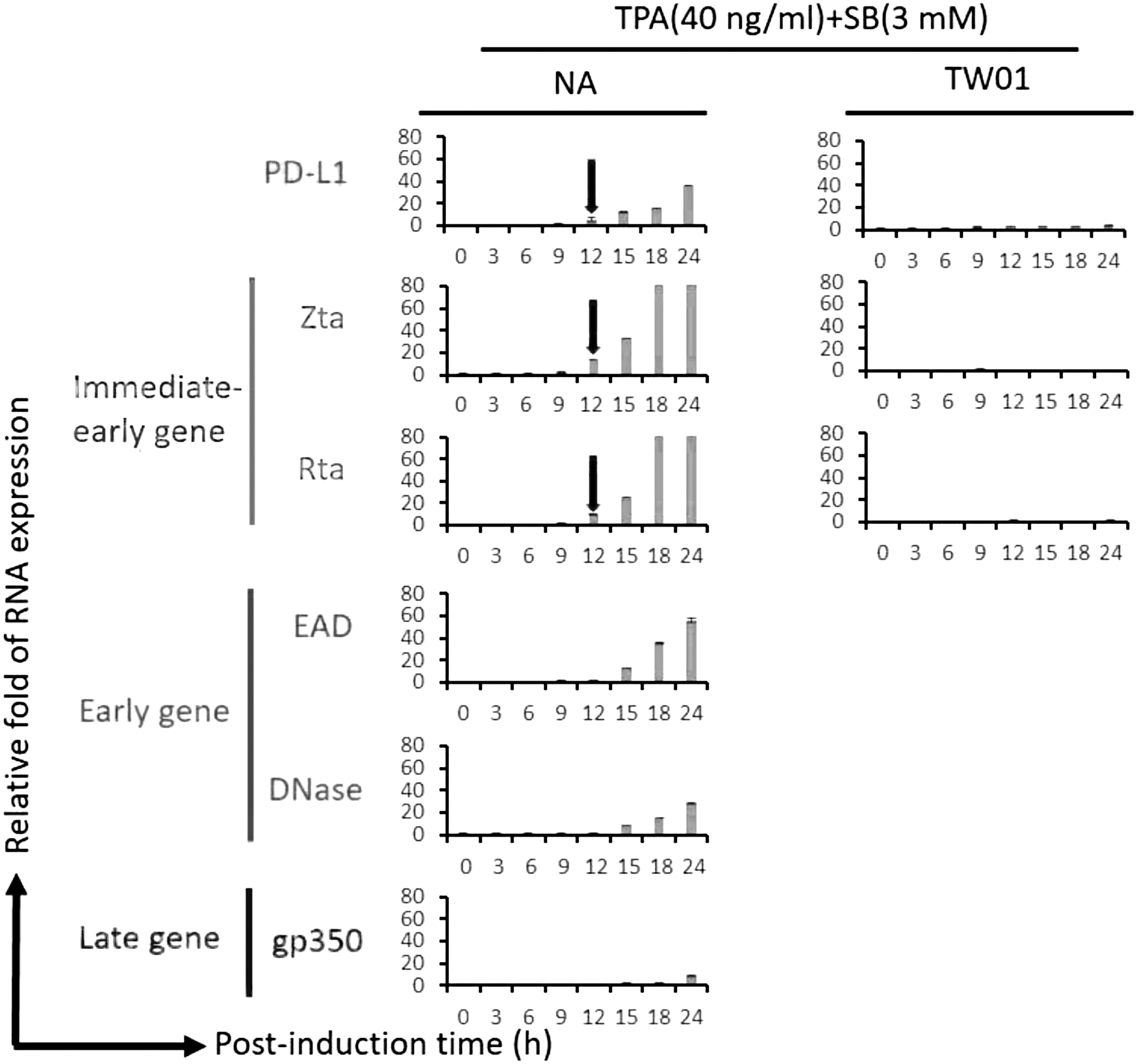
The expression patterns of EBV lytic proteins and PD-L1 within 24 h postinduction in the NPC cell line. To determine the relationship between EBV lytic progression and PD-L1 expression at the early induction stage, EBV-positive NPC cell line NA was induced by TPA (40 ng/mL) + SB (3 mM) and the mRNA expression of EBV lytic genes and PD-L1 were analyzed by RT-qPCR at the indicated time. Three substages of lytic genes (immediate-early, early, and late genes) were detected. The detailed protocols were described in the “Materials and Methods” section. The red arrows indicate that the PD-L1, Zta, and Rta RNA expressions were increased simultaneously at postinduction 12 h. The value of each point was obtained from three independent tests. Relative fold of RNA expression was represented as the relative fold of each sample's value compared with that at 0 h. EBV, Epstein-Barr virus; NPC, nasopharyngeal carcinoma; RT-qPCR, real-time quantitative polymerase chain reaction; SB, sodium butyrate; TPA, 12-O-tetradecanoyl-phorbol-1,3-acetate.
These results hint that EBV immediately-early genes may play roles in the induction of PD-L1 expression.
EBV immediate-early genes enhanced PD-L1 expression in NPC cells
Next, we further checked whether EBV immediately-early genes Zta or Rta could induce mRNA expression of PD-L1 in the NPC cell line. As given in Figure 2, as a potent inducer, the IFN-γ–treated group displayed a robust enhancement in mRNA expression of PD-L1 compared with mock control. Furthermore, Rta expression, but not Zta, induced a significant increase in PD-L1 expression compared with mock control (Fig. 2). Moreover, we determined whether coexpression of Zta and Rta could induce an additive effect on PD-L1 expression. As given in Figure 2, we did not observe the additive effect of PD-L1 expression that occurred by coexpression of Zta and Rta. We further found that the IFN-γ + Rta treatment group enhanced this PD-L1 induction by Rta significantly (Fig. 2).

The EBV immediate-early genes enhanced mRNA expression of PD-L1 in NPC cells. To determine whether Zta or Rta enhanced PD-L1 expression, Zta or/and Rta-expressing plasmid were transfected into TW01 cells with or without IFN-γ treatment for 24 h, respectively. The empty vectors transfected TW01 cells with or without IFN-γ treatment were used as mock and positive controls. The value of each sample was obtained from three independent tests. The relative fold of PD-L1 RNA expression was represented as the relative fold of each value compared with that of mock control. *p < 0.05; **p < 0.01. IFN-γ, interferon γ.
To demonstrate further, we checked the protein expression of PD-L1 induced by Rta expression. As given in Figure 3, the results showed the Rta transfection enhanced the PD-L1 expression in a dose-dependent manner at 24 and 48 h.

The ectopic Rta expression increased PD-L1 expression in NPC cells. TW01 cells were transfected with different dosages of Rta-expressing plasmids adjusted by vector plasmids for equal DNA amounts between each group. After 24 and 48 h incubation, cell lysates were collected for western analysis to detect Rta, PD-L1, and β-actin expressions.
Taken together, these results demonstrated that Rta can induce PD-L1 expression in NPC cells.
pGL3-PD-L1 reporter is activated by the Rta expression
The results above imply that Rta seems to be involved in the transcriptional regulation of PD-L1. To address this, the reporter plasmid harboring PD-L1 promoter (−460 to +1) has been used for luciferase reporter assay (Fig. 4a). As given in Figure 4b, in the groups with pGL3-PD-L1 transfection, IFN-γ treatment and Rta expression significantly enhanced luciferase activities compared with the mock control. In addition, as expected, the pGL3 transfected groups maintained a low level of luciferase activity (Fig. 4b). This result revealed that Rta induced PD-L1 expression by activating PD-L1 promoter activity.
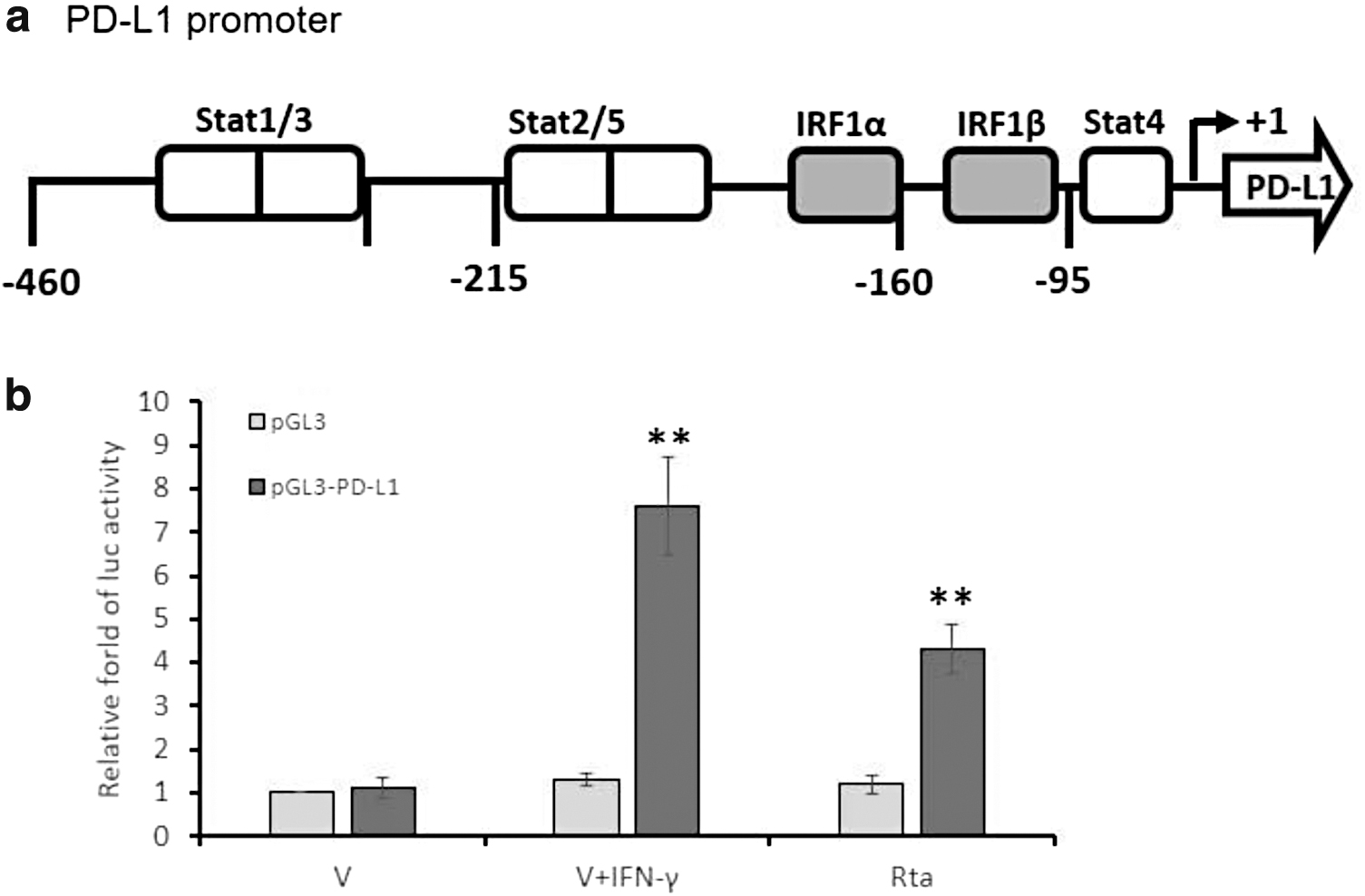
Rta expression activated PD-L1 promoter activity in the NPC cell line. The effect of Rta expression on PD-L1 promoter activity was determined by luciferase reporter assay.
Rta-induced PD-L1 expression attenuated the IL-2 secretion by T cells
It is known that PD-L1 expressed in cancer cells can cause the exhaustion of T cells (Freeman et al., 2000). Moreover, IL-2 is an essential indicator for T cell activation. We tried to determine whether Rta-induced PD-L1 expression could impair the IL-2 secretion from T cells. For this examination, Jurkat T cells were treated with TPA (50 ng/mL) and PHA (1 μg/mL) for 24 h to activate PD-1 expression. In the meantime, TW01 cells were treated with or without IFN-γ, or transfected with empty vector and Rta-expressing plasmids, respectively, for 24 h. Subsequently, Jurkat T and TW01 cells were cocultured at the ratio of 5:1 (Jurkat T: TW01). After 24 h coculture, the culture mediums were collected for detecting IL-2 secretion by ELISA assay.
As given in Figure 5, activated Jurkat T cells cocultured with IFN-γ–treated TW01 cells inhibited the IL-2 secretion robustly. Moreover, Rta expression displayed a significant decrease in the IL-2 secretion compared with vector control. If we treated with atezolizumab to block PD-1/PD-L1 signaling, the secretion of IL-2 was significantly recovered in both IFN-γ treatment and Rta expression groups (Fig. 5).
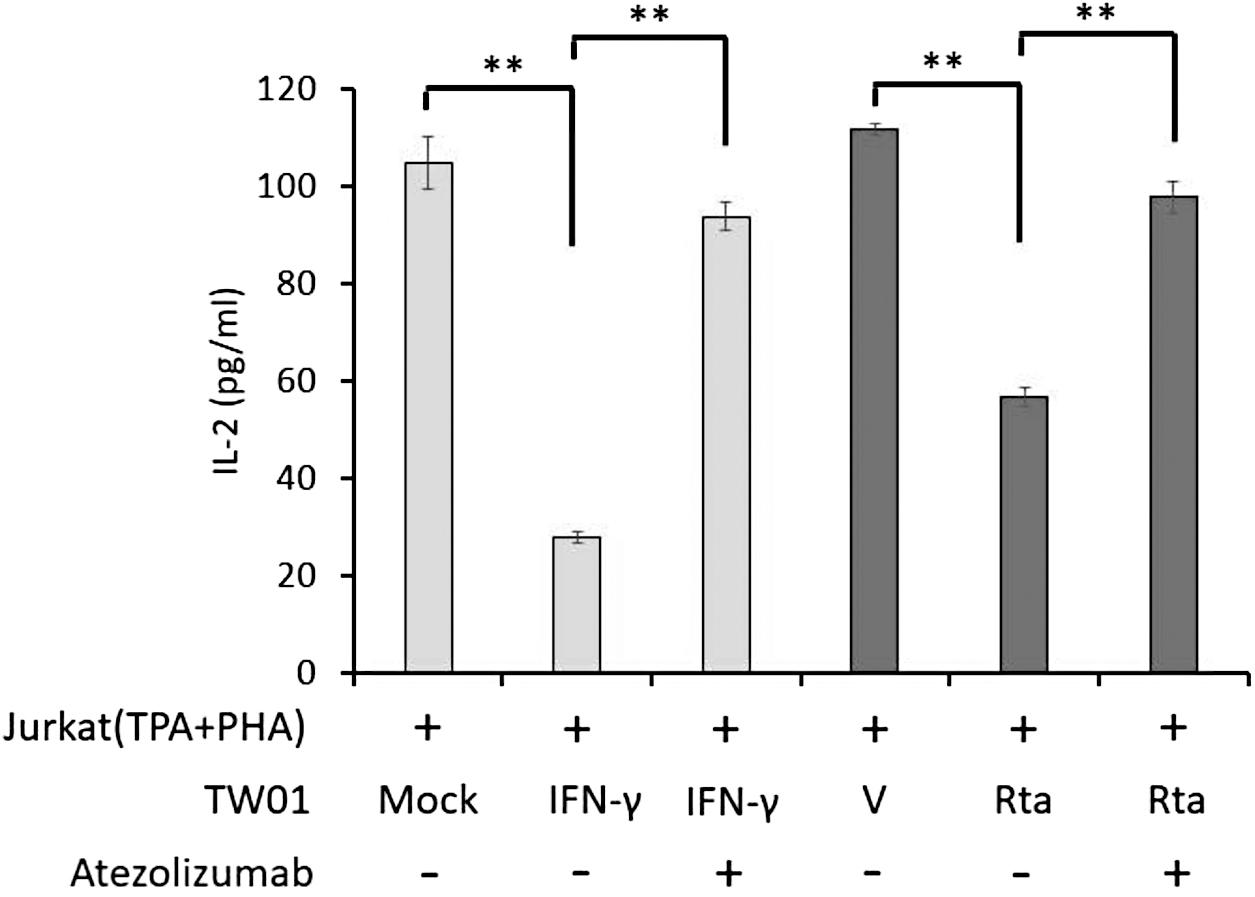
Impairment of IL-2 secretion by activated Jurkat T cells coculturing with Rta-expressed NPC cells. Jurkat T cells were pretreated with TPA + PHA (2 μg/mL) for 48 h. After that, activated T cells were cocultured with TW01 cells subjected for mock, IFN-γ and IFN-γ plus PD-L1 inhibitor atezolizumab, or transfected with empty vector and Rta plasmids with and without atezolizumab for further 24 h. Culture supernatants were collected to detect the amount of IL-2 by ELISA assay. The value of each sample was obtained from three independent experiments. **p < 0.01. ELISA, enzyme-linked immunosorbent assay; IL-2, interleukin 2.
These results revealed that Rta-induced PD-L1 expression is able to impair the IL-2 secretion by T cells.
ERK signaling played a critical role in Rta-induced PD-L1 expression
PD-L1 expression is regulated by many signal pathways, including the ERK pathway (Sun et al., 2018). Moreover, the ERK pathway is also an important signaling response for Rta activation (Lee et al., 2008). Based on this, we tried to determine whether the ERK pathway was involved in Rta-induced PD-L1 expression in TW01 cells. As given in Figure 6, Rta expression induced ERK phosphorylation and thus activated PD-L1 expression, which is the same as observed previously. However, this phosphorylation was gradually inhibited by increasing U0126 treatment. Meanwhile, the PD-L1 expression was also decreasing, suggesting that the ERK pathway is essential for Rta-induced PD-L1 expression. These results demonstrated that Rta induced PD-L1 expression through ERK signaling, possibly leading to T cell exhaustion.
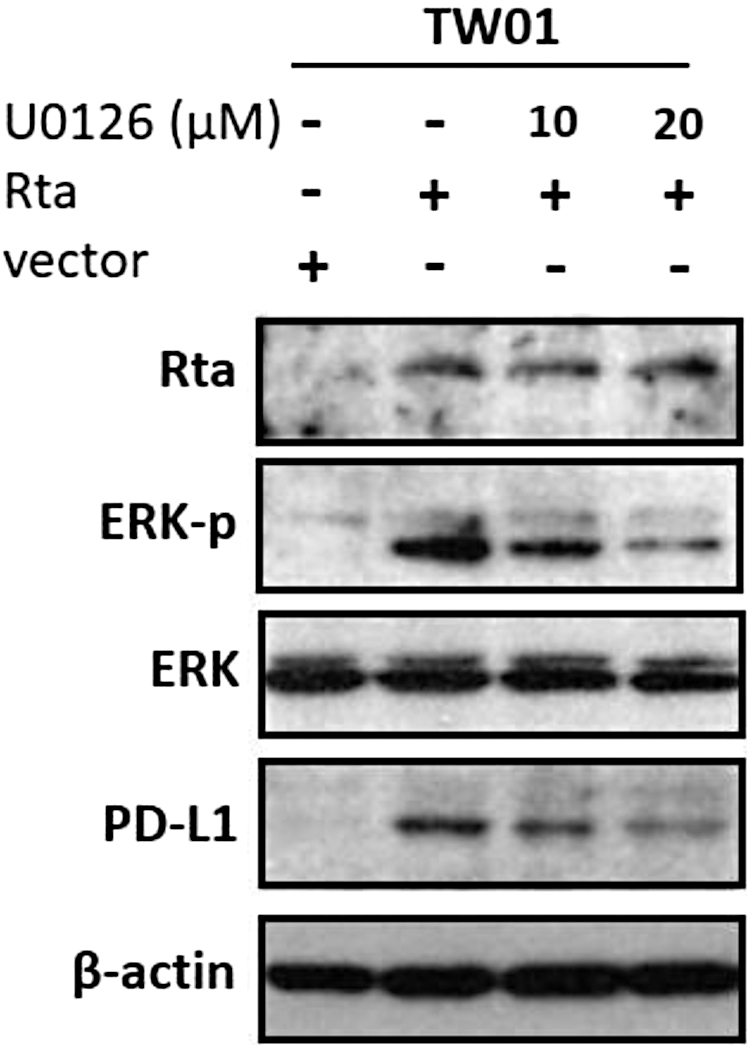
Erk signal pathway was involved in Rta-induced PD-L1 expression in NPC cells. TW01 cells were transfected with Rta plasmids and treated with different concentrations of Erk inhibitor U0126 (10 and 20 μM) for 24 h. Cell lysates were subjected to western blot analysis to detect the indicated protein levels. Cells transfected with empty vectors were represented as a control.
Discussion
IC pathways have now been known as critical factors in exploring cancer origins and therapeutic approaches. They largely weaken the capability of the human immune system to surveillance and destroy tumor cells (Anderson and Simon, 2020). In virus-associated cancers, IC expression also protects viruses' survival from immune clearance (Schonrich and Raftery, 2019). Therefore, studying the interaction between IC molecules and viruses is necessary for therapeutic purposes. As a well-known oncovirus, the interaction between EBV and PD-L1 was studied early (Anastasiadou et al., 2019; Fang et al., 2014). However, most of those studies focused on EBV latent proteins, not lytic ones. Considering the increasing importance of the EBV lytic stage on the oncogenesis of NPC, we tried to determine whether lytic proteins are involved in PD-L1 regulation in NPC cells. Through time-course analysis of mRNA expression, we found Rta, but not Zta, is a potent activator of PD-L1 (Figs. 1 and 2).
Moreover, when Rta was expressed in NPC cell line, an increase in PD-L1 expression and activation of the reporter with PD-L1 promoter were observed (Figs. 3 and 4). Subsequently, Rta-induced PD-L1 expression caused the impairment in IL-2 secretion by T cells (Fig. 5). We finally found that U0126 treatment inhibited PD-L1 expression, suggesting the ERK pathway may play an essential role in this phenomenon (Fig. 6). To our knowledge, Rta is the first lytic protein of EBV reported to own this ability in NPC cells. This finding also displays the importance of the EBV lytic cycle on PD-L1 regulation.
To escape host immunity, EBV evokes numerous viral proteins with immune evasion functions, which eventually help EBV to make a long-term persistence in host bodies. For latent proteins, EBNA1 inhibits translation and proteasomal degradation to keep a low level of viral antigen presented to CD8+ T cells. EBAN2 induces IFN-stimulated gene production and STAT3 activity to repress the production of inflammatory mediators (Ho and Ivashkiv, 2006; Muromoto et al., 2009). LMP-1 promotes cell growth by mimicking the constitutive CD40 signaling to activate numerous signals, including NF-κB, JAK/STAT3, and PI3K pathways (Middeldorp and Pegtel, 2008). Through regulating these signaling, LMP-1 can control the host immune response, including the induction of IRF7 and reduction of TLR9 expression (Fathallah et al., 2010; Huye et al., 2007). LMP2a and 2b have been shown to increase the turnover rate of the IFN receptor, which results in diminishing the host's sensitivity to IFN-α and IFN-γ (Shah et al., 2009). In addition, LMP-1 and EBNA2 were found to regulate PD-L1 expression in the last 10 years (Anastasiadou et al., 2019; Fang et al., 2014).
For lytic infection of EBV, several lytic proteins have been found to be contributing to the downregulation of the HLA I molecule. BGLF5 (DNase) expression degrades the mRNA of the HLA I molecule and then represses T cell recognition (Rowe et al., 2007). BNLF2a inhibits the transporter function associated with antigen processing (TAP) and reduces T cell recognition (Hislop et al., 2007). BILF1 downregulates HLA I presentation by interfering with the transportation and protein stability of the HLA I molecule (Zuo et al., 2009). In addition to evasion of CD8+ T cells, EBV also develops multiple mechanisms in the lytic stage to evade the recognition of CD4+ T cells: BZLF2 blocks the interaction between HLA II and TCR (Ressing et al., 2005), BGLF5 downregulates the mRNA of HLA II (Rowe et al., 2007), and BZLF1 impairs post-transcriptional HLA II presentation (Zuo et al., 2011). Besides, BCRF1, a viral IL-10 homolog, can inhibit CD4+ T cell responses (Zuo et al., 2011). This study demonstrated that EBV BRLF1 could induce PD-L1 expression to impair T cell functions and avoid T cell clearance. This finding also suggested the importance of the EBV lytic stage in PD-L1 regulation and NPC TME.
Although ERK pathway contributes distinctly to Rta-induced PD-L1 expression, there is another interesting question: whether another mechanism is involved in this phenomenon. Until now, several pathways, including RAS/MEK/ERK, PI3K/AKT, and JAK/STATs, are well-known as the major signals contributing to PD-L1 induction (Sun et al., 2018). On the contrary, Rta could also activate several signal pathways. In addition to ERK signaling, Rta is able to indirectly activate PI3K/AKT (Darr et al., 2001), p38 and JNK kinase pathways (Adamson et al., 2000) to accomplish its task of lytic induction. Comparing these hints, ERK pathway is reasonable to contribute to Rta-induced PD-L1 expression.
Moreover, the PI3K/AKT line should also be expected to play a specific role in this activation. In fact, in our preliminary screening, with treatment by MEK/ERK inhibitor U0126, PI3K inhibitor wortmannin, p38 MAPK inhibitor SB203580, and JNK inhibitor SP600125, U0126 inhibited PD-L1 expression effectively; the PI3K inhibitor revealed moderate inhibition and the rest showed less inhibition on PD-L1 expression (data not shown). This question is worth studying further.
In conclusion, we provide evidence to demonstrate that EBV Rta can induce PD-L1 expression, revealing the importance of the EBV lytic stage on PD-L1 expression. This finding provides a novel view of the interaction between EBV lytic stage and PD-L1 expression, giving an alternative choice for NPC therapy.
Footnotes
Acknowledgment
The authors thank Dr. Shao-Chih Chiu (China Medical University Hospital, Taichung, Taiwan) for critical comments and financial support on this article.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This work was supported by China Medical University Hospital, Taiwan (Grant No. DMR-110–184, DMR-CELL-2108, DMR-112-148) and National Science and Technology Council, Taiwan (Grant No. MOST 110-2320-B-039-009)
Supplementary Material
Supplementary Figure S1
Supplementary Figure S2
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.