
Abstract
Human papillomavirus (HPV) is a circular, double-stranded DNA virus and recognized as the most prevalent sexually transmitted infectious agent worldwide. The HPV life cycle encompasses three primary stages. First, the virus infiltrates the basal cells of the stratified epidermis. Second, there is a low-level expression of viral genes and preservation of the viral genome in the basal layer. Lastly, productive replication of HPV occurs in differentiated cells. An effective immune response, involving various immune cells, including innate immunity, keratinocytes, dendritic cells, and natural killer T cells, is instrumental in clearing HPV infection and thwarting the development of HPV-associated tumors. Vaccines have demonstrated their efficacy in preventing genital warts, high-grade precancerous lesions, and cancers in females. In males, the vaccines can also aid in preventing genital warts, anal precancerous lesions, and cancer. This comprehensive review aims to provide a thorough and detailed exploration of HPV infections, delving into its genetic characteristics, life cycle, pathogenesis, and the role of high-risk and low-risk HPV strains. In addition, this review seeks to elucidate the intricate immune interactions that govern HPV infections, spanning from innate immunity to adaptive immune responses, as well as examining the evasion mechanisms used by the virus. Furthermore, the article discusses the current landscape of HPV vaccines and common treatments, contributing to a holistic understanding of HPV and its associated diseases.
Introduction
Human papillomavirus (HPV)
HPV is a DNA virus from the Papillomaviridae family, which includes over 200 identified types (Van Doorslaer, 2022). HPV can infect both skin (cutaneous) and mucosal membranes in various anatomical locations, including the genital area and oral cavity (Van Doorslaer, 2022).
HPV stands as the most prevalent sexually transmitted infectious agent on a global scale (Kombe Kombe et al., 2020). It has the potential to cause or contribute to the development of cervical cancer and various types of epithelial malignancies (Tumban, 2019). Following infection, high-risk (HR) HPV can continue to persist, leading to the development of precancerous lesions. If left undetected and untreated, these lesions have the potential to progress into invasive cancer over time (Burger et al., 2017). Approximately 90% of cases of cervical intraepithelial neoplasia grade 1 (CIN1) occur in individuals who have tested positive for HPV. Moreover, over 99% of cervical cancer cases are also associated with HPV-positive patients (Liu et al., 2017).
According to a report from the World Health Organization, cervical cancer holds the position of being the fourth most common cancer among females, and represents roughly 6.6% of all female cancer cases (Hemmat and Bannazadeh Baghi, 2019). While certain risk factors, such as gene polymorphisms, are considered to contribute to the development of this particular type of cancer, it is worth highlighting that most cases are significantly linked to persistent HPV infections (Hemmat and Bannazadeh Baghi, 2019). In addition to cervical cancer, HPV infection has been established as a causal factor in several other cancer types, such as head and neck cancers, tonsillar carcinomas, and penile cancer (Hemmat and Bannazadeh Baghi, 2019).
The natural progression of HPV infection varies by gender and the specific anatomical site of infection (Giuliano et al., 2015). Gaining a comprehensive understanding of these differences is crucial for devising effective preventive measures aimed at preventing the various HPV-related cancers that affect both males and females (Giuliano et al., 2015).
HPV Genome
Papillomaviruses (PVs) are a type of DNA viruses characterized by a circular, double-stranded structure (Mirabello et al., 2018). Studies suggest that PVs have undergone coevolution with both animal and human host populations over the course of millions of years, and they are found worldwide, exhibiting a ubiquitous presence (Mirabello et al., 2018). The genetic material of the HPV is present in the form of an episome, consisting of around 8 kilobase pairs. This structure is divided into three distinct regions: two coding sections, which carry the instructions for producing viral proteins, and a noncoding region, responsible for regulating viral transcription and replication (Della Fera et al., 2021; Magalhães et al., 2021). Within the viral genome of HPV, the coding regions feature seven to nine open reading frames (ORFs) arranged into early and late regions. These ORFs are responsible for encoding two sets of highly conserved core proteins, E1 and E2, crucial for viral DNA replication, and L1 and L2, the structural proteins essential for capsid formation and vital in the assembly of virions (Della Fera et al., 2021; Magalhães et al., 2021). Beyond the core proteins, the HPV genome encompasses accessory genes such as E4, E5, E6, and E7. These accessory genes play a crucial role in modifying the host epithelium to establish a conducive environment for viral replication and aid in evading innate immune responses (Della Fera et al., 2021; Magalhães et al., 2021). The E1 and E2 proteins of HPV are primarily engaged in viral replication and the regulation of viral transcription. In contrast, the main oncogenes, E6 and E7, are thought to be instrumental in niche adaptation to specific anatomical or cellular regions, viral amplification, and unintentional stimulation of carcinogenesis (Mirabello et al., 2018). The noncoding section of the viral genome, also referred to as the upstream regulatory region, is situated ahead of the early region. It encompasses numerous cis-regulatory elements essential for transcriptional control, along with the origin of replication (Della Fera et al., 2021).
To date, researchers have identified a total of over 200 HPVs, which have been classified into five distinct categories: alpha, beta, gamma, mu, and nu PVs (Pešut et al., 2021). Typically, HPVs belonging to the alpha, mu, and nu genera are known to induce a diverse spectrum of skin warts. Conversely, HPVs belonging to the beta and gamma categories are primarily linked to asymptomatic skin infections (Warburton et al., 2021). In addition to causing skin warts, certain alpha PVs have the ability to infect the mucosal epithelial tissue situated at the mucocutaneous junctions found in body openings, including the oral and anogenital regions (Warburton et al., 2021). Within the various genera of HPVs, there is a subset of viruses that are classified as oncogenic. Persistent infection with these oncogenic HPVs can potentially result in the occurrence of anogenital or oropharyngeal cancers (Warburton et al., 2021).
Out of the approximate 60 types of alpha-HPVs, 13 have been categorized and labeled as unequivocally or possibly cancer causing (HR). These alpha-HPV types, known for their HR nature, contribute to more than 95% of cervical cancers globally. The specific types considered HR include HPV 16, 18, 31, 33, 35, 39, 45, 51, 52, 56, 58, 59, and 68 (Mirabello et al., 2018). Among the mentioned HPV types, each harbors various lineages and sublineages that demonstrate differences in intratypic genome sequences. These differences can range from 1.0% to 10% for lineages and from 0.5% to 1.0% for sublineages (Mirabello et al., 2018).
Specifically, HR HPV E6 aims to degrade the p53 tumor suppressor protein. By doing so, it prevents p53 from carrying out its normal cellular functions, such as orchestrating cell cycle arrest and triggering apoptosis in response to cellular stress signals (Lo Cigno et al., 2020). On the contrary, HR HPV E7 induces the pRb degradation, a protein known as the retinoblastoma tumor suppressor. This degradation process eliminates the E2F-mediated transcriptional activation of genes involved in the S-phase (Lo Cigno et al., 2020). Significantly, both HPV E6 and E7 are pivotal in inducing epigenetic changes within the chromatin. These changes occur through the alteration of regulation of gene expression or enzymatic activity of various epigenetic modifiers, encompassing histone deacetylases, histone demethylases, histone acetyltransferases, and histone methyltransferases (Lo Cigno et al., 2020).
The application of next-generation sequencing technologies in studying numerous primary cervical cancers has led to significant advancements. These recent findings indicate that the conventional model of HPV-induced carcinogenesis requires revision (Gao et al., 2019). A comprehensive review of numerous cases of cervical cancers positive for HPV16 and HPV18 has uncovered an interesting distinction. It has been observed that the incorporation of HPV within the human genome is more prevalent in HPV18-positive cervical cancers, occurring in the majority of cases. However, this integration is detected in only about 75% of HPV16-positive cervical cancers (Gao et al., 2019). Furthermore, research found that there is no specific preference for the HPV E2 gene alteration in cervical cancer. Breakpoints, which indicate the sites of genetic rearrangements, were observed to occur across the entire HPV genome in cervical cancer cases (Gao et al., 2019). Moreover, it has been identified that there are specific clustered hotspots where HPV integrates into the human genome. Notably, these integration sites often coincide with important cancer-related genes or are located in close proximity to them. This implies a possible connection between HPV integration and the dysregulation of crucial genes involved in cancer development (Gao et al., 2019). As a result, it has become evident that crucial human genes can play a role in the process of carcinogenesis. Consequently, the specific location(s) where HPV integrates into the human genome may hold greater significance than originally anticipated. This highlights the importance of studying the integration patterns and their potential influence on cervical cancer progression (Gao et al., 2019). The utilization of whole-genome sequencing has provided valuable insights into the intricate nature of the integration of HPV18 in the HeLa cell line. This analysis has revealed that the integration event takes place within a specific hotspot adjacent to the c-MYC oncogene, which is unique to HPV18. This finding highlights the complexity and specificity of the integration of HPV18 in the HeLa cell line (Gao et al., 2019). Also, the integration of HPV18 is linked to selective amplification of HPV18 sequences as well as neighboring human sequences. The total size of the disruption area resulting from this integration is estimated to be over 300 kb (Gao et al., 2019) (Fig. 1).
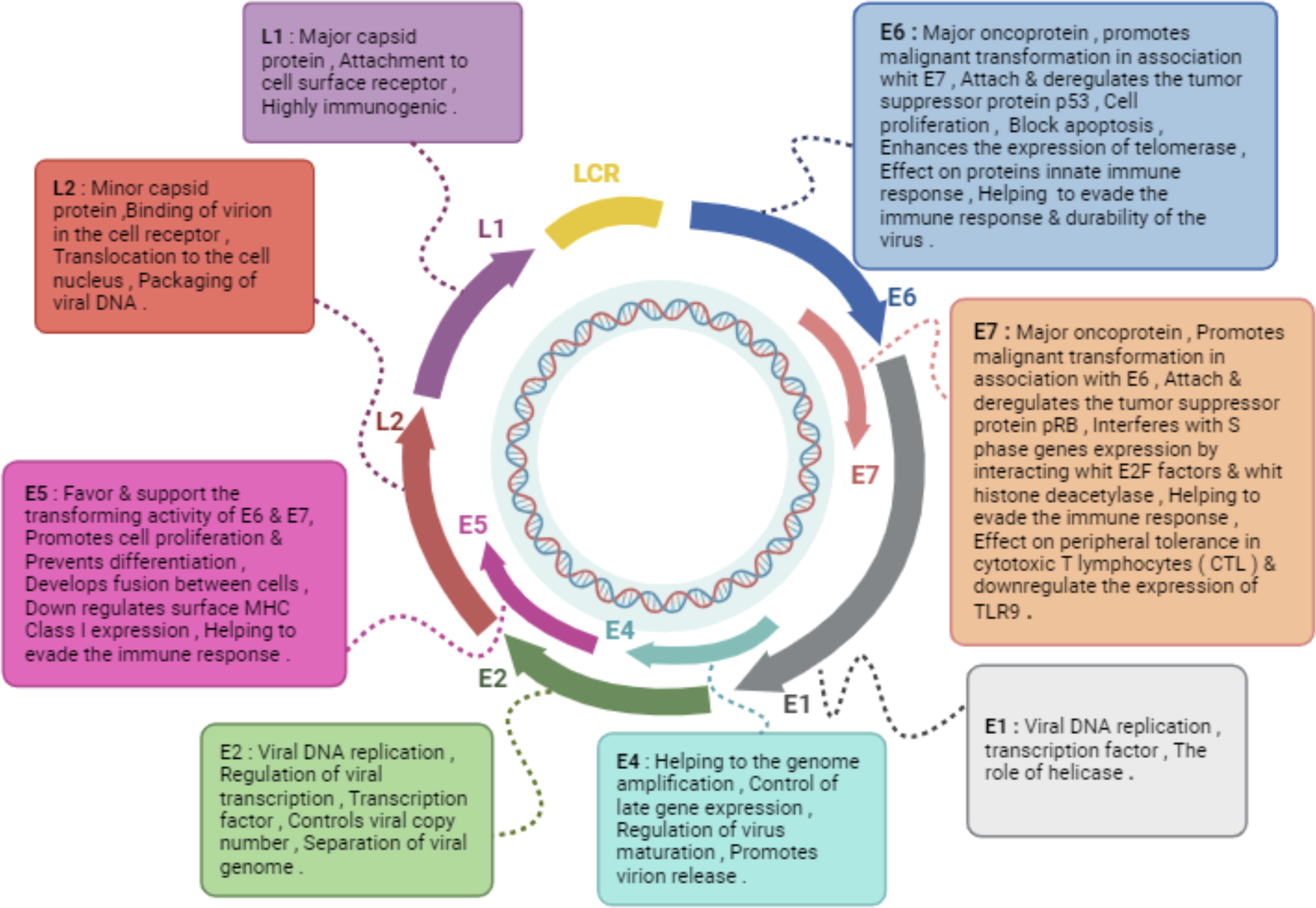
Schematic representation of functional properties of HPV proteins.
There is poor correlation between HPV genetic relatedness and carcinogenicity: HPV31 and HPV35 are the types most closely related (genetically similar) to HPV16, but they are much less carcinogenic. At the same time, HPV18 is highly carcinogenic, but it is relatively distantly related to HPV16 and preferentially causes glandular lesions (Nelson and Mirabello, 2023).
Life Cycle
The life cycle of HPV, contingent on cellular differentiation, can be succinctly described in three stages as follows: The penetration of HPV into the basal cells located in the stratified epidermis. The basal layer exhibiting minimal viral gene expression and ensuring the upkeep of the viral genome. The productive replication of HPV taking place in the differentiated cells (Kajitani et al., 2012).
HPV infection is primarily transmitted through direct contact between the skin or mucous membranes of individuals (Brianti et al., 2017). HPV targets and infects areas of the stratified epithelium that have been compromised, such as wounds or other forms of epithelial damage (Brianti et al., 2017). HPV infects epithelial cells by interacting with specific receptors on the cell surface, such as integrin α6. These receptors are plentifully present in basal cells and epithelial stem cells (Brianti et al., 2017).
In most cases, most of the HPV genome persists in an episomal condition within infected cells and premalignant lesions. It is believed that the virus infects keratinocytes and remains dormant, usually kept under control by the immune system (Christensen, 2016). Symptoms are not observed during either the transmission of HPV from one person to another or the transmission from an infected person to unaffected cells within the body (Christensen, 2016). Following resolution of the infection through immune system activity, it is plausible for the HPV to endure in a latent state within squamous cells that do not exhibit dysplasia. Consequently, Papanicolaou (Pap) smears have the potential to yield normal results. Nonetheless, it is imperative to maintain regular Pap screenings as per professional recommendations to effectively monitor any prospective alterations in cellular morphology (Christensen, 2016). In cases where the patient’s immune system is suppressed, the inactive virus has the potential to become contagious, leading to the possibility of basal epithelial cell differentiation in the infected individual (Christensen, 2016). Most HPV infections are temporary and exhibit no symptoms, and they are typically eradicated by the immune system (Christensen, 2016).
To generate virions and successfully conclude the infectious cycle, the HPV life cycle does not involve a lytic phase and instead relies on the final keratinocyte differentiation program as a necessary requirement (Ilahi and Bhatti, 2020; Pinidis et al., 2016). It is hypothesized that infection takes place when a microabrasion exposes the proliferative cell layer (Basal Stem Cells) to an HPV virion (Fisher, 2015).
After entering the cell, the viral DNA is in an unenveloped state and undergoes acidic compartments such as late endosomes and lysosomes (Fisher, 2015). The main role of the viral capsid protein L2 is to facilitate the transportation of the viral DNA to the nucleus by interacting with various cytoplasmic partners in a series of steps (Fisher, 2015).
In the initial phase within the basal cell layer, the viral DNA undergoes amplification to approximately 50–300 copies within each cell upon entering the nucleus (Fisher, 2015; Pinidis et al., 2016). Every protein encoded by the HPV genome is integral to the HPV life cycle [14]. The E1 and E2 viral proteins work together to commence the replication of viral DNA and establish viral genomes (Fisher, 2015; Mac and Moody, 2020). The HPV DNA persists in actively dividing the basal cell layer for an extended period of time at a relatively consistent copy number. In the S-phase of the cell cycle, the HPV DNA is replicated utilizing the host cell’s replication machinery (Gupta and Mania-Pramanik, 2019). Most antiviral strategies primarily focus on the maintenance phase of replication, given that persistent infection by HR HPV is the main risk factor for cervical carcinogenesis (Gupta and Mania-Pramanik, 2019). During the maintenance phase, replication takes place through either an ordered or random mechanism, influenced by the nuclear environment and the levels of E1 in the host cell (in the ordered mechanism, each episome is replicated once per cell cycle, while in the random mechanism, some episomes replicate multiple times while others do not) (Gupta and Mania-Pramanik, 2019). Extensive research has demonstrated the essential role of viral proteins E1, E2, E6, and E7 in both the establishment and continual maintenance of viral DNA episomes within host cells (Murakami et al., 2019).
The vital helicase function of the highly conserved HPV protein E1, operating in the 3′ to 5′ direction, plays a crucial role in HPV genome replication and is believed to be significant for the sustained maintenance of HPV (Fisher, 2015; Mac and Moody, 2020). The function of E1 is reliant on the hexameric helicase assembly at the origin of replication (ori) in collaboration with E2 (Fisher, 2015; Mac and Moody, 2020), which is entirely dependent on the cooperation between E1 and the host cell replication machinery, as they closely interact to enhance bidirectional HPV genome replication (Fisher, 2015). E2 plays a multifaceted role, serving as a crucial protein involved in both viral DNA replication and transcriptional control. This dual function is essential for the preservation of episomes (Zhang et al., 2019). E2 is thought to regulate the early promoter by exerting both activating and inhibitory effects on its own expression, along with that of E6, E7, and E1 (Fisher, 2015; Magalhães et al., 2021). E8^E2C represents a shortened form of E2 that seems to hold significance during the establishment phase. It functions to limit early genome amplification and inhibit HPV replication, ultimately contributing to the maintenance of a stable copy number. This reinforces the notion of E2’s crucial role in copy number regulation (Fisher, 2015). In addition, E2 has a distinctive function in tethering HPV genomes to mitotic chromosomes, guaranteeing the accurate separation of viral DNA as cells divide (Prabhakar et al., 2023). The trans-repressive role of E2 on the early promoter, which occurs post-HPV DNA integration, is believed to contribute to carcinogenesis by increasing the expression of oncogenes (Kalathil et al., 2020).
Crucially, the introduction of external E2 into cervical cancer cells hosting integrated HPV genomes lacking functional E2 genes reinstates the regulation of E6 and E7 oncogenes, leading to apoptosis or senescence. This demonstrates the persistence of apoptotic and senescence pathways, which are, however, inhibited by viral oncogenes in invasive cervical cancer (Fisher, 2015; Pinidis et al., 2016).
The E7 oncoprotein accelerates cell cycle advancement and enhances replication competence in basal cells by promoting the degradation of the pRb tumor-suppressor protein, and by interacting with a variety of cellular components, including the activation of CDK2 (Fisher, 2015; Pinidis et al., 2016). Likewise, the versatile oncoprotein E6 serves multiple functions, primarily incapacitating p53, but it also inhibits apoptosis and regulates several cellular processes, including immune evasion. Furthermore, it promotes an extended lifespan by inducing telomere maintenance through hTERT (Fisher, 2015; Pinidis et al., 2016).
Working in tandem, E6 and E7 induce substantial alterations in the epithelial transcriptome, fostering an environment conducive to finely tuned cell cycle progression that supports viral DNA maintenance and productive replication. Simultaneously, they safeguard against senescence, extending the lifespan of the host cell (Fisher, 2015).
In a wart, a secondary amplification stage takes place, giving rise to infectious virions (Fisher, 2015). As cells undergo differentiation and migrate from the basal layer to the suprabasal layers of the epithelium, there is a significant upsurge in the transcripts of E1, E2, E1^E4, E5, and E8. This phenomenon leads to substantial amplification of viral episomes (Fisher, 2015).
The viral oncogenes E6 and E7 significantly contribute to DNA amplification in this stage as they assist in establishing an environment conducive to the progression of the cell cycle and synthesis of DNA within the differentiated, postmitotic cells (Burley et al., 2020) (Fig. 2).
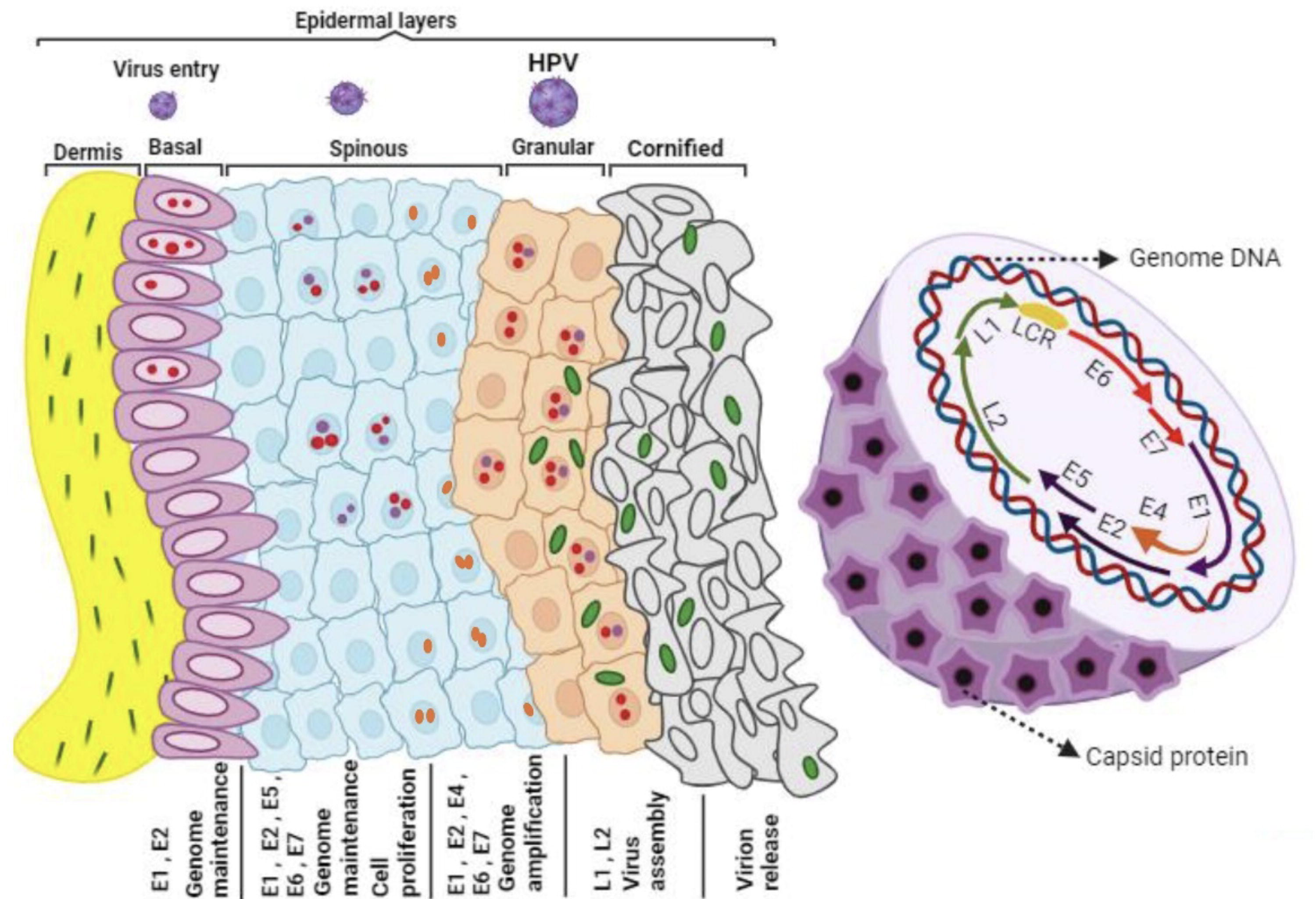
High-risk HPV life cycle: From basal cells to disease progression. High-risk HPVs infect the basal cells in the ectocervix by accessing microwounds. These cells serve as an infection reservoir, maintaining the viral genome. As they divide, daughter cells migrate toward the surface. In lesions caused by high-risk HPV types, cells express E6 and E7 proteins, driving cell division, whereas mid-layer cells exhibit elevated proteins for genome amplification. Upper layer cells exit the cell cycle and produce L2 and L1 proteins for packaging viral genomes. Infection may also extend to other cell types, affecting disease progression and contributing to adenocarcinoma development at the cervical transformation zone and endocervix.
HPV Pathogenesis
In most instances of cervical cancers, the viral genetic material becomes integrated into the cellular genome, which facilitates the transformation of the infected cells (Hampras et al., 2014). During the integration process, viral DNA breakpoints induced by HPV E1, E6, and E7 proteins or oxidative stress contribute to the increased expression and translation of viral oncogenes, particularly E6 and E7 (Folliero et al., 2023). Available evidence indicates that infection with cutaneous HPV types elevates the likelihood of nonmelanoma skin cancer in both individuals with a functioning immune system and those with compromised immunity, possibly through an indirect mechanism (Hampras et al., 2014). One proposed mechanism of cutaneous HPV carcinogenic activity is the inhibition of UV-induced apoptosis (Hampras et al., 2014).
Most HPV infections are typically limited to a temporary duration and will be cleared spontaneously, making them of a transient nature (Sadri Nahand et al., 2020). Contracting various types of HPV types can lead to benign conditions that are frequently subclinical. However, they may progress to the development of papillomas or warts on the epithelial surface (Mac and Moody, 2020). Nevertheless, the prolonged infection with HR strains of HPV, which are classified as group I carcinogens and are responsible for cancers in the anogenital tracts and oropharynx, can give rise to the emergence of a premalignant condition referred to as cervical neoplasia (Mac and Moody, 2020; Sadri Nahand et al., 2020). The incorporation of HPV E7 and E6 oncogenes into the host genome is regarded as a critical step in the progression of cervical cancer (Sadri Nahand et al., 2020). Immune evasion is a significant factor in the persistence of HPV infection (Hatano et al., 2021). The prolonged infection with HR strains of HPV, characterized by the incorporation of HPV E7 and E6 oncogenes into the host genome, is regarded as a critical step in the progression of cervical cancer (Kombe et al., 2023). No detectable viremia or cytolysis is evident during the early stages following the primary HPV infection in the cervix, which means that inflammation is not readily observable, and the innate immune system remains inactivated (Ujma et al., 2019). The completion of the virus’s life cycle occurs within actively dividing cells, utilizing the host cellular machinery (Sadri Nahand et al., 2020). Upon establishing a persistent infection, it induces alterations in the secretion of inflammatory cytokines, leading to the infiltration of immune cells (Sadri Nahand et al., 2020). In older individuals experiencing persistent HPV infection, there have been observations of changes in immune system responsiveness and elevated systemic levels of inflammatory cytokines (Bhuyan et al., 2022). Clinical studies have indicated that individuals in this age bracket are more commonly diagnosed with cervical cancer and are prone to exhibit a resistant increase in cytokines, contributing to the tumorigenesis associated with HPV (Das et al., 2023). Inflammation constitutes a vital aspect of the intricate biological response initiated by the body’s tissues in reaction to detrimental stimuli, which can include pathogens, irritants, injured cells, extreme temperatures, radiation exposure, or autoimmune processes (Nakkala et al., 2021; Sadri Nahand et al., 2020). Persistent HPV infection can lead to alterations in the secretion of inflammatory cytokines, contributing to the infiltration of immune cells and chronic inflammation, which is closely linked to HPV-associated cervical cancer (Avila et al., 2023). Inflammation is a designed protective response that involves blood vessels, immune cells, and molecular mediators (Zhao et al., 2021). The onset of inflammation swiftly releases chemokines and cytokines, triggering the innate immune response of the host (Zhao et al., 2021). These mediators collaborate and aid in recruiting effector cells to the location of injury or infection (Zhao et al., 2021). If the stimulus persists continuously, the initial acute inflammation can evolve into a chronic state, and chronic inflammation is closely linked to the onset of cancer (Zhao et al., 2021).
However, the involvement of HPV infection in triggering chronic inflammation and the connection between chronic inflammation and HPV-induced cervical cancer remain subjects of debate and controversy (Kudela et al., 2021). The development of cervical cancer necessitates several factors, including viral interactions, host-dependent factors, and environmental influences (Kudela et al., 2021). Moreover, there is supporting evidence suggesting that the epigenetic regulatory mechanisms may result in the aberrant regulation of tumor-suppressor genes and the activation of oncogenes, promoting the malignant phenotype in cancer cells (Kudela et al., 2021). In this context, microRNAs function as crucial regulators controlling cell cycle progression, apoptosis, metastasis, and resistance to chemical treatments (Kudela et al., 2021) (Fig. 3).
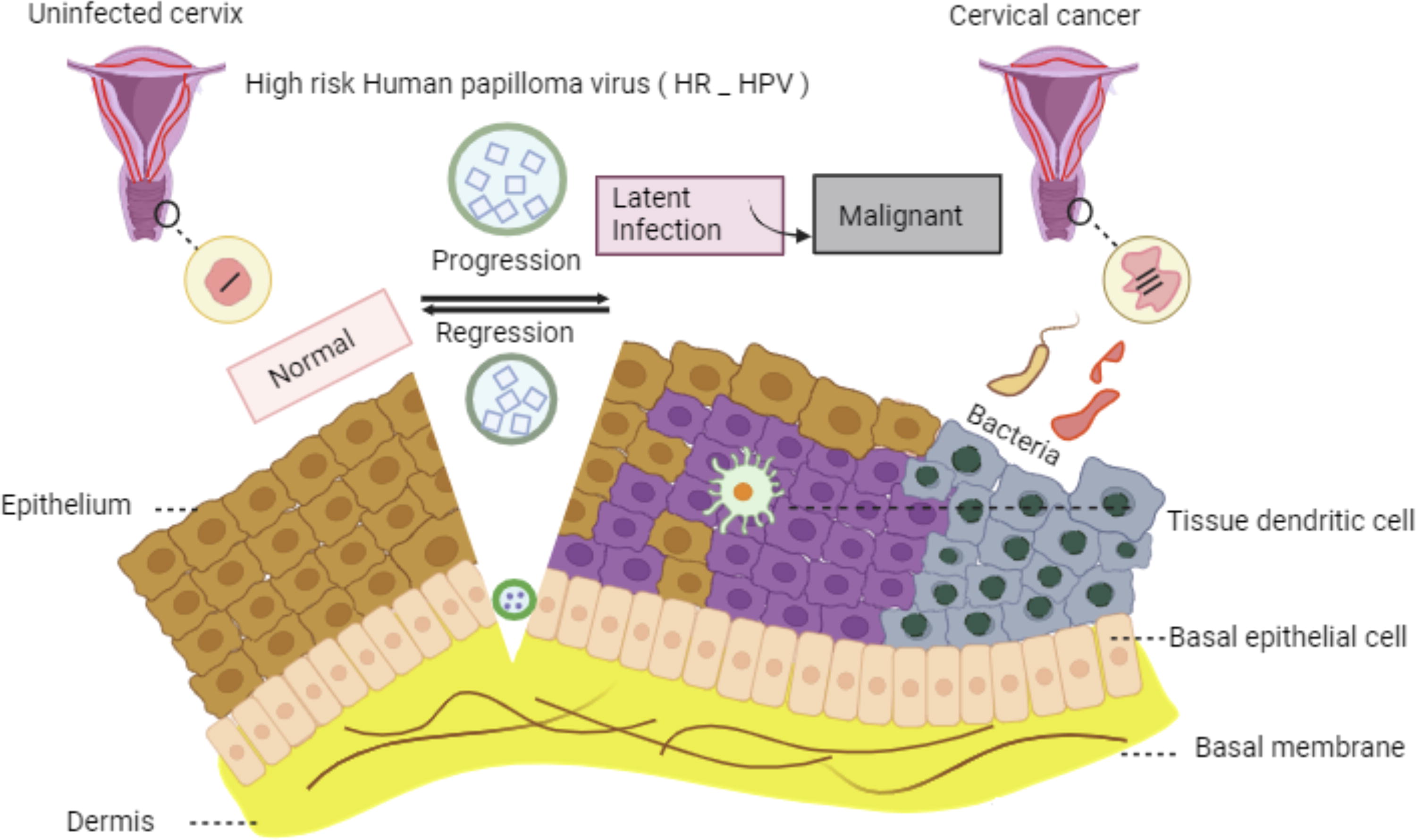
The pathogenesis and natural progression of high-risk HPV infection. In the initial stage, the virus enters the basal layer of the epithelium through a scratch or microinjury, initiating gradual proliferation. Although most infections resolve spontaneously, some may remain latent for an extended period. In the final stage, the genetic material of the virus integrates into the human genome, resulting in increased replication rates and ultimately contributing to the progression of cervical cancer.
HR or Oncogenic HPV Types
Based on epidemiological and molecular biological data, acquiring specific types of HPV stands out as the most prevalent risk factor contributing to the progression of cervical cancer (Muñoz-Bello et al., 2022). Over 200 HPV variations have been recognized, distinguished by genomic variations, and they are typically categorized into low-risk (LR) and HR types (Miyagi et al., 2019). This classification is rooted in the virus’s oncogenic potential and its tendency to induce cancer, which is linked to its presence in either benign or malignant cervical lesions (Orlando et al., 2012). LR and HR-HPV types exhibit distinct molecular disparities, and they also differ in terms of their infection duration within a host as well as their modes of transmission between hosts (Orlando et al., 2012).
The carcinogenic HR types encompass HR-HPV 16, 18, 31, 33, 35, 39, 45, 51, 52, 56, 58, 59, 66, 68, 73, and 82, indicating their association with cancer (Colpani et al., 2020). In the HR category, certain HPV strains are infrequently detected in cancers but are often found in squamous epithelial lesion cells (Miyagi et al., 2019). In the category of HR HPV infections, HPV16 and 18 stand out as the predominant types connected to cervical cancer, constituting roughly 60% and 10%, respectively (Colpani et al., 2020). Cancers affecting the cervix, vagina, vulva, as well as anal squamous cell carcinomas are primarily linked to HR HPVs, with HPV 16 being particularly noteworthy in this context (Cornall et al., 2013; Kombe Kombe et al., 2020).
LR HPV Strains
LR HPV types encompass LR-HPV 6, 11, 40, 42, 43, 44, 54, 61, 70, 72, and 81, which are primarily associated with benign genital warts (Colpani et al., 2020). The presence of over 200 common LR HPVs leads to the development of various seemingly benign epithelial lesions, such as genital warts (caused by HPV 6 and 11), common warts (caused by HPV 2, 27, and 57), flat warts (caused by HPV 3 and 10), verrucas or myrmecia (caused by HPV 1), and numerous other skin lesions (Egawa and Doorbar, 2017; Kombe Kombe et al., 2020). Generally, the immune system tends to eliminate LR HPV types more rapidly compared with HR types (Orlando et al., 2012).
In the general populace, LR HPV infection is not generally regarded as a significant contributor to malignant transformation, and the connection between LR HPV and cancer is uncertain, except in cases involving individuals with weakened immune systems and specific genetic backgrounds (Egawa and Doorbar, 2017). However, research has demonstrated that the risks of developing cancers associated with HPV are elevated in diseases induced by LR HPV types (Colpani et al., 2020).
Studies indicates that the E7 protein found in HPV6 and HPV11 exhibits a reduced binding affinity for pRb compared with the E7 protein found in HR HPV types. This difference in binding affinity may be one of the factors contributing to the lower efficiency of malignant transformation by HPV6 and HPV11 (Ivancic et al., 2020). Furthermore, in HR strains, E7 is responsible for triggering pRb degradation facilitated by the proteasome, thereby releasing the transcription factor E2F to drive cells into the S phase. In addition, exclusively HR E6 has the capability to facilitate the degradation of p53 through an E6AP-dependent mechanism, which compromises the host cell’s capacity to effectively complete cell cycle checkpoints (Ivancic et al., 2020). Both HR and LR HPV strains possess the ability of E6 to interact with p300/CBP, preventing the process of acetylating p53. However, HR E6 exhibits a stronger binding affinity to p300/CBP compared with LR E6 (Ivancic et al., 2020).
Immune Interactions
Around 80% of individuals experience anogenital HPV infection at some stage in their lives. However, these infections are predominantly cleared by the immune system of the host, a process that may extend over 1–2 years from the initial exposure to the virus. Only a small percentage of individuals who have contracted the virus go on to develop HPV-related tumors (Lo Cigno et al., 2020). The persistent presence of E6 and E7 in tumors implies their involvement in both the development and sustenance of these cancerous growths (Kajitani and Schwartz, 2022). The activity of E6 and E7 likely exerts significant control over cell proliferation, a function typically governed by proteins such as p53 and pRb. This control allows for the buildup of genetic mutations and abnormalities in chromosomes, ultimately contributing to the onset of cancer (Kajitani and Schwartz, 2022).
The latency phase of HPV infection typically spans about 10 years, during which the host’s immune response significantly influences the advancement or decline of cervical HR HPV infection (Song et al., 2015). The virus is naturally cleared by a robust and effective immune response. Conversely, a weakened immune response often initiates the pathological process, leading to the development of CIN and eventually carcinogenesis (Song et al., 2015).
Infection with HR HPV prompts the mobilization of immune cells to the dermal layer (Song et al., 2015). T lymphocytes, macrophages, KCs, Langerhans cells (LCs), natural killer (NK) cells, dendritic cells (DCs), and B lymphocytes within the squamous epidermis all contribute significantly to orchestrating the immune response to infection (Aghbash et al., 2022) (Fig. 4). HR HPV infection has the potential to enhance immune system resistance, but it may also create a microenvironment that is susceptible to subsequent infections, thereby facilitating the progression of CIN (Aghbash et al., 2022). The proposed and verified mechanisms include the following: To begin with, HR HPV remains latent for an extended period, and its replication and assembly do not induce cell lysis or cytopathic host cell death (Song et al., 2015). Next, HR-HPV interferes with the synthesis of interferon (IFN) by means of the E6 and E7 oncoproteins, which disrupt IFN signaling pathways (Song et al., 2015). Moreover, HR-HPV infection prompts the infiltration of regulatory T cells (Tregs) and the release of interleukin (IL)-10, along with the production of transforming growth factor β (Song et al., 2015). Furthermore, infected cells exhibit reduced expression of major histocompatibility complex class I (MHC1), compromising the function of cytotoxic T lymphocytes (CTLs) (Song et al., 2015). Finally, the accumulation of nonfunctional CD4 and CD8 T lymphocytes can be induced in stage II/III CINs (Song et al., 2015).
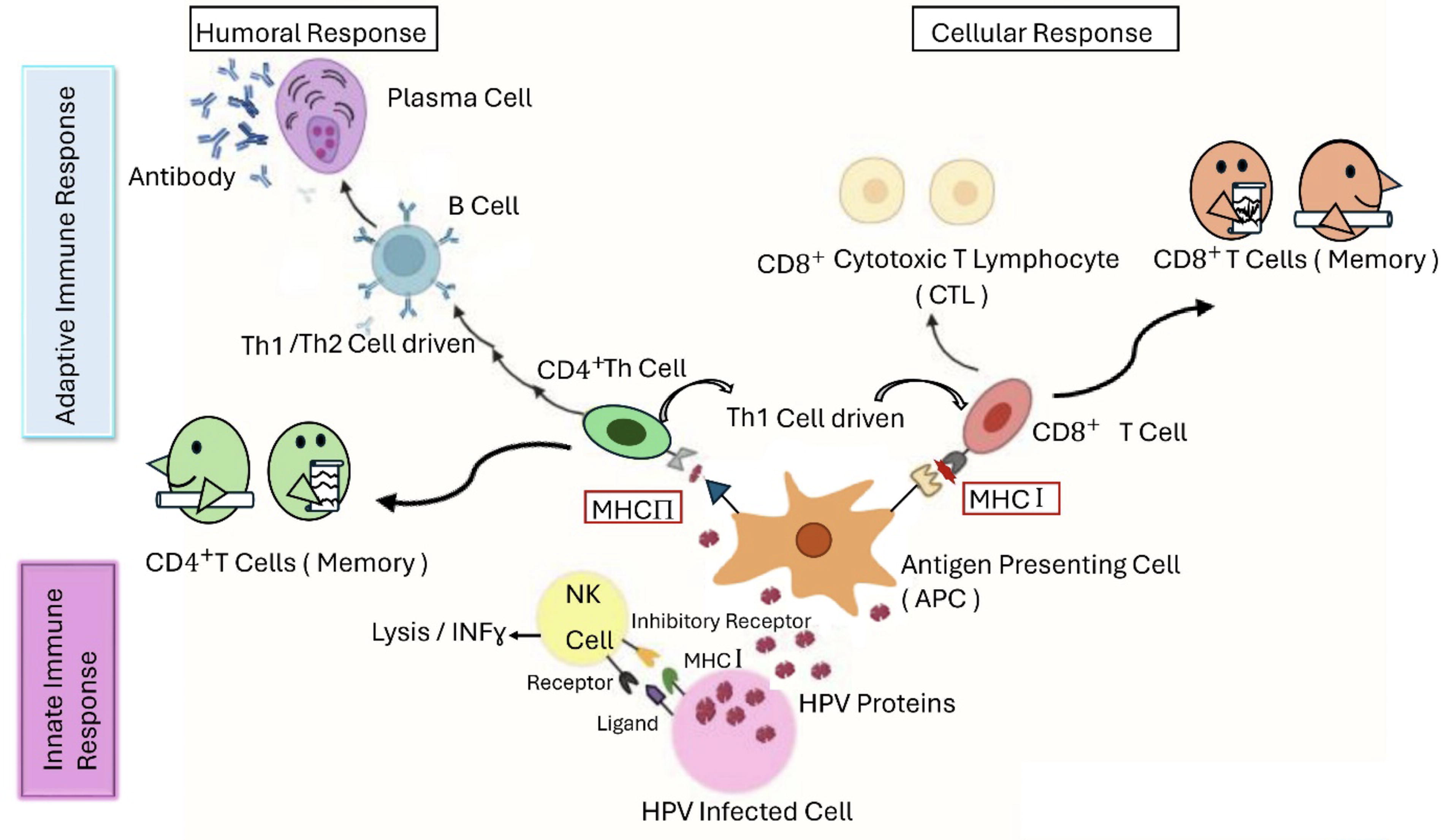
Schematic illustration of innate and adaptive immune responses against HPV. During the initial phase of infection, innate immune cells, including natural killer cells, macrophages, and dendritic cells, initiate the immune response by neutralizing HPV and releasing cytokines. Particularly, dendritic cells function as antigen-presenting cells following virion destruction. During intracellular processes, they present specific peptides on their surface, which are then recognized by the major histocompatibility complex I and II molecules. This process ultimately triggers both cellular and humoral immune responses, which persist for an extended duration.
Simultaneously, the oncogenic stimuli initiated by HPV oncoproteins compel host cells to enhance their innate immune response against the virus (Lo Cigno et al., 2020). Nevertheless, HPVs have developed tactics to circumvent the host’s antiviral immune response, enabling them to successfully finish their viral life cycle and endure within the host cell (Lo Cigno et al., 2020).
The role of innate immunity and its evasion in HPV infection
The initial defense against HPV infection in its early stages is provided by the innate immune response of the host (Amador-Molina et al., 2013; Nakkala et al., 2021). The successful activation of the immune response marks a critical juncture that determines whether the virus will be cleared or persist in the host (Le Tortorec et al., 2020). An improper release of proinflammatory agents and the initiation of a persistent inflammatory reaction can potentially play a role in cancer progression, a phenomenon commonly observed in tumors (Le Tortorec et al., 2020) (Fig. 4).
Keratinocytes, alongside innate immune cells, considered nonprofessional immune cells, are also susceptible to HPV infection and exhibit pattern recognition receptors (PRRs) (Andrei et al., 2023). These receptors have the ability to detect microbial pathogens or signals of damage, referred to as pathogen-associated molecular patterns (PAMPs) or damage-associated molecular patterns (Andrei et al., 2023). PRRs encompass a variety of types, such as nucleotide-binding oligomerization domain-like receptors (NLRs), toll-like receptors (TLRs), cytosolic DNA sensors, and retinoic acid-inducible gene I-like receptors (Andrei et al., 2023).
PRRs recognize microbial molecules, specifically nucleic acids that have the potential to form aggregates during viral replication (Li and Wu, 2021; Okude et al., 2020). Elevated levels of TLR3, TLR7, TLR8, and TLR9 expression, known for their ability to detect viral nucleic acids, correlate with clearance of HPV and has been utilized as an indicator of clearance in women with HPV16 infection, as demonstrated in prior research (Okude et al., 2020; Mohan et al., 2022). The collaboration between TLRs and an HPV16 E6-specific effector response is notably linked to the clearance of the virus (Chitsike and Duerksen-Hughes, 2020; Okude et al., 2020). Nonetheless, certain HR HPV types have been demonstrated to have the capability to undermine the effectiveness of the innate immune response. They not only downregulate the expression of certain TLRs but also disrupt critical pathways that involve transcription factors such as nuclear factor kappa B (NF-kB) and IFN regulatory factor 3 (IRF3). These actions aid the virus in evading the immune system and persisting in the host (Okude et al., 2020).
Articles have reported the existence of variations in genes related to innate immunity in the context of HPV infection, particularly in genes associated with infection sensors and ILs (Nunes et al., 2018). Despite TLR9 being acknowledged as a DNA sensor, it appears that variations in TLR9 are not linked to either the clearance or persistence of the virus (Nunes et al., 2018).
Conversely, variations in other innate immune genes, including IL-18, IL-1b, NLR3, and NLR1, have been linked to HPV infection and its persistent presence (Nunes et al., 2018). Interferon gamma-inducible protein 16 (IFI16), an additional sensor for extrinsic DNA, has been demonstrated to regulate HPV18 replication and transcription by inducing changes in chromatin structure. This action reduces viral load and facilitates viral clearance (Brezgin et al., 2021). The APOBEC protein family, referred to as apolipoprotein B mRNA editing enzyme catalytic polypeptide-like proteins, represents another antiviral mechanism. Members of this protein family are involved in modifying viral genomes, serving as a preventive measure against HPV infection (Brezgin et al., 2021).
Interestingly, extended activation of APOBEC during HPV infection can elevate genome mutagenesis, potentially contributing to the development cancers associated with HPV (Porter and Marra, 2022). The HPV success in establishing infection and persisting within the host hinges on the utilization of multiple strategies to avoid detection by the host’s innate immune system (Porter and Marra, 2022). These mechanisms encompass the reduction of the sensor expression, the blocking of signal transduction pathways, and the interference with the activation of transcription factors, among other strategies (Porter and Marra, 2022).
These strategies, which involve the adjustment of cytokines and chemokines, diminution in the expression of adhesion molecules, hindrance of antigen presentation, suppression of IFN pathways, are primarily orchestrated by the E6 and E7 oncoproteins (Yuan et al., 2021). For instance, HR HPV has the capability to hinder production of cytokine by impeding NF-kB activation in response to stimulation of the innate immune system (Yuan et al., 2021). Viral oncoproteins possess the capability to elevate the levels of IFN-related developmental regulator 1 (IFRD1). IFRD1 plays a role in attracting histone deacetylase 3 (HDAC3) and HDAC1, resulting in the deacetylation of NF-kB and suppressing its responsiveness to immunological signals (Yuan et al., 2021).
Moreover, HR HPV has the ability to obstruct signaling through PRRs by stimulating the expression of ubiquitin C-terminal hydrolase L1 (UCHL1). UCHL1 functions to impede the activation of IRF3 and NF-kB, both being transcription factors responsible for promoting the production of proinflammatory cytokines and chemokines (Nunes et al., 2018). Furthermore, HR HPV has the capacity to diminish the expression of inflammatory elements and other downstream mediators associated with PRRs, including the IL-1b network genes. This interference not only affects innate immunity but also modulates the onset of the adaptive response, a process further regulated by IL-1b (Nunes et al., 2018). HPV16 E6 has been demonstrated to directly influence IL-1b by interacting with and recruiting the E6AP ubiquitin-protein ligase. This interaction leads to the breakdown of cytokines via the proteasome system (Nunes et al., 2018).
Other HPV proteins can also contribute to immune evasion. For instance, HPV16 E2 is capable of influencing the expression of 92 genes related to the innate immune response, such as the stimulator of IFN genes, INF-stimulated genes, and IFN-kappa (Nunes et al., 2018).
Another tactic used by HPV to escape the immune system involves impairing the IFN-mediated response (Huérfano et al., 2022). Viral oncoproteins diminish the production of IFN in keratinocytes, hinder the phosphorylation of mediators in the IFN pathway, and disrupt signaling via IRFs (Huérfano et al., 2022). For instance, cervical tissues and cancer cell lines exhibit decreased expression of IRF1, and the diminished phosphorylation of IRF3 is linked to HR HPV infection (Huérfano et al., 2022). Hence, IFN plays a pivotal role as a part of the innate immune response against HR HPV (Huérfano et al., 2022).
Lastly, it is crucial to emphasize the ability of HPV to disrupt the movement and attachment of innate immune cells and to influence the determination of cell phenotypes (Brown et al., 2023). For instance, HPV infection can compromise the functionality of NK cells, macrophages, and antigen-presenting cells (APCs) (Brown et al., 2023). The DC population, professional APCs responsible for initiating T cell-mediated immune responses, is diminished in CINs (Brown et al., 2023). Moreover, DCs from HR HPV-positive individuals exhibit increased levels of programmed death-ligand 1 (PD-L1), which interacts with programmed cell death protein 1 (PD-1) to induce T cell exhaustion, compared with DCs from those who are negative for HR HPV (Brown et al., 2023). Therefore, a malfunctioning innate immune response can play a role in the persistence of HPV, a factor recognized in the transformation of cells and the progression of tumors induced by HPV infection (Brown et al., 2023).
Role of keratinocytes in HPV infection
HPV’s natural target cells are keratinocytes found in the basal, proliferative layer of the stratified squamous epithelium. These keratinocytes are present in the skin, as well as in mucosal sites such as the anogenital and upper respiratory tracts (Lo Cigno et al., 2020). Keratinocytes have the capacity to recognize a broad spectrum of PAMPs and can react by releasing various cytokines and chemokines (Richards et al., 2015). Subsequently, these chemokines draw specialized immune cells to the infection site and trigger an immune response (Richards et al., 2015). Keratinocytes are equipped with several PRRs, such as TLRs, that are proficient at identifying viral pathogens and stimulating the innate immune response (Richards et al., 2015). TLRs exhibit responses to a variety of viral PAMPs, including double-stranded RNA (recognized by TLR3), single-stranded RNA or imidazoquinoline compounds (recognized by TLR7/TLR8), and hypomethylated DNA (recognized by TLR9) (Richards et al., 2015).
Despite keratinocytes expressing several PRRs, HPVs have evolved mechanisms to create a permissive cellular environment that is unresponsive, allowing for viral replication, persistence, and tumorigenesis (Lo Cigno et al., 2020). Persistent infection by HR HPV genotypes and the disruption of regular viral gene expression, resulting in the sustained overexpression of E6 and E7, are proposed as shared factors contributing to the onset of cervical cancer (Lo Cigno et al., 2020). It is commonly acknowledged that the integration of HPV viral DNA into the host genome occurs frequently, particularly in cases of persistent HPV infection caused by HR genotypes (Lo Cigno et al., 2020). Consequently, this occurrence leads to the stabilization of E6 and E7 transcription, providing a favorable environment for the growth of host cells (Lo Cigno et al., 2020).
Role of DCs in HPV infection
DCs are essential components of the immune system, serving to initiate and regulate immune responses (Nikmanesh et al., 2020; Nikmanesh et al., 2019). The central role of DCs involves recognizing foreign antigens and presenting them to T cells, thereby triggering an immune response (Nikmanesh et al., 2020; Nikmanesh et al., 2019). Upon stimulation by injury or inflammatory signals, DCs move to the adjacent lymph nodes and prompt the differentiation of T cells, thereby triggering an immune response. Hence, the movement of DCs to the nearby lymph nodes is crucial for orchestrating immunity against tumors (Bošnjak et al., 2022) (Fig. 4).
There are reports indicating a notable increase in the count of CD83-positive DCs in CIN, whereas a marked decrease is observed in adjacent lymph nodes (Kara et al., 2009). Alterations in DCs could potentially aid tumor cells in evading the immune system (Tucci et al., 2019). Hence, understanding the mechanisms behind this occurrence is of utmost importance (Tucci et al., 2019).
Prostaglandin E2 (PGE2) is a product induced by cyclooxygenase (COX) from arachidonic acid, which is released from membrane phospholipids. PGE2 plays a role in regulating numerous pathological and physiological processes (Tsuge et al., 2019).
Several investigations have established a close association between PGE2 and the progression of diverse malignant lesions (Tsuge et al., 2019). In addition, the expression of PGE2 is elevated in cervical lesions in comparison with normal tissues (Tsuge et al., 2019). Moreover, some studies have shown that the migration of DCs is regulated by PGE2 (Tsuge et al., 2019). Therefore, PGE2 may be linked to the changes observed in DCs in cervical lesions (Tsuge et al., 2019).
It has been reported that E6, specifically one of its effects on DCs, diminishes the migration of LCs and their progenitors (Doebel et al., 2017). In addition, E6 regulates the transcription of COX-2 (Subbaramaiah and Dannenberg, 2007). The COX pathway is linked to the generation of biologically active prostanoids, which include PGE2 (Mazaleuskaya and Ricciotti, 2020). Hence, it is believed that the modulation of E6 impacts the production of PGE2, subsequently influencing the migration of DCs within cervical lesions (Huang et al., 2020).
DCs are crucial adaptive immune cells that recognize distinctive pathogen patterns through TLRs (Song et al., 2015). DCs initiate immune responses by displaying antigens to T cells using MHC molecules (Song et al., 2015). Elevated levels of stromal DCs are typically associated with a greater likelihood of infection regression (Song et al., 2015). Immature DCs, often in conjunction with HR HPV infection, typically hinder the activation of an effective immune response by CTLs or killer T cells, leading to the promotion of lesion growth (Song et al., 2015). Patients with cervical cancer typically display a very low presence or total lack of operational DCs (Song et al., 2015). The full maturation of monocytes into fully operational DCs can be inhibited by cells expressing HR-HPV E6 or cancer cells that are HR-HPV positive (Song et al., 2015). A reduced percentage of plasmacytoid DCs (pDCs) in the bloodstream is significantly linked to chronic HR HPV infection (Song et al., 2015). Furthermore, diminished levels of CD80 and CD86 expression in DCs are positively correlated with higher CIN grades in patients who are HR-HPV positive (Song et al., 2015).
The impairment of DC activation in HR HPV infection is thought to be influenced by the activation of the PD-1/PD-L1 (CD279/CD274) pathway (Song et al., 2015). The activation of PD-1/PD-L1 is linked to the weakened immune response mediated by DCs against HR HPV infection, establishing an immune-tolerant microenvironment, that facilitates chronic infection and the progression of CIN (Song et al., 2015).
PDCs serve a critical function in linking the innate and adaptive branches of the immune response in the immune system (Han et al., 2018). pDCs secrete IFN type I at levels 10–100 times higher than that observed in other cell types, such as myeloid DCs (Han et al., 2018). HPV 16 virus-like particles (VLPs) can stimulate the generation of type I IFN in pDCs (Han et al., 2018). Simultaneously, the innate immune response provoked by infection of HPV VLPs is relatively restricted, frequently insufficient to eradicate the virus, leading to chronic infection (Han et al., 2018). The E6 and E7 oncoproteins, which are major players in disrupting the cell cycle, apoptosis, and adaptive immune surveillance, are regarded as specific targets for immunotherapy (Han et al., 2018). Prior research has explored the use of DCs loaded with HPV 16/18 E7 antigen as a cellular tumor vaccine (Viscidi et al., 2023).
During the maturation of pDCs, the essential involvement of TLRs is observed in enhancing the expression and migration of chemokines (Viscidi et al., 2023). TLRs are receptors of the innate immune system that identify conserved patterns on microbes, initiating inflammatory signals (Viscidi et al., 2023). TLRs are limited to TLR7 and TLR9 in both human and mouse pDCs (Viscidi et al., 2023).
Role of NK cells in HPV infection
NK cells, part of the innate immune system, are found in the cervix and possess the ability to eliminate virus-infected and transformed cells. They achieve this by directly recognizing surface ligands, which are often downregulated MHCI molecules, and NK cells are resistant to killing by CTLs (Zhou et al., 2019). NK cells have been observed to appear in the early stages of HPV-infected lesions (Zhang et al., 2019) (Fig. 4). NK cells exert their function through two pathways that do not necessitate prior sensitization: granule-dependent cytotoxicity and the apoptosis pathway in the target cells (Amador-Molina et al., 2013). Tumor cells use strategies to evade NK cell attacks, whereas viruses such as HPV have inherent mechanisms that can activate NK cells, resulting in the depletion of their membrane receptors. This can result in NK cell dysfunction and, consequently, may contribute to carcinogenesis (Amador-Molina et al., 2013).
The effectiveness of NK cells is intricately managed through an equilibrium between inhibitory and activating receptors (Amador-Molina et al., 2013). Individuals with cervical cancer and precursor lesions exhibit diminished levels of NKp30, NKp44, and NKp46 receptors in their NK cells. This reduction in receptor levels is linked to the diminished cytotoxic activity of NK cells in these individuals (Amador-Molina et al., 2013). Another crucial receptor in NK cells associated with cytotoxicity is NKG2D. It plays a role in cell lysis by interacting with major histocompatibility complex class I-related chain A (MICA) proteins (Amador-Molina et al., 2013). NKG2D and MICA undergo changes in the context of HPV infection [49]. In an NK cell line (NKL), the downregulation of the NKG2D receptor and the elevation of free MICA levels in the serum were observed in correlation with HPV persistence and the development of cervical cancer (Amador-Molina et al., 2013).
HPV can also evade NK cell cytotoxic mechanisms by elevating the levels of the immunosuppressive enzyme indoleamine-2,3-dioxygenase (IDO). The expression of IDO is associated with tumor cells escaping from immune surveillance and impairs NK cell function in cervical lesions that are positive for HPV (Amador-Molina et al., 2013; Zhou et al., 2019). IDO expression in the tumor environment may be heightened by activated macrophages, APCs, and tumor cells, which can increase the susceptibility of cervical cancer cells to NK cells (Amador-Molina et al., 2013; Zhou et al., 2019).
Role of NKT cells in HPV infection
NKT cells, also known as natural killer T cells, represent a subset of T lymphocytes that coexpress markers related to both conventional NK cells and T cells, such as NK1.1 and NKp46 (Nelson et al., 2021). NKT cells participate significantly in monitoring and modulating antitumor immunity, by recognizing and responding to lipid and glycolipid antigens expressed by tumor cells (Nelson et al., 2021). NKT cells with a semi-invariant TCR are the best-characterized subset, and they release high amounts of proinflammatory cytokines upon detecting endogenous and exogenous glycolipids presented by MHCI-like molecules known as CD1d molecules. These CD1d molecules showcase lipid antigens on the surface of both APCs and tumor cells (Nelson et al., 2021; Zhou et al., 2019).
HPV 16-infected cells often exhibit reduced levels of CD1d, both in vivo and in vitro. This decline is attributed to the expression of HPV16 E5. This decrease in CD1d expression could potentially assist HPV-infected cells in evading protective responses from NKT cells (Starska-Kowarska, 2023). In investigations of high-grade lesions associated with HPV infection, there has been noted an accumulation of NKT cells producing IFN-γ. This observation suggests that NKT cells may contribute to the immune response against lesions induced by HPV (Starska-Kowarska, 2023). This paradoxical role of IFN-γ-producing NKT cells as immunosuppressive in HPV-related carcinogenesis is an interesting observation and highlights the intricacies of the immune response to HPV infection (Starska-Kowarska, 2023).
Delving deeper into the source and roles of these NKT cells, researchers observed the presence of myeloid cells characterized by CD11c+ F4/80hi CD1dhi in the E7 transgenic skin (Zhou et al., 2019). Considering the diminished CD1d expression in keratinocytes infected with HPV, there is a likelihood that these CD1dhi myeloid cells serve as an alternate CD1d source, potentially resulting in the stimulation of immunosuppressive NKT cells within the local tissue milieu (Zhou et al., 2019). Furthermore, IFN-γ is recognized as a stimulator of IDO1 expression (Zhou et al., 2019). Hence, the heightened expression of IDO1 in the tumor environment linked to HPV could be stimulated by NKT cells producing IFN-γ, creating a positive feedback loop of immunosuppression (Zhou et al., 2019).
The role of adaptive immunity and its evasion in HPV infection
The HPV life cycle occurs entirely within the epithelial cells, and virions are only produced by the fully differentiated upper layer of the skin (Hewavisenti et al., 2023). Consequently, there is no cytolysis or viremia induced by the virus, restricting the exposure of HPV to systemic immune responses (Hewavisenti et al., 2023). Nonetheless, research has demonstrated that the activation of T cell responses in the host is necessary for the clearance of HPV-infected cells (Hewavisenti et al., 2023). A robust correlation exists between the regression rate in cervical precancerous lesions and the presence of cytotoxic T cells expressing granzyme B within the epithelium (Borella et al., 2020).
According to a recent investigation, LCs obtained from women with chronic HPV16 infection have the capacity to express HPV antigens and stimulate HPV16-specific CD8+ T cells (Borella et al., 2020). Moreover, in the latest mouse PV (Mus musculus PV 1 or MmuPV1) model, Handisurya et al. showed that successful MmuPV1 infection and the development of papillomas depend on the functions of both CD4+ and CD8+ T cells (Fig. 4). Resistance to productive MmuPV infection was found in C57BL/6 mice lacking either CD4+ or CD8+ T cells. However, in immunocompetent mice, removal of CD4+ and CD8+ T cells resulted in infection and the formation of papillomas (Borella et al., 2020). Furthermore, the immune suppression caused by ultraviolet-B irradiation increases susceptibility to MmuPV1 infection in mice, resulting in eventual squamous cell carcinoma formation (Westrich et al., 2017). Collectively, it has been demonstrated that host’s ability to clear HPV infection relies significantly on T cell-mediated immune responses (Westrich et al., 2017). The current HPV vaccines, their immunogenicity, and effectiveness provide support for the concept that antibody-mediated immunological memory can hinder HPV infection as well (Westrich et al., 2017). Nonetheless, the levels of antibodies produced in response to natural HPV infection tend to be insufficient to confer protective effects. This observation indicates that HPV has evolved effective strategies to evade the immune response mediated by host antibodies in the course of natural infection (Westrich et al., 2017).
Role of humoral response (B cells) in HPV infection
B cells are essential in the humoral immune response as they generate antibodies capable of neutralizing and opsonizing viral agents (Deligeoroglou et al., 2013). In the human immune system, different B cell subsets exist, encompassing plasma cells, immature B cells, memory B cells, B cell progenitors, and immunosuppressive or regulatory B cells (B-regs) (Kim et al., 2020). The establishment of germinal centers is a distinctive feature of B cell-mediated adaptive immunity, essential for the effective affinity maturation of B cells and the generation of diverse antibodies (Kim et al., 2020) (Fig. 4).
The activation of humoral immunity involves APCs and follows the Th2 cytokine pattern. It relies on the assistance of CD4 helper T cells, which support B cell maturation and the production of antibodies targeting specific epitopes (Kennedy and Celis, 2008). Anti-HPV antibodies primarily target the L1 capsid protein. However, there are also weaker antibodies targeting E2, E6, E7, and L2 (Kennedy and Celis, 2008). The predominant class of antibodies in response to these antigens is IgG1, which is a typical immune reaction against viral elements (Kennedy and Celis, 2008). There are two categories of neutralizing L1 antibodies: the initial type prevents attachment to the cell surface, whereas the second type obstructs attachment to the basement membrane (Kennedy and Celis, 2008). The two categories of neutralizing L1 antibodies appear to inhibit viral internalization whether through directly binding to the virus or by obstructing essential conformational changes (Kennedy and Celis, 2008).
Around 8–9 months following a natural HPV infection, seroconversion and the presence of neutralizing antibodies can be detected, although they are often present in low levels and may not be evident in all females (Deligeoroglou et al., 2013). Neutralizing antibodies targeting the L1 capsid protein, specifically those in the second category preventing attachment to the basement membrane, are more abundant in response to VLPs used in prophylactic vaccines than in natural infection, and these vaccine-induced antibodies can persist at high levels in long-term studies (Deligeoroglou et al., 2013; Olczak and Roden, 2020).
Role of cell-mediated immunity (T cells) in HPV infection
The innate immune response is triggered upon the introduction of a pathogen (Meyer et al., 2014). The pathogen is enclosed within the DC, subsequently transforming it into an APC (Meyer et al., 2014). This cell conveys the antigen to the lymph nodes, subsequently initiating the adaptive immune response (Meyer et al., 2014). The transformation of naive cells into cytotoxic T cells (CD8+) and T helper cells (CD4+) can trigger cellular and humoral immune responses (Meyer et al., 2014) (Fig. 4).
There are two presentation pathways that initiate the adaptive immune response: cytotoxic T cells (CD8+) are stimulated via MHCI, whereas T helper cells (CD4+) are activated through MHCII (Cho et al., 2021; Raskov et al., 2021). In addition, an inflammatory response and the generation of cytokines are essential components of the immune reaction (Cho et al., 2021; Soto-Heredero et al., 2020). HLA, also known as human leukocyte antigen, represents another vital component [82]. The presence of HLA alleles in cancerous cells is indispensable for the effectiveness of cytotoxic T cells (CD8+) (Cho et al., 2021; Raskov et al., 2021). CD8+ plays a pivotal role in cancer-specific immunity, as it identifies antigens with the help of MHC/HLA class I (Cho et al., 2021; Raskov et al., 2021).
HPVs possess a range of characteristics that allow them to evade the innate immune response and postpone the adaptive immune response activation (Ko et al., 2022). These traits hinder the inflammatory reaction, the synthesis of proinflammatory cytokines, and the stimulation of DCs, all of which are necessary to initiate a cell-mediated immune (CMI) response (Ko et al., 2022). HPV16 E5 has the capacity to suppress CD8+ cells by reducing the expression of MHC/HLA class I (Ko et al., 2022). HPVs also reduce the production of antiviral cytokines in keratinocytes, and the viral early proteins, E6 and E7, inhibit the expression of antiviral cytokines (Ko et al., 2022). In summary, if the CMI response is inadequate or unsuccessful, HPV-infected cells may not be eradicated (Ko et al., 2022). Consequently, the virus may persist, and the infection may endure (Ko et al., 2022).
The shift from HR-HPV infection to cervical cancer is fundamentally linked to compromised adaptive immunity (van Bockel and Kelleher, 2023). Research has demonstrated that compromised CD4+ T cells contribute to the development of HPV-associated cancers in individuals with HIV infection (van Bockel and Kelleher, 2023). In these individuals, the inefficient activation of CD4+ T cells due to HR-HPV infection is the element that promotes the advancement of CIN lesions (van Bockel and Kelleher, 2023). The cellular immune response during HR-HPV infection is marked by an imbalance in the ratio of type 1 T-helper cells (Th1) to Th2 cells (van Bockel and Kelleher, 2023). Both intraepithelial and invasive cervical lesions exhibit a distinct Th2 cytokine profile (Schindler et al., 2022). Elevated levels of Th2 cytokines (specifically IL-10) and decreased levels of Th1 cytokines (including IFN-γ, IL-12, IL-2, and tumor necrosis factor-α) have been identified in the cervical secretions of HR-HPV-positive individuals (Schindler et al., 2022). This observation indicates that the diminished Th1 response and heightened Th2 response contribute to the suppression of cellular immunity and the progression of cervical lesions (Schindler et al., 2022). An additional investigation demonstrated a reduction in the synthesis of specific Th2 cytokines during this progression, implying that both the Th1 and Th2 phenotypes, with a particular emphasis on the Th1 phenotype, might be inhibited by HR-HPV infection (Schindler et al., 2022).
One method by which HR-HPV modulates the activation of T cell involves E6 and E7 oncoprotein expression. These oncoproteins have the capability to increase the expression of adhesion molecules, including intercellular adhesion molecule in the infected cells, vascular cell adhesion molecule 1, and E-selectin, which are considered classical immunosuppressive factors (Rasheed, 2020; Song et al., 2015). In summary, the compromised T cell activation and the disrupted balance between Th1 and Th2 responses provide HR-HPV with the capability to evade the human immune defense and contribute to lesion progression (Ojha et al., 2022; Song et al., 2015).
Vaccines
Roughly 60% to 70% of females who become infected with HPV develop a detectable antibody response specific to the HPV L1 capsid protein, characterized by epitopes that become apparent in their serum. This response serves as a less sensitive indicator of the overall exposure to HPV over time (Beachler et al., 2016). In theory, these naturally acquired polyclonal antibodies could potentially provide protection against subsequent infections with the same type of HPV (Beachler et al., 2016). Indeed, this natural HPV immunity can be considered a factor in cost-effectiveness models when assessing the advantages of administering prophylactic HPV vaccines to older individuals (Beachler et al., 2016).
Six licensed prophylactic HPV vaccines are available: three bivalent, two quadrivalent, and one nonavalent vaccine (Williamson, 2023). These include the 2-valent HPV-16/18 AS04-adjuvanted vaccine (AS04-HPV-16/18v; Cervarix, GSK), the 4-valent (4vHPVv) (6,11,16,18), and the 9-valent (9vHPVv) (6,11,16,18,31,33,45,52,58) aluminum hydroxyphosphate sulfate adjuvant vaccines (Gardasil, Merck) and a bivalent HPV vaccine (HPV-16 and HPV-18) (Cecolin, Xiamen Innovax Biotech Co., Ltd., Xiamen, China) (Khan et al., 2023). There is limited information available regarding the newer HPV vaccines. Two recently developed Chinese vaccines underwent testing in a randomized, blinded, noninferiority phase III trial to assess the effectiveness of their innovative four-valent and nine-valent HPV vaccines and were demonstrated to be equally effective as GARDASIL in terms of both immune response and safety (Williamson, 2023). These vaccines use eukaryotic cells or Escherichia coli to produce HPV L1 VLPs (Kheirvari et al., 2023). Research has shown that these vaccines can be effective in preventing not only genital warts but also high-grade vaginal, anal, cervical, and vulvar cancers and precancerous lesions in women. In addition, they have demonstrated efficacy in preventing cancers, precancerous lesions, and genital warts in men (Chaturvedi et al., 2018).
The 2-valent HPV-16/18 vaccine is designed to combat two specific types of HPV, 16 and 18, which collectively contribute to up to 70% of cervical cancers. Moreover, it has demonstrated cross-protection against HPV types 31, 33, and 45, which are the subsequent most prevalent types of HPV associated with cervical cancer (Mehanna et al., 2019). HPV vaccines work via VLPs, which are noninfectious assemblies of the L1 HPV capsid protein. VLPs induce a strong neutralizing antibody response, preventing HPV uptake by cervix basal cells. The vaccines provide subtype-specific protection and cross-protection against other HPV types (Illah and Olaitan, 2023). The existing Gardasil HPV vaccine has proven to elevate the population of NK cells following immunization. This increase is linked to heightened levels of receptors such as NKp30, NKp46, NKG2D, and ILT2 in NK cells. This suggests the involvement of additional pathways, extending beyond the augmentation of neutralizing antibodies, in contributing to the effectiveness of the vaccine (Amador-Molina et al., 2013; Akhatova et al., 2022).
A comprehensive examination of real-world experiences with the 4vHPV vaccine on a global scale demonstrated a reduction of over 90% in HPV 6/11/16/18 infections, a 90% decline in genital warts, a 45% decrease in low-grade cytological cervical abnormalities, and an 85% decrease in high-grade cervical abnormalities among women in the younger age groups who are the primary targets of national immunization programs in nine countries (Maver and Poljak, 2018). The efficacy of HPV vaccines is influenced by several factors, including vaccine coverage, the birth cohorts’ age targeted for vaccination, the implementation of comprehensive programs in older age groups, the time between program commencement and outcome evaluation, and the duration of follow-up. These elements collectively contribute to the overall impact of HPV vaccination programs (Maver and Poljak, 2018). Nevertheless, a documented decline in the prevalence of HPV infections and genital warts among unvaccinated women and men during the vaccine era suggests the possibility of herd protection (Maver and Poljak, 2018).
Up to this point, there has been limited exploration into the impact of vaccination on oral HPV infection (Mehanna et al., 2019; Nielsen et al., 2021). It is crucial to note that all studies have utilized oral rinse for assessment, and there have been no investigations that simultaneously examine HPV prevalence using both oral rinse and tonsil tissue, or evaluate the vaccine’s impact on HPV prevalence in tonsil tissue, which is the primary site associated with oropharyngeal cancer (Mehanna et al., 2019). In addition, there is a lack of research examining the effectiveness of vaccination initiatives in reducing oral HPV prevalence among children, and no studies have explored the protection of men against oral HPV infection through the potential herd effect resulting from a national vaccination program targeting females exclusively (Elit et al., 2022; Mehanna et al., 2019) (Fig. 5).
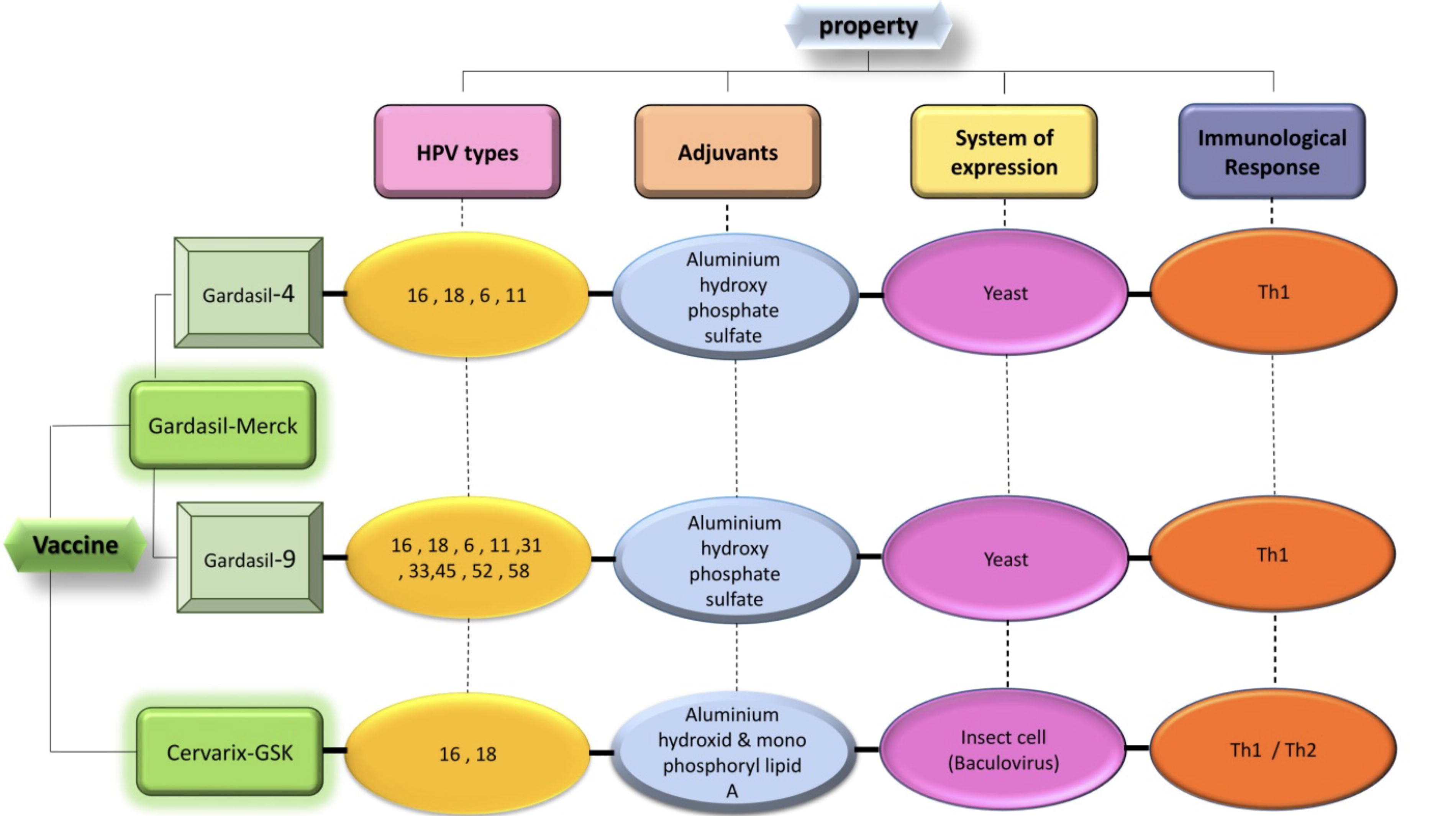
Characteristics and content of HPV prophylactic vaccines.
Common Treatments
There are limited data available regarding the comprehensive treatment of cervical diseases associated with HPV (Kechagias et al., 2022). The main objective of treatment is to relieve symptoms, eliminate wart-infected transformation zones, and decrease the likelihood of subsequent invasive cervical cancer (Kechagias et al., 2022). Managing and treating diseases associated with HPV greatly depend on several factors, such as the particular HPV types involved, the variety of treatment options available, and the stage or progression of the disease (Bosch et al., 2013; Gupta et al., 2018). These critical factors influence the approach taken to effectively manage and treat HPV-related conditions (Bosch et al., 2013).
To address external genital warts resulting from nononcogenic HPVs, the common recommendation is the use of trichloroacetic acid, sinecatechins, podophyllotoxin, and imiquimod at concentrations ranging from 80% to 90% (Sindhuja et al., 2022). Furthermore, in specific cases, other limited therapies such as photodynamic therapy, intralesional/topical IFN, and 5-fluorouracil may be considered for managing external genital warts resulting from nononcogenic HPVs (Gilson et al., 2020) (Table 1).
Suggested Treatments and Limited Strategies to Deal with Low-Risk Papilloma Infections
It is typically recommended to use surgical methods to address cervical precancerous lesions induced by oncogenic HPVs. These methods may include laser therapy, loop electrosurgical excision procedure, electrosurgery (such as cone biopsy or conization), cryosurgery (freezing), as well as excisional procedures with local anesthesia (Lee et al., 2022). Numerous studies have demonstrated that the choice of a specific surgical method has minimal impact on the likelihood of recurrence in managing cervical precancerous lesions attributed to oncogenic HPVs. Therefore, the selection of treatment may be contingent on the accessibility of the method and the physician’s discretion (Cusimano et al., 2019) (Table 2).
Suggested Treatments to Deal with Cervical Precancerous Lesions Caused by High-Risk HPVs
Current treatment approaches for cervical cancer frequently involve a combination of adjuvant or neoadjuvant chemotherapy, radiation therapy, and often a hysterectomy. These strategies aim to effectively manage and treat the disease while considering its stage and extent (Miriyala et al., 2022). Research has indicated that chemotherapy is an important and efficient choice for treating cervical cancer, applicable across a spectrum of stages, ranging from locally advanced to metastatic cases (Gadducci and Cosio, 2020; Miriyala et al., 2022). In the past, cisplatin-based chemotherapy (CRT) was the preferred treatment for women with distant metastases or recurrent cervical cancer (Do et al., 2021). Currently, the widely acknowledged standard treatment for invasive cervical cancer is definitive radiation therapy with concurrent CRT, even though it comes with a somewhat higher risk of recurrence (estimated at 25–40%). This treatment approach is considered the gold standard for managing this condition (Khairkhah et al., 2022). Chemotherapy can be administered either as postoperative adjuvant therapy or as neoadjuvant chemotherapy before surgery in the treatment of cervical cancer. Interestingly, meta-analyses suggest that both approaches yield similar overall survival rates, providing flexibility in the choice of treatment strategy for individual patients (Lorentzen et al., 2022). Recent investigations and analyses suggest that combining various chemotherapy agents, including capecitabine, vinorelbine, pemetrexed, paclitaxel, irinotecan, ifosfamide, topotecan, and S-1, can enhance the efficacy of cervical cancer treatment. The goal of this combined therapy approach is to enhance outcomes for individuals diagnosed with cervical cancer (Duenas-Gonzalez and Gonzalez-Fierro, 2019). The choice of these drugs can be determined according to the specific condition of each patient (Duenas-Gonzalez and Gonzalez-Fierro, 2019).
Hysterectomy is a surgical intervention that entails the extraction of the uterus (Kumar and Vijan, 2021). Radical hysterectomy, a procedure that encompasses the removal of the uterus along with the parametrium and the upper part of the vagina, has been demonstrated to be the most efficient approach (Kumar and Vijan, 2021). Nonetheless, it is crucial to emphasize that a hysterectomy can have adverse effects on fertility and may result in physical and psychological health challenges that can affect a patient’s overall quality of life (Elsaied et al., 2020) (Table 3).
Suggested Treatment Strategies to Deal with Cervical Cancer
Current research has explored the potential advantages of integrating checkpoint inhibitors into existing cervical cancer treatments. Immune checkpoint pathways, particularly cytotoxic lymphocyte antigen 4 and PD-1, can inhibit the body’s immune response and diminish T cell function (Chitsike and Duerksen-Hughes, 2020). Monoclonal antibodies such as pembrolizumab, nivolumab, and ipilimumab have been created and sanctioned by the Food and Drug Administration to specifically target the PD-1 axis for the treatment of cervical cancer (Liu et al., 2019).
Discussion
HPV infection represents a significant global health concern due to its widespread prevalence and association with various cancers. HPV is a circular, double-stranded DNA virus with over 200 identified types, making it the most prevalent sexually transmitted infectious agent worldwide. The HPV life cycle involves infiltration into basal cells of the epidermis, minimal gene expression, and productive replication in differentiated cells. Integration of HPV into the host cell’s DNA contributes to cellular transformation and malignant progression.
Immune interactions play a pivotal role in determining the outcome of HPV infection. Innate immunity, mediated by cells such as macrophages, DCs, and NK cells, serves as the first line of defense. Subsequent adaptive immune responses involving T and B lymphocytes are crucial for eliminating infected cells and generating long-term immunity. However, the balance between clearance and persistence of HPV infection is determined by the effectiveness of the host immune response, highlighting the importance of a well-coordinated immune response.
Prophylactic HPV vaccines have emerged as a cornerstone of prevention efforts, demonstrating efficacy in preventing genital warts, high-grade precancerous lesions, and cancers. Available vaccines offer protection against a range of HPV types, providing options for both females and males. It is essential to note that HPV vaccination does not treat existing infections but prevents future infections and associated health risks.
Effective management and treatment of HPV-related diseases are vital for reducing morbidity and mortality. Surgical interventions, chemotherapy, and radiation therapy play integral roles in addressing precancerous lesions and cervical cancer. In addition, the integration of checkpoint inhibitors into existing treatments represents a promising avenue for enhancing therapeutic outcomes and overcoming immune evasion mechanisms used by HPV.
In conclusion, this comprehensive review provides insights into the genetic, immunological, and clinical aspects of HPV infection and its associated diseases. By elucidating these aspects, evidence-based strategies for the prevention, diagnosis, and treatment can be informed. Collaborative efforts across disciplines are essential for advancing our understanding of HPV and mitigating its impact on global public health.
Conclusion
HPV poses a significant public health challenge, demanding a nuanced understanding of its life cycle, immune response dynamics, and available preventive measures. The advent of prophylactic HPV vaccines has been instrumental in curbing the prevalence of genital warts, cervical cancer, and related diseases, emphasizing their pivotal role in public health strategies. Encouraging vaccination, particularly during adolescence or early adulthood, holds promise in reducing HPV infection rates and averting associated health risks, underscoring the importance of proactive immunization efforts.
Comprehensive approaches that integrate vaccination programs, awareness campaigns, and evidence-based interventions are crucial for effectively addressing HPV impact. By fostering collaboration across disciplines and prioritizing preventative measures, we can advance toward the goal of minimizing the burden of HPV-related diseases on a global scale.
Author Confirmation Statement
Ms. Nikmanesh, Ms. Mirbagheri, Ms. Asadsangabi, Mr. Fattahi, Mr. Safarpour, Mr. Fallahzade Abarghooee, Ms. Shamsdin, and Mr. Akrami are from Shiraz University of Medical Sciences (Shiraz, Iran); Mr. Moravej is from Fasa University of Medical Sciences (Fasa, Iran); Ms. Hosseini is from the University of Kurdistan (Shiraz, Iran); Mr. Saghi is from Larestan University of Medical Sciences (Larestan, Iran) and Mashhad University of Medical Sciences (Mashhad, Iran), and Dr. Nikmanesh is from Shiraz University of Medical Sciences (Shiraz, Iran), all where education and research are the primary functions.
Footnotes
Author Contributions
Conceptualization: Y.N. Data curation: Y.N., S.H., and N.N. Formal analysis: S.H. Funding acquisition: M.R.F. and A.R.S. Investigation: E.F.A., A.M., S.A.S., and H.A. Methodology: Y.N., S.H., and N.N. Project administration: Y.N., M.R.F., and A.R.S. Resources: S.H., N.N., and S.A.Sa. Software: F.M. and S.A.Sa. Supervision: Y.N. and S.H. Validation: E.F.A., A.M., S.A.S., and H.A. Visualization: Y.N. and S.H. Writing—original draft: S.H., N.N., F.M., and K.A. Writing—review and editing: Y.N., S.H., M.R.F., and A.R.S.
Author Disclosure Statement
The authors declare that they have no conflict of interest.
Funding Information
No funding was received for this article.