
Abstract
Aging is physiologically associated with a decline in the function of the immune system and subsequent susceptibility to infections. Interferon-gamma (IFN-γ), a key element in the activation of cellular immunity, plays an important role in defense against virus infections. Decreased levels of IFN-γ in the elderly may explain their increased risk for viral infectious diseases such as COVID-19. There is accumulating evidence that ascorbic acid (vitamin C [VitC]) and α-tocopherol together help improve the function of the immune system in the elderly, control infections, and decrease the treatment duration. A SARS-CoV-2 strain was isolated from a patient and then cultured in the Vero cell line. The isolated and propagated virus was then inactivated using formalin and purified by the column chromatography. The inactivated SARS-CoV-2 was formulated in the Alum adjuvant combined with VitC or α-tocopherol and/or both of them. The vaccines were injected twice to young and aged C57BL/6 mice. Two weeks later, IFN-γ, IL-4, and IL-2 cytokines were assessed using ELISA Kits. Specific IgG and IgG1/IgG2a were assessed by an in-house ELISA. In addition, the expression of PD1 and TERT genes in the spleen tissue of the mice was measured using real-time PCR. IL-4 and IFN-γ cytokines showed a significant increase in both aged and young mice compared with the Alum-based vaccine. In addition, our results exhibited a significant decrease and increase in specific total IgG and the IgG2a/IgG1 ratio, respectively. Furthermore, the vaccine formulated in α-tocopherol + VitC led to decreased PD1 and increased TERT gene expression levels. In conclusion, our results demonstrated that α-tocopherol + VitC formulated in the inactivated SARS-CoV-2 vaccine led to a shift toward Th1, which may be due to their effect on the physiology of cells, especially aged ones and changing their phenotype toward young cells.
Introduction
In December 2019, SARS-CoV-2, a novel coronavirus, emerged in Wuhan, China, the capital of the Chinese province of Hubei, which led to a severe acute respiratory disease officially called COVID-19. The disease rapidly crossed China’s borders, spread all over the world, and caused a pandemic (Chang et al., 2020; Zhou et al., 2020b). SARS-CoV-2 is the seventh coronavirus to infect humans, and it is among the human coronaviruses that led to serious diseases such as SARS and MERS (Cui et al., 2019; Zhu et al., 2020). Angiotensin-converting enzyme 2 (ACE2), which is highly expressed in the lungs and intestinal epithelium, is a receptor for SARS-CoV-2 and is necessary for viral entry (Xu et al., 2020). According to evidence, COVID-19 spreads person-to-person through direct contact or droplets (Carlos et al., 2020; Li et al., 2020). The highest mortality rate appears to occur in the elderly population (Kang and Jung, 2020). Although the fatality rate of COVID-19 is 2.3%, evidence indicates that the mortality rate of COVID-19 is elevated in elderly people by 22% (Garnier‐Crussard et al., 2020; Le Couteur et al., 2020; Leung, 2020; Nicol et al., 2020). Moreover, according to previous reports, 90% and 80% of deaths in patients aged ≥60 years and >70 years have been recorded in Italy and Korea, respectively (Kang and Jung, 2020; Miranda-Massari et al., 2020).
Aging is physiologically accompanied by a decrease in the function of the immune system, in which there is a relationship between alteration in the immune system and susceptibility to infections. In addition, immune responses to the vaccines decline in the elderly so that vaccines are not protective enough in the elderly and aged people are at risk for infectious disease. Cytokine interferon-gamma (IFN-γ), mainly produced by type 1 T cells, has a crucial role in the regulation of cellular immune responses and, thereby, control viral infections. Of note, production of the IFN-γ cytokine decreases in the elderly, which may explain the reason for more susceptibility of aged people to viral infectious diseases, such as influenza and COVID-19. Moreover, previous studies reported that the efficacy of the vaccine in the elderly is lower than others due to the lower level of immune responses (Aspinall et al., 2007; Gustafson et al., 2020; Ouyang et al., 2002; Sandmand et al., 2002; Valiathan et al., 2016). In contrast, there is evidence indicating that the administration of vitamins might improve immune responses. The effects of vitamins, such as ascorbic acid (vitamin C [VitC]), on the improvement of the IFN-γ cytokine and cellular immune responses have been previously reported in a variety of studies (Abobaker et al., 2020; Karlsson et al., 2017b; Kouakanou et al., 2020; Lee and Han, 2018; Wu et al., 2018).
According to the reports, the use of VitC not only leads to decreased mortality in patients with Covid-19, especially those with acute respiratory failure, but also decreases the length of stay of patients in the intensive care unit (ICU) (Marik et al., 2017; Miranda-Massari et al., 2020; Mitchell et al., 2020). α-Tocopherol, the active form of vitamin E, is a fat-soluble antioxidant that protects cells from peroxidation, and VitC regenerates vitamin E in the cell membrane by scavenging free radicals (Adnan, 2020). A wide variety of studies demonstrated the benefits of α-Tocopherol in animal and human models of aging through improved function and modulation of the immune system. In addition, VitC and α-Tocopherol, as biological antioxidants, play roles in protecting cellular macromolecules such as lipids, proteins, and DNA against free radical oxidation and modulate the immune system against infection, stress, and pollution (Carr and Rowe, 2020; Patterson et al., 2021; Thakurta et al., 2012).
Immune senescence or age-related immune system remodeling might be considered the main reason for high vulnerability to infections, especially respiratory infections, such as influenza and COVID-19. In addition, partial immune responses to vaccination (Li et al., 2011; Pawelec, 2018), despite age-oriented modifications in innate and adaptive immune responses against COVID-19 infection, need a more detailed study to be fully understood, and impairment of a physiological regulatory mechanism in older people could be inferred. For instance, senescence-related defects in the type 1 IFN response can lead to an increase in influenza viral reproduction within the cell culture (Kim et al., 2016). Furthermore, older people exhibit a defect in type 1 IFN response to influenza vaccination (Thakar et al., 2015). Consequently, age-related fragile type 1 IFN responses along with direct viral suppression might serve as a crucial innate immune mechanism, presumably leading to the weakness of cell-mediated immune responses and increased vulnerability of aged people to COVID-19 infection with the therapeutic implication (Sallard et al., 2020).
A variety of studies have indicated that elderly people are more vulnerable to COVID-19 infection due to the inefficiency of current SARS-CoV-2 vaccines, highlighting the need for developing a new approach to induce protection in such populations. For this purpose, we developed a SARS-CoV-2-inactivated vaccine formulated in VitC and α-tocopherol and assessed its potency in aged mice. In addition, the expression levels of TERT and PD1 were evaluated to estimate the efficacy of the vaccine. In fact, we hypothesized that VitC and α-tocopherol, with antiaging activity, may increase vaccine potency in aged mice.
Materials and Methods
Virus isolation and propagation
SARS-CoV-2 was isolated from the nasal swab sample of a patient (positive in real-time PCR) from Karaj, Iran. Vero cells were cultured in Dulbecco’s modified Eagle’s medium supplemented with 3% heat-inactivated fetal bovine serum. The SARS-CoV-2 propagation was carried out in biosafety level-III facilities in the Razi Vaccine and Serum Research Institute of Iran, Karaj (Kaffashi et al., 2021).
Virus inactivation and quantification
The isolated SARS-CoV-2 strain was cultivated in the Vero cell line for propagation, and the supernatant of the infected cells was inactivated using formalin (0.03% vol/vol). Following clarification of the cell debris and ultrafiltration, purification was performed using PEG-6000 and column chromatography. The purified inactivated viruses were dialyzed against phosphate-buffered saline (PBS), passed through a 0.22-μm pore size filter, and quantified using the Bradford method. The quantified virus aliquots were then stored at −70°C until use (Kaffashi et al., 2021).
Vaccine formulation
The purified inactivated SARS-CoV-2 virus, after standard quality control tests, was used for vaccine formulation. To this end, 2 µg of the inactivated SARS-CoV-2 virus was mixed with 200 µg of the Alum hydroxide adjuvant for each dose of vaccine (Pasteur Institute of Iran), shaken at 100 RPM for 60 min, and allowed the viral particles to bond to the surface of the Alum gel. Subsequently, this vaccine was further formulated with α-tocopherol (Sigma, USA) and VitC (Sigma, USA). VitC was dissolved in PBS, 1 mg and/or 10 mg of VitC was added to the Alum-formulated vaccine for each dose, and the vaccine was injected after one h of shaking. In addition, α-tocopherol was dissolved in ethanol alcohol and added to the Alum-formulated vaccine at a dose of 1 mg for each dose, and the sample was shaken for one h. For the mixed form, α-tocopherol and VitC were added to the Alum-formulated vaccine. After the final vaccine formulation, 100 µL of each vaccine contained 2 µg of inactivated SARS-CoV-2 virus and the desired amount of α-tocopherol and VitC. It is important to note that a freshly prepared vaccine was used for each immunization.
Mice
Sixteen-to-20-month-old female C57BL/6 mice, aged mice (n = 56), and six-to-eight-week-old female C57BL/6 mice, young mice (n = 56), (Eshraghi et al., 2022) were purchased from Pasteur Institute of Iran (Karaj, Iran). The mice were housed for one week before the experiments, given ad libitum access to food and water, and maintained in 12-h light/12-h dark cycles. The experimental mice were handled by an expert technician in accordance with the Animal Care and Use Protocol of the Ethics Committee’s recommendations in the research of Tehran University of Medical Sciences (Ethics Code IR.TUMS.AEC.1401.202).
Experimental groups and immunization
Experimental young and aged mice were randomly divided into seven groups and each one consisting of 8 mice as below:
Young and aged mice were subcutaneously immunized with 2 µg of each vaccine twice at 14-day interval. Cellular and humoral immune responses were assessed two weeks after the last immunization. In addition, TERT and PD1 gene expression levels were assessed in the spleen tissues of the experimental groups.
Blood sampling and serum separation
Two weeks after the final immunization, blood samples were prepared from the mice and stored at 4°C for 3 h. The serum samples were separated by centrifugation at 10,000 RPM/15 min, and then the samples were stored at −70°C until use.
ELISA for IFN-γ, IL-4, and IL-2 cytokines
Two weeks after the final immunization, the serum levels of IFN-γ, IL-2, and IL-4 cytokines were assessed using quantitative mouse IFN-γ, IL-2, and IL-4 cytokine ELISA Kits (Mabtech, Stockholm, Sweden), according to the manufacturer’s instructions. The ELISA of all cytokines was performed in the presence of cytokine standards, and using the standard curve of each cytokine, the pg/mL of each sample was reported. The IFN-γ/IL-4 ratio was calculated for each mouse by dividing the quantity of IFN-γ in each mouse by the IL-4 ratio.
ELISA for specific total IgG antibodies and specific IgG1/IgG2a isotypes
Specific total IgG antibody against SARS-CoV-2 was determined by an optimized indirect ELISA setup in our laboratory. Briefly, 100 µL of 5 µg/mL of inactivated SARS-CoV-2 in PBS (pH 7.2–7.4) was added to the 96-well ELISA Maxisorp plates (Greiner, Germany) and incubated at 4°C during the night. The plates were then washed with washing buffer (PBS containing 0.1% Tween 20) and blocked for 1 h at 37°C with blocking buffer (2% specific total IgG antibody against SARS-CoV-2 was determined using an optimized indirect ELISA setup in our laboratory). Briefly, 100 µL of 5 µg/mL inactivated SARS-CoV-2 in PBS (pH 7.2–7.4) was added to 96-well ELISA Maxisorp plates (Greiner, Germany) and incubated at 4°C overnight. The plates were then washed with washing buffer (PBS containing 0.1% Tween 20) and blocked for 1 h at 37°C with blocking buffer (2% skim milk in washing buffer). The plates were washed with washing buffer, and 100 µL of 1/25 diluted sera up to 12 serial dilutions was added to each well and incubated at 37°C for 90 min. The wells were washed five times with washing buffer and incubated for 2 h with 100 µL of a 1/8,000 dilution of antimouse conjugated to HRP (Razirad, Iran). The wells were washed five times and incubated for 30 min with 100 µL TMB substrate in the dark. The reaction was stopped by adding 100 µL 2N H2SO4, and the color density was measured at A450 nm using an ELISA immunosorbent assay (ELISA) plate reader (AWARENESS TECHNOLOGY, USA). In addition, detection of specific IgG1 and IgG2a subclasses was performed using goat antimouse IgG1 and IgG2a secondary antibodies (Sigma, USA), according to the manufacturer’s instructions.
PD1 and TERT gene expression in spleen tissue
The expression of PD1 and TERT genes in the spleen tissue of experimental mice was measured using real-time PCR. Briefly, RNA was extracted from spleen tissue samples of aged and young mice using the TRIzol method. The cDNA was synthesized using a RevertAidTM First Strand cDNA Synthesis Kit (Ampliqon, Denmark). The reaction contained 5x solution buffer (4 μL), RNA (10 μL), random hexamer primer (1 μL), deoxynucleotide triphosphate mixture (1 μL), reverse transcriptase (1 μL), and double-distilled water (3 μL) to obtain a final volume of 20 μL for cDNA synthesis. Finally, real-time PCR was performed using primers (Table 1) for the target genes TERT, PD1, and β-actin as an internal control gene. The temperature considered for each gene was as follows: 55°C Tm, Time: 15 s, TERT: 57°C Tm, Time: 15 s, PD1: 54°C Tm, Time: 15 s. Finally, ΔΔCT was used to analyze gene expression.
Table of forward and reverse designed primers
Statistical analyses
Data are presented as the mean ± standard deviation of each experiment. Statistical significance of differences was analyzed using two-way ANOVA followed by the Tukey test for specific IgG responses (GraphPad Prism v 6.01 software, La Jolla, CA). In addition, statistical analyses of IgG1 and IgG2a isotypes, IFN-γ, IL-2, and IL-4 cytokines, and the gene expression assay were carried out using the Mann–Whitney U test. In all cases, p values less than 0.05 was considered significant.
Results
IFN-γ cytokine in aged population
The results of the IFN-γ cytokine assay in the aged experimental groups demonstrated that mice immunized with inactivated SARS-CoV-2-Alum + α-tocopherol + VitC1mg and inactivated SARS-CoV-2-Alum + α-tocopherol + VitC10mg showed a 47.85% and 76.11% increase in IFN-γ levels, respectively, compared with the SARS-CoV-2-Alum group (p = 0.5338 and p = 0.2949, respectively). In addition, mice immunized with SARS-CoV-2-Alum + VitC10mg showed a significant increase in the IFN-γ level compared with the SARS-CoV-2-Alum group (p = 0.0260), whereas the SARS-CoV-2-Alum + VitC1mg group showed a borderline increase in the IFN-γ level compared with the SARS-CoV-2-Alum group (p = 0.0513). Furthermore, no significant effect on the IFN-γ level was observed in mice immunized with SARS-CoV-2-Alum + α-tocopherol + VitC1mg and inactivated SARS-CoV-2-Alum + α-tocopherol + VitC10mg compared with the SARS-CoV-2-Alum + VitC1mg and SARS-CoV-2-Alum + VitC10mg groups (p > 0.2086) (Fig. 1A).

IFN-γ in young population
Results from the IFN-γ cytokine assay in the young groups revealed that mice immunized with inactivated SARS-CoV-2-Alum + α-tocopherol + VitC1mg and inactivated SARS-CoV-2-Alum + α-tocopherol + VitC10mg led to a 37.60% and 59.04% increase in IFN-γ levels compared with the SARS-CoV-2-Alum group (p = 0.7789 and p = 0.5358, respectively). No significant differences in IFN-γ levels were observed among mice immunized with SARS-CoV-2-Alum + α-tocopherol + VitC1mg and inactivated SARS-CoV-2-Alum + α-tocopherol + VitC10mg compared with the SARS-CoV-2-Alum + VitC1mg and SARS-CoV-2-Alum + VitC10mg groups (p > 0.2810) (Fig. 1B).
IL-2 cytokine in aged mice
Results from the IL-2 cytokine assay in the aged groups revealed that mice immunized with all vaccine formulations showed a significant increase in IL-2 levels compared with the PBS control group (p < 0.0169). In addition, mice immunized with inactivated SARS-CoV-2-Alum + α-tocopherol + VitC10mg led to a 13.92% increase in IL-2 levels compared with the SARS-CoV-2-Alum group (p = 0.8612). No significant differences in IL-2 levels were observed among the vaccinated groups (p > 0.2465) (Fig. 2A).
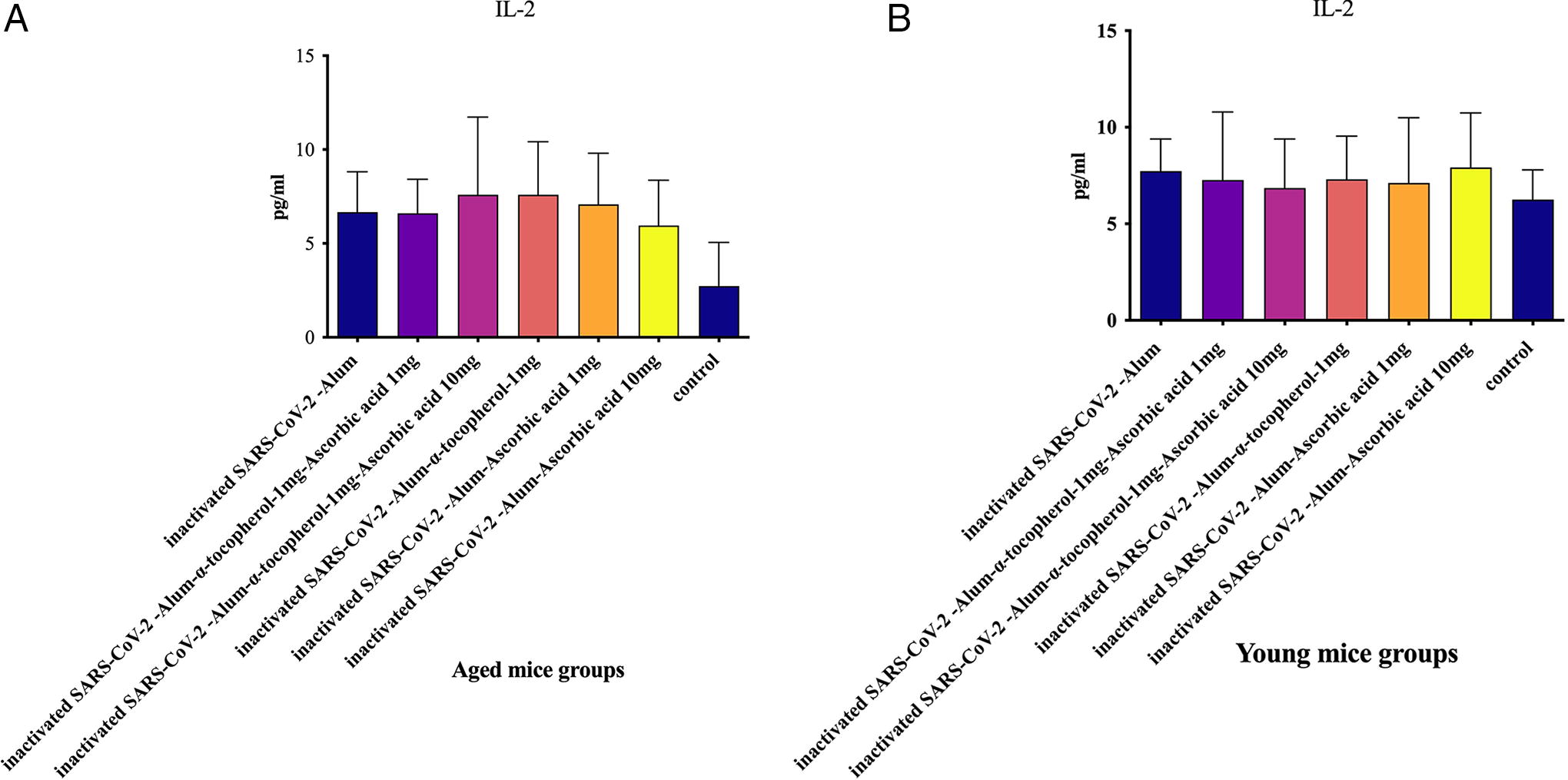
IL-2 in young mice
The results from the IL-2 cytokine assay in the young groups revealed no significant differences in IL-2 levels among the vaccinated groups (p > 0.3206). In addition, in the vaccinated groups there were increased, but not significant, IL-2 cytokine responses compared with the PBS control group (p > 0.2677) (Fig. 2B).
IL-4 cytokine in aged mice
Results from the IL-4 cytokine assay indicated that mice immunized with inactivated SARS-CoV-2-Alum resulted in a significant decrease in the IL-4 level compared with the PBS control group (p = 0.0426); however, SARS-CoV-2-Alum + α-tocopherol showed a borderline decrease in IL-4 levels compared with the PBS control group (p = 0.0507). In addition, mice immunized with inactivated SARS-CoV-2-Alum + α-tocopherol + VitC1mg and inactivated SARS-CoV-2-Alum + α-tocopherol + VitC10mg exhibited a 42.15% and 94.43% increase in IL-4 levels, respectively, compared with the SARS-CoV-2-Alum group (p = 0.3975 and p = 0.5054, respectively). Furthermore, mice immunized with SARS-CoV-2-Alum + VitC1mg showed a 94.54% increase in the IL-4 level compared with the SARS-CoV-2-Alum group (p = 0.5737), whereas those immunized with SARS-CoV-2-Alum + α-tocopherol + VitC10mg showed a significant increase in IL-4 levels compared with the SARS-CoV-2-Alum group (p = 0.0401) (Fig. 3A).
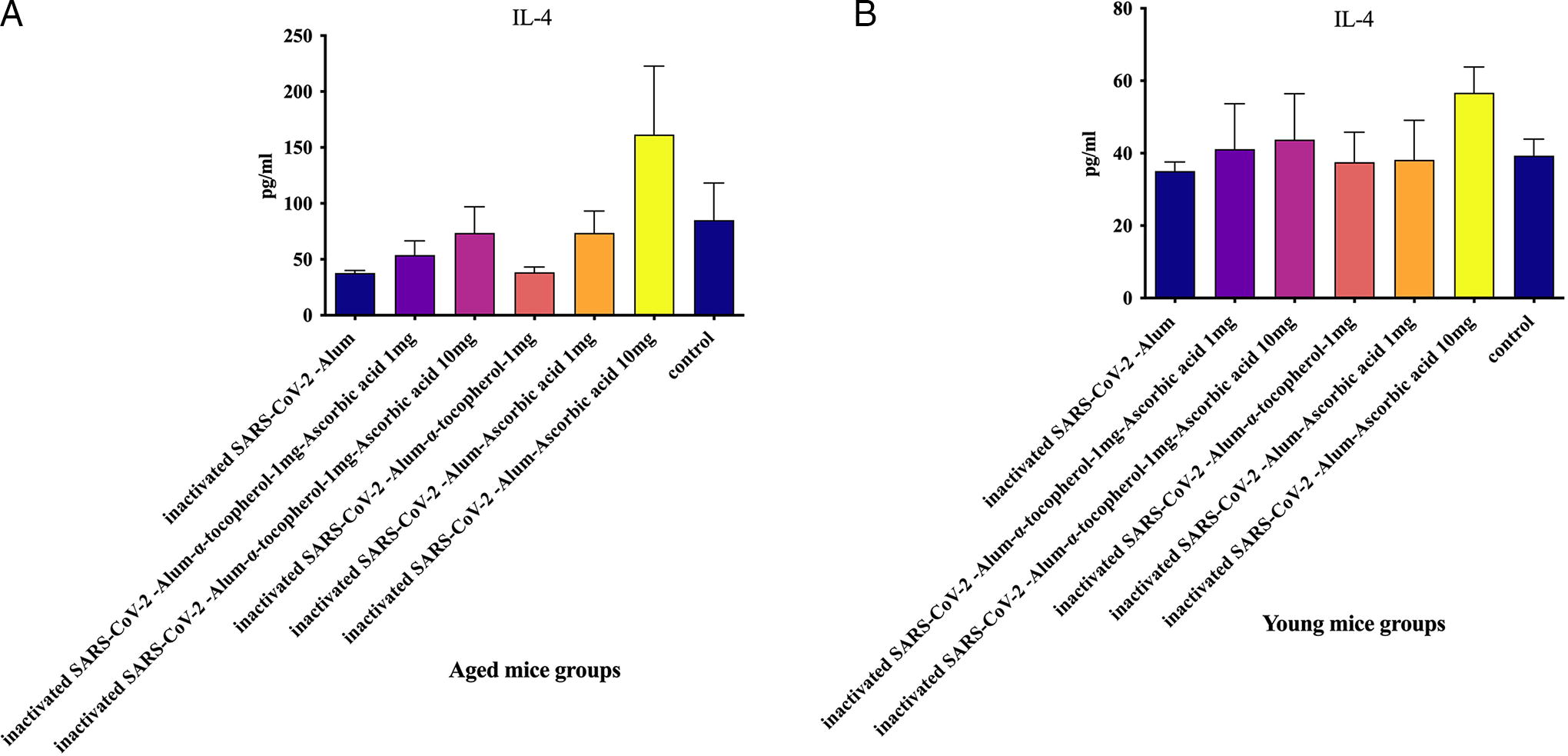
IL-4 in young mice
The results of the IL-4 cytokine assay in the experimental groups showed that mice immunized with SARS-CoV-2-Alum + VitC10mg displayed a significant increase in IL-4 levels compared with the SARS-CoV-2-Alum and SARS-CoV-2-Alum + α-tocopherol + VitC1mg groups (p = 0.0350 and p = 0.0183, respectively). In addition, mice immunized with SARS-CoV-2-Alum + VitC10mg showed a significant increase in IL-4 levels compared with the SARS-CoV-2-Alum + α-tocopherol and SARS-CoV-2-Alum + VitC1mg groups (p = 0.0350 and p = 0.0350, respectively). Furthermore, mice immunized with inactivated SARS-CoV-2-Alum + α-tocopherol + VitC1mg and inactivated SARS-CoV-2-Alum + α-tocopherol + VitC10mg showed a 17.25% and 24.76% increase in IL-4 levels, respectively, compared with the SARS-CoV-2-Alum group (p = 0.2673 and p = 0.5899, respectively) (Fig. 3B).
IFN-γ/IL-4 ratio in aged mice
Results from the IFN-γ/IL-4 ratio in the aged experimental groups demonstrated that mice immunized with inactivated SARS-CoV-2-Alum + α-tocopherol + VitC1mg and inactivated SARS-CoV-2-Alum + α-tocopherol + VitC10mg showed a significant increase in the IFN-γ/IL-4 ratio compared with those immunized with PBS (p = 0.0109 and p = 0.0295, respectively). However, mice immunized with SARS-CoV-2-Alum + α-tocopherol + VitC1mg and SARS-CoV-2-Alum + α-tocopherol + VitC10mg showed a 46.94% and 37.95% increase in the IFN-γ/IL-4 ratio compared with the SARS-CoV-2-Alum group (p = 0.2319 and p = 0.4634, respectively). No significant differences were observed in the IFN-γ/IL-4 ratio between experimental groups (Fig. 4A).
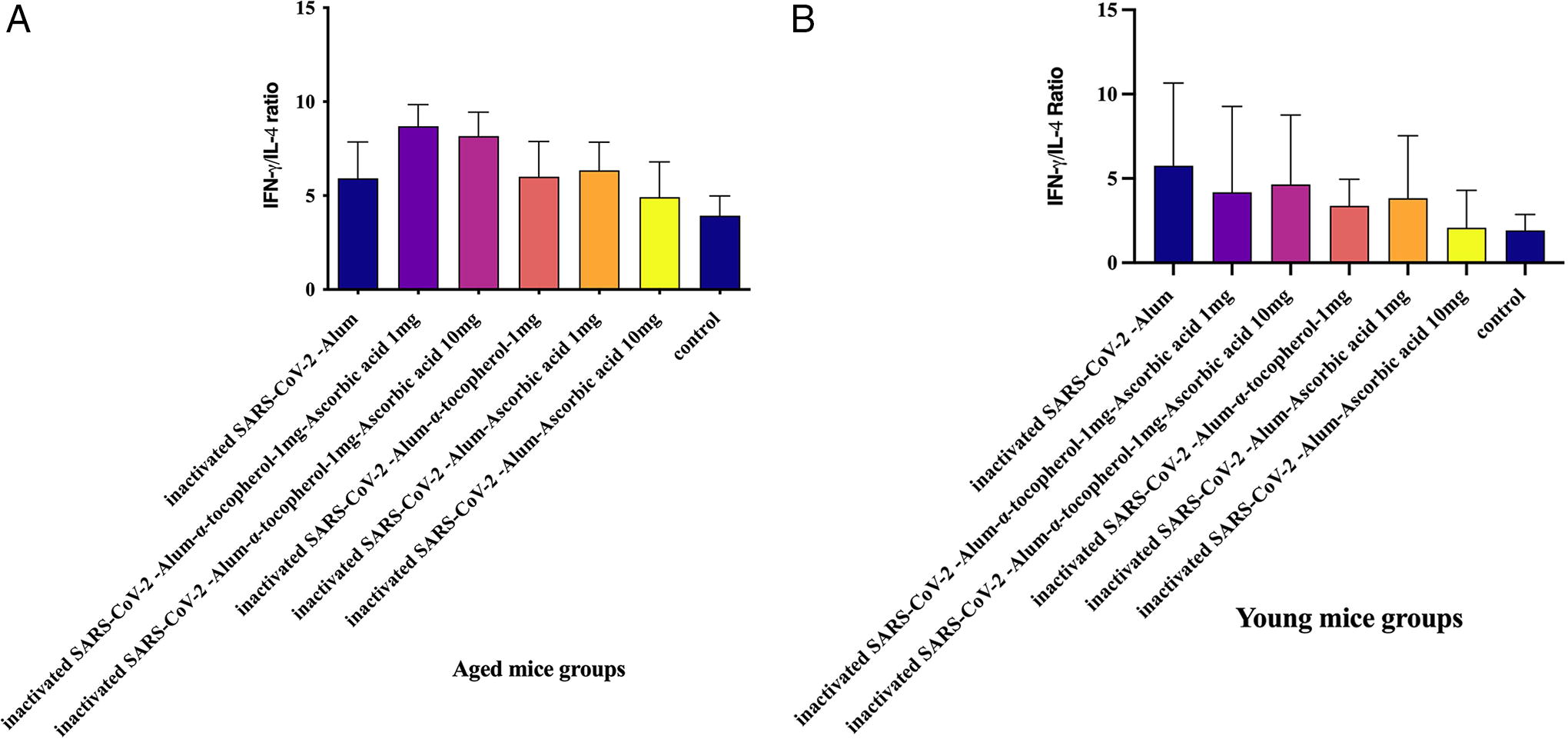
IFN-γ/IL-4 ratio in young mice
The IFN-γ/IL-4 ratio in the young experimental groups demonstrated no significant differences among the experimental groups (p > 0.0734) (Fig. 4B).
Specific total IgG response in aged mice
Results from the specific total IgG response in aged experimental groups revealed that mice immunized with inactivated SARS-CoV-2-Alum (at dilutions of 1/25 up to 1/1600; p < 0.0006), inactivated SARS-CoV-2-Alum + α-tocopherol + VitC1mg (at dilutions of 1/25 up to 1/400; p < 0.0078), inactivated SARS-CoV-2-Alum + α-tocopherol + VitC10mg (at dilutions of 1/25 up to 1/200; p < 0.0179), inactivated SARS-CoV-2-Alum + α-tocopherol (at dilutions of 1/25 up to 1/1600; p < 0.0111), inactivated SARS-CoV-2-Alum + VitC1mg (at dilutions of 1/25 up to 1/800; p < 0.0376), and inactivated SARS-CoV-2-Alum + VitC10mg (at dilutions of 1/25 up to 1/200; p < 0.0219) showed significant differences in the IgG response compared with the PBS group. In addition, mice immunized with inactivated SARS-CoV-2-Alum + α-tocopherol + VitC1mg (at dilutions of 1/25 up to 1/1600; p < 0.0483), inactivated SARS-CoV-2-Alum + α-tocopherol + VitC10mg (at dilutions of 1/25 up to 1/1600; p < 0.0367), inactivated SARS-CoV-2-Alum + VitC1mg (at dilutions of 1/25 up to 1/800; p < 0.0184), and inactivated SARS-CoV-2-Alum + VitC10mg (at dilutions of 1/25 up to 1/1600; p < 0.0107) showed a significant decrease in the IgG response compared with the SARS-CoV-2-Alum group. Mice immunized with inactivated SARS-CoV-2-Alum + VitC10mg at dilutions of 1/25 up to 1/200 showed a significant decrease in the IgG response compared with the inactivated SARS-CoV-2-Alum + VitC1mg group (p < 0.0424). Furthermore, mice immunized with inactivated SARS-CoV-2-Alum + α-tocopherol + VitC10mg at dilutions of 1/25 and 1/50 showed a significant decrease in the IgG response compared with the inactivated SARS-CoV-2-Alum + α-tocopherol + VitC1mg group (p < 0.0018). In addition, mice immunized with inactivated SARS-CoV-2-Alum + α-tocopherol showed a significant increase in the IgG response compared with those immunized with inactivated SARS-CoV-2-Alum + VitC1mg (at dilutions of 1/25 up to 1/400; p < 0.0015), SARS-CoV-2-Alum + VitC10mg (at dilutions of 1/25 up to 1/800; p < 0.0002), SARS-CoV-2-Alum + α-tocopherol + VitC1mg (at dilutions of 1/25 up to 1/800; p < 0.0030), and SARS-CoV-2-Alum + α-tocopherol + VitC10mg (at dilutions of 1/25 up to 1/800; p < 0.0005) (Fig. 5A).
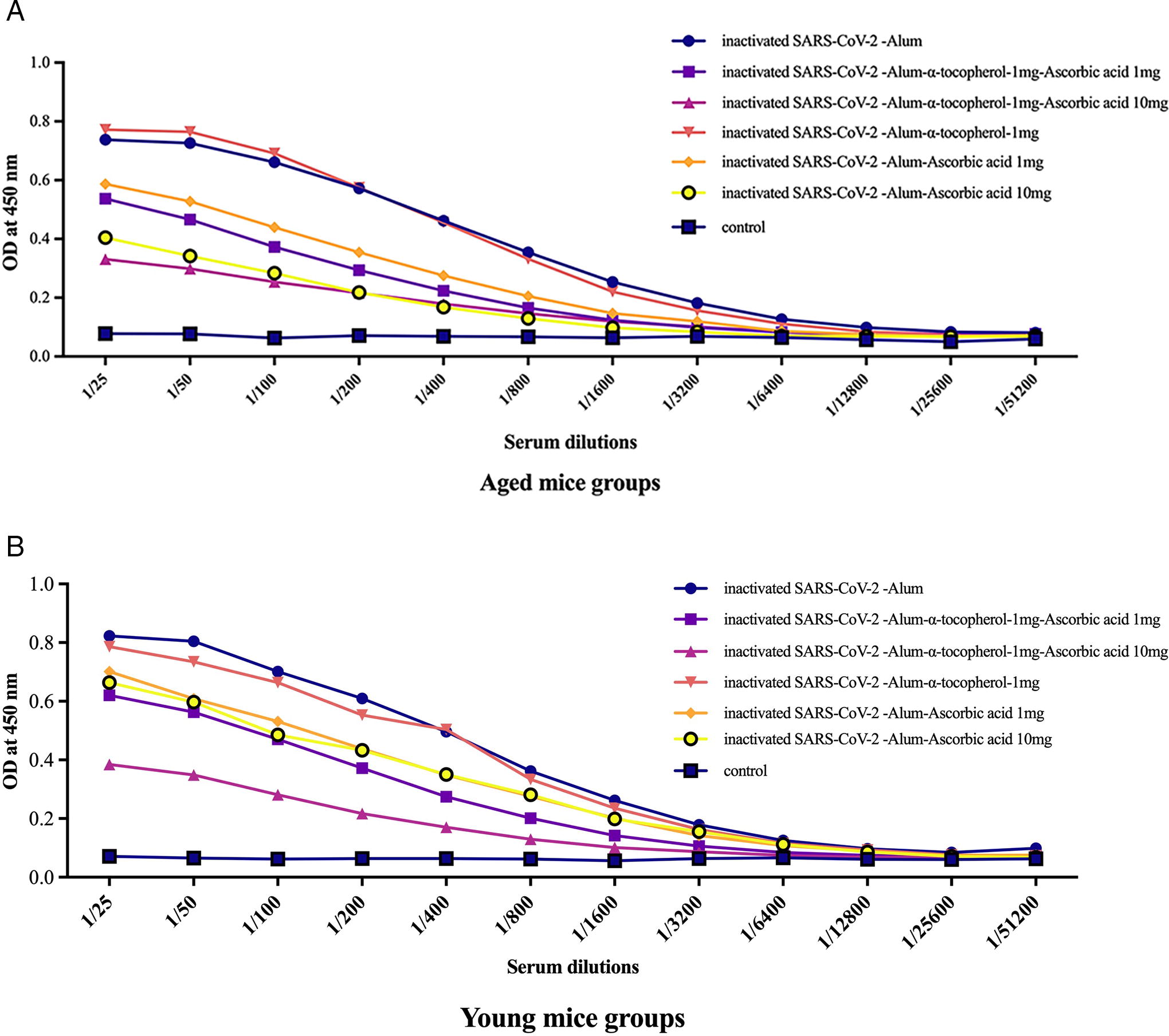
Specific total IgG response in young mice
Results from the specific total IgG in young experimental groups revealed that mice immunized with inactivated SARS-CoV-2-Alum (at dilutions of 1/25 to 1/1600; p < 0.0002), inactivated SARS-CoV-2-Alum + α-tocopherol + VitC1mg (at dilutions of 1/25 to 1/800; p < 0.0298), inactivated SARS-CoV-2-Alum + α-tocopherol + VitC10mg (at dilutions of 1/25 to 1/200; p < 0.0112), inactivated SARS-CoV-2-Alum + α-tocopherol (at dilutions of 1/25 to 1/1600; p < 0.0021), inactivated SARS-CoV-2-Alum + VitC1mg (at dilutions of 1/25 to 1/1600; p < 0.0273), and inactivated SARS-CoV-2-Alum + VitC10mg (at dilutions of 1/25 to 1/1600; p < 0.04787) showed significant differences in IgG compared with the PBS group. In addition, mice immunized with inactivated SARS-CoV-2-Alum + α-tocopherol + VitC1mg (at dilutions of 1/25 up to 1/800; p < 0.0061), inactivated SARS-CoV-2-Alum + α-tocopherol + VitC10mg (at dilutions of 1/25 up to 1/1600; p < 0.0063), inactivated SARS-CoV-2-Alum + VitC1mg (at dilutions of 1/25 up to 1/400; p < 0.0240), and inactivated SARS-CoV-2-Alum + VitC10mg (at dilutions of 1/25 up to 1/400; p < 0.0390) showed a significant decrease in the IgG response compared with the SARS-CoV-2-Alum group. Mice immunized with inactivated SARS-CoV-2-Alum + α-tocopherol + VitC1mg at dilutions of 1/25 and 1/200 showed a significant decrease in the IgG response compared with the inactivated SARS-CoV-2-Alum + α-tocopherol + VitC10mg group (p < 0.0063). In addition, mice immunized with inactivated SARS-CoV-2-Alum + α-tocopherol group showed a significant increase in the IgG response compared with the SARS-CoV-2-Alum + VitC10mg (at dilutions of 1/25 up to 1/100; p < 0.0041), SARS-CoV-2-Alum + α-tocopherol + VitC1mg (at dilutions of 1/25 up to 1/800; p < 0.0479), and SARS-CoV-2-Alum + α-tocopherol + VitC10mg groups (at dilutions of 1/25 up to 1/1600; p < 0.0448) (Fig. 5B).
Specific IgG1 isotype in aged mice
Results from the specific IgG1 isotype response in aged experimental groups revealed that mice immunized with inactivated SARS-CoV-2-Alum, inactivated SARS-CoV-2-Alum + α-tocopherol + VitC1mg, inactivated SARS-CoV-2-Alum + α-tocopherol + VitC10mg, inactivated SARS-CoV-2-Alum + α-tocopherol, inactivated SARS-CoV-2-Alum + VitC1mg, and inactivated SARS-CoV-2-Alum + VitC10mg showed a significant difference in the IgG1 response compared with the PBS group (p < 0.0012). In addition, mice immunized with inactivated SARS-CoV-2-Alum + α-tocopherol, inactivated SARS-CoV-2-Alum + α-tocopherol + VitC10mg, inactivated SARS-CoV-2-Alum + VitC1mg, and inactivated SARS-CoV-2-Alum + VitC10mg showed a significant decrease in the IgG1 response compared with the SARS-CoV-2-Alum group (p < 0.0006). Mice immunized with inactivated SARS-CoV-2-Alum + VitC10mg showed a borderline decrease in the IgG1 response compared with the inactivated SARS-CoV-2-Alum + VitC1mg group (p < 0.0503). Furthermore, mice immunized with inactivated SARS-CoV-2-Alum + α-tocopherol + VitC10mg showed a significant decrease in the IgG1 response compared with the inactivated SARS-CoV-2-Alum + α-tocopherol + VitC1mg group (p < 0.0012). In addition, mice immunized with inactivated SARS-CoV-2-Alum + α-tocopherol showed a significant decrease in the specific IgG1 response compared with those immunized with inactivated SARS-CoV-2-Alum + VitC1mg (p < 0.0291) and inactivated SARS-CoV-2-Alum + α-tocopherol + VitC1mg (p < 0.0006) (Fig. 6A).

Specific IgG1 isotype in young mice
Results from the specific IgG1 isotype response in young experimental groups revealed that mice immunized with inactivated SARS-CoV-2-Alum, inactivated SARS-CoV-2-Alum + α-tocopherol + VitC1mg, inactivated SARS-CoV-2-Alum + α-tocopherol + VitC10mg, inactivated SARS-CoV-2-Alum + α-tocopherol, inactivated SARS-CoV-2-Alum + VitC1mg, and inactivated SARS-CoV-2-Alum + VitC10mg exhibited significant differences in the IgG1 response compared with the PBS group (p < 0.0012). In addition, mice immunized with inactivated SARS-CoV-2-Alum + α-tocopherol + VitC1mg, inactivated SARS-CoV-2-Alum + α-tocopherol + VitC10mg, and inactivated SARS-CoV-2-Alum + VitC10mg (p < 0.0012) showed a significant decrease in the IgG1 response compared with the SARS-CoV-2-Alum group. Mice immunized with inactivated SARS-CoV-2-Alum + VitC10mg showed a significant decrease in the IgG1 response compared with the inactivated SARS-CoV-2-Alum + VitC1mg group (p = 0.0052). Furthermore, mice immunized with inactivated SARS-CoV-2-Alum + α-tocopherol + VitC10mg at dilutions showed a significant decrease in the IgG1 response compared with the inactivated SARS-CoV-2-Alum + α-tocopherol + VitC1mg group (p = 0.0344). In addition, mice immunized with inactivated SARS-CoV-2-Alum + α-tocopherol showed a significant increase in the specific IgG1 response compared with the inactivated SARS-CoV-2-Alum + VitC1mg (p = 0.0402), inactivated SARS-CoV-2-Alum + VitC10mg (p = 0.0012), inactivated SARS-CoV-2-Alum + α-tocopherol + VitC1mg (p = 0.0023), and inactivated SARS-CoV-2-Alum + α-tocopherol + VitC10mg groups (p = 0.0006) (Fig. 6B).
Specific IgG2a isotype in aged mice
Results from the specific IgG2a isotype response in aged experimental groups revealed that mice immunized with inactivated SARS-CoV-2-Alum + α-tocopherol + VitC1mg (p = 0.0146) and inactivated SARS-CoV-2-Alum + VitC10mg (p = 0.0128) showed a significant decrease in the IgG2a response compared with the SARS-CoV-2-Alum group. In addition, mice immunized with inactivated SARS-CoV-2-Alum + α-tocopherol showed a significant increase in the specific IgG2a response compared with those immunized with inactivated SARS-CoV-2-Alum + VitC10mg (p = 0.0134) and inactivated SARS-CoV-2-Alum + α-tocopherol + VitC1mg (p < 0.0443) (Fig. 7A).

Specific IgG2a isotype in young mice
Results from the specific IgG2a isotype antibody in young experimental groups revealed a small increase in the IgG2a antibody in the inactivated SARS-CoV-2-Alum + α-tocopherol group compared with all other experimental groups, although the difference was not statistically significant (p > 0.4441) (Fig. 7B).
IgG2a/IgG1 ratio in aged mice
Results from the IgG2a/IgG1 ratio in aged experimental groups revealed that mice immunized with inactivated SARS-CoV-2-Alum + α-tocopherol + VitC1mg (p = 0.0012), inactivated SARS-CoV-2-Alum + α-tocopherol + VitC10mg (p = 0.0175), and inactivated SARS-CoV-2-Alum + VitC10mg (p = 0.0006) showed a significant increase in the IgG2a/IgG1 ratio compared with the inactivated SARS-CoV-2-Alum group. Mice immunized with inactivated SARS-CoV-2-Alum + VitC10mg showed a significant increase in the IgG2a/IgG1 ratio compared with the inactivated SARS-CoV-2-Alum + α-tocopherol group (p = 0.0006). Furthermore, mice immunized with inactivated SARS-CoV-2-Alum + α-tocopherol + VitC10mg showed a significant increase in the IgG2a/IgG1 ratio compared with the inactivated SARS-CoV-2-Alum + α-tocopherol group (p = 0.0262). However, mice immunized with inactivated SARS-CoV-2-Alum + α-tocopherol + VitC10mg showed a 21.20% and 29.94% increase in the IgG2a/IgG1 ratio, respectively, compared with those immunized with inactivated SARS-CoV-2-Alum + α-tocopherol + VitC1mg (p = 0.1649) and inactivated SARS-CoV-2-Alum + VitC1mg (p = 0.0728), respectively (Fig. 8A).
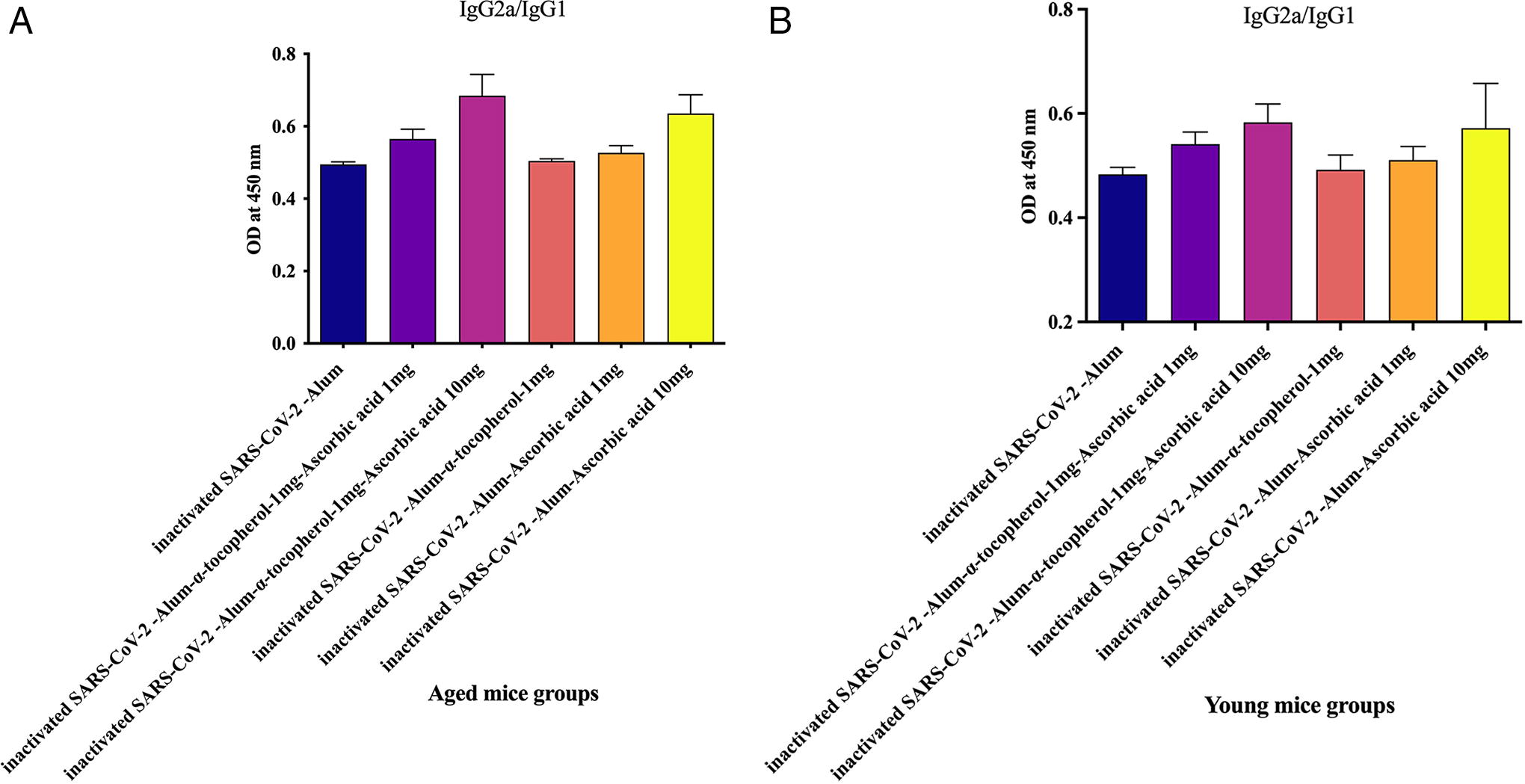
IgG2a/IgG1 ratio in young mice
Results from the IgG2a/IgG1 ratio in young experimental groups revealed that mice immunized with inactivated SARS-CoV-2-Alum + α-tocopherol + VitC1mg (p = 0.0006), inactivated SARS-CoV-2-Alum + α-tocopherol + VitC10mg (p = 0.0006), and inactivated SARS-CoV-2-Alum + VitC10mg (p = 0.0012) showed a significant increase in the IgG2a/IgG1 ratio compared with the inactivated SARS-CoV-2-Alum group. Mice immunized with inactivated SARS-CoV-2-Alum + VitC10mg showed a significant increase in the IgG2a/IgG1 ratio compared with the inactivated SARS-CoV-2-Alum + α-tocopherol group (p = 0.0350), but exhibited a 12.02% increase compared with the inactivated SARS-CoV-2-Alum + VitC1mg group (p = 0.1948). Furthermore, mice immunized with inactivated SARS-CoV-2-Alum + α-tocopherol + VitC10mg showed a significant increase in the IgG2a/IgG1 ratio compared with those immunized with inactivated SARS-CoV-2-Alum + α-tocopherol (p = 0.0262), inactivated SARS-CoV-2-Alum + α-tocopherol (p = 0.0006), and inactivated SARS-CoV-2-Alum + α-tocopherol + VitC1mg (p = 0.0006) (Fig. 8B).
PD1 gene expression in aged mice
Results from PD1 gene expression in aged experimental groups demonstrated that immunization with inactivated SARS-CoV-2-Alum + α-tocopherol + VitC1mg, inactivated SARS-CoV-2-Alum + α-tocopherol + VitC10mg, and inactivated SARS-CoV-2-Alum + VitC10mg led to a significant decrease in PD1 gene expression compared with the PBS control group (p < 0.0453). In addition, mice immunized with inactivated SARS-CoV-2-Alum + α-tocopherol + VitC1mg and inactivated SARS-CoV-2-Alum + α-tocopherol + VitC10mg showed a significant decrease in PD1 gene expression compared with the SARS-CoV-2-Alum group (p = 0.0286 and p = 0.0001, respectively). Furthermore, mice immunized with inactivated SARS-CoV-2-Alum + α-tocopherol + VitC10mg showed a significant decrease in PD1 gene expression compared with the inactivated SARS-CoV-2-Alum + α-tocopherol + VitC1mg, inactivated SARS-CoV-2-Alum + α-tocopherol, inactivated SARS-CoV-2-Alum + VitC1mg, and inactivated SARS-CoV-2-Alum + VitC10mg groups (p < 0.0002) (Fig. 9A).
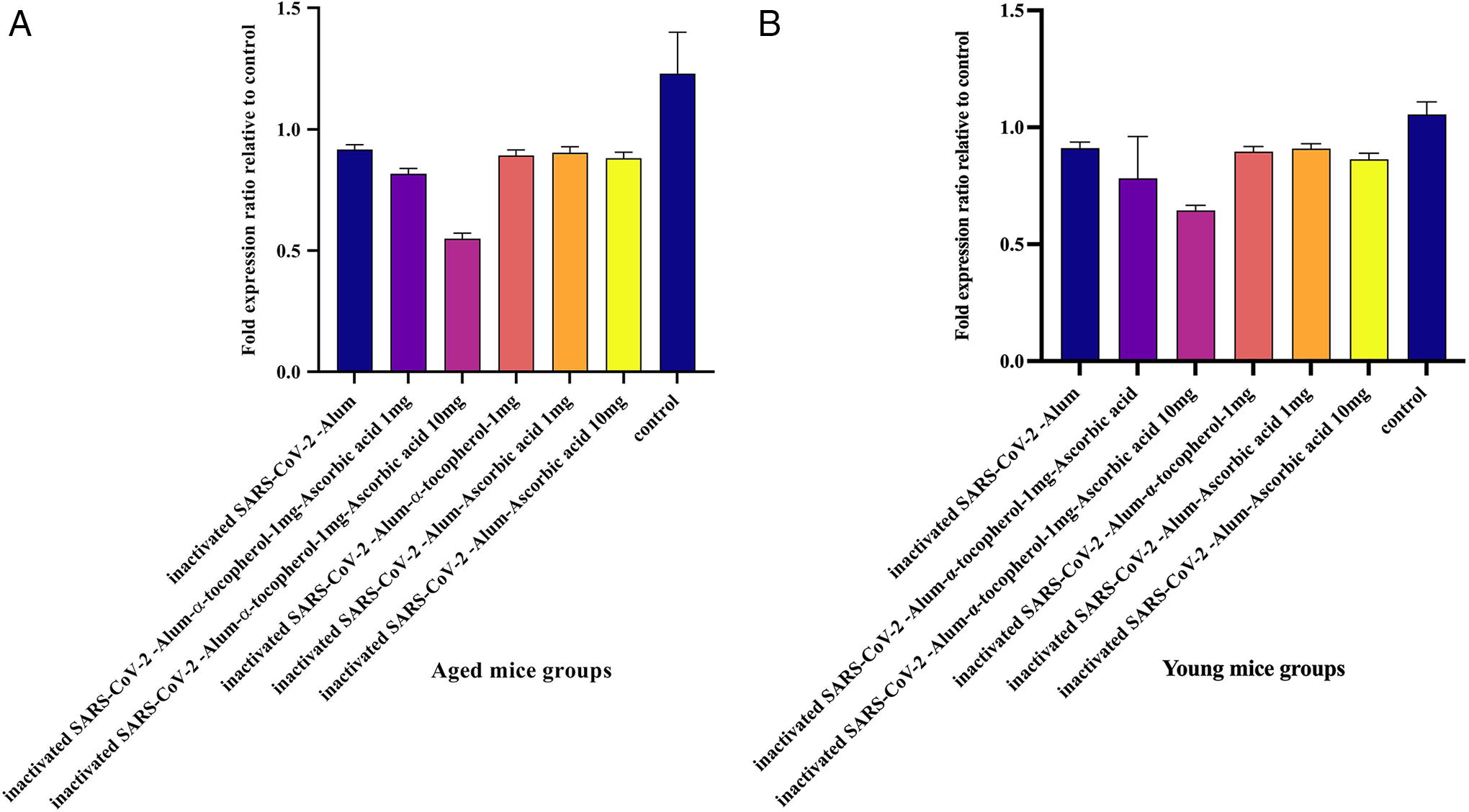
PD1 gene expression in young mice
Results from PD1 gene expression in young experimental groups demonstrated that immunization with inactivated SARS-CoV-2-Alum + α-tocopherol + VitC10mg, inactivated SARS-CoV-2-Alum + α-tocopherol, inactivated SARS-CoV-2-Alum + VitC1mg, and inactivated SARS-CoV-2-Alum + VitC10mg led to a significant decrease in PD1 gene expression compared with the PBS control group (p = 0.0001, p = 0.0388, p = 0.0046, and p = 0.0261, respectively). In addition, mice immunized with inactivated SARS-CoV-2-Alum + α-tocopherol + VitC10mg showed a significant decrease in PD1 gene expression compared with the SARS-CoV-2-Alum group (p = 0.0010). Furthermore, mice immunized with inactivated SARS-CoV-2-Alum + α-tocopherol + VitC10mg displayed a significant decrease in PD1 gene expression compared with the inactivated SARS-CoV-2-Alum + α-tocopherol, inactivated SARS-CoV-2-Alum + VitC1mg, and inactivated SARS-CoV-2-Alum + VitC10mg groups (p = 0.0009, p = 0.0001, and p = 0.0025, respectively) (Fig. 9B).
TERT gene expression in aged mice
Results from TERT gene expression in aged experimental groups demonstrated that immunization with inactivated SARS-CoV-2-Alum, inactivated SARS-CoV-2-Alum + α-tocopherol + VitC1mg, inactivated SARS-CoV-2-Alum + α-tocopherol + VitC10mg, inactivated SARS-CoV-2-Alum + α-tocopherol, inactivated SARS-CoV-2-Alum + VitC1mg, and inactivated SARS-CoV-2-Alum + VitC10mg led to a significant increase in TERT gene expression compared with the PBS control group (p < 0.0413). In addition, mice immunized with inactivated SARS-CoV-2-Alum + α-tocopherol + VitC1mg and inactivated SARS-CoV-2-Alum + α-tocopherol + VitC10mg showed a significant increase in TERT gene expression compared with the SARS-CoV-2-Alum group (p = 0.0042 and p = 0.0001, respectively). Mice immunized with inactivated SARS-CoV-2-Alum + α-tocopherol + VitC10mg showed a significant increase in TERT gene expression compared with inactivated SARS-CoV-2-Alum + α-tocopherol + VitC1mg, inactivated SARS-CoV-2-Alum + α-tocopherol, inactivated SARS-CoV-2-Alum + VitC1mg, and inactivated SARS-CoV-2-Alum + VitC10mg groups (p = 0.0013, p = 0.0002, p = 0.0059, and p = 0.0163, respectively) (Fig. 10A).
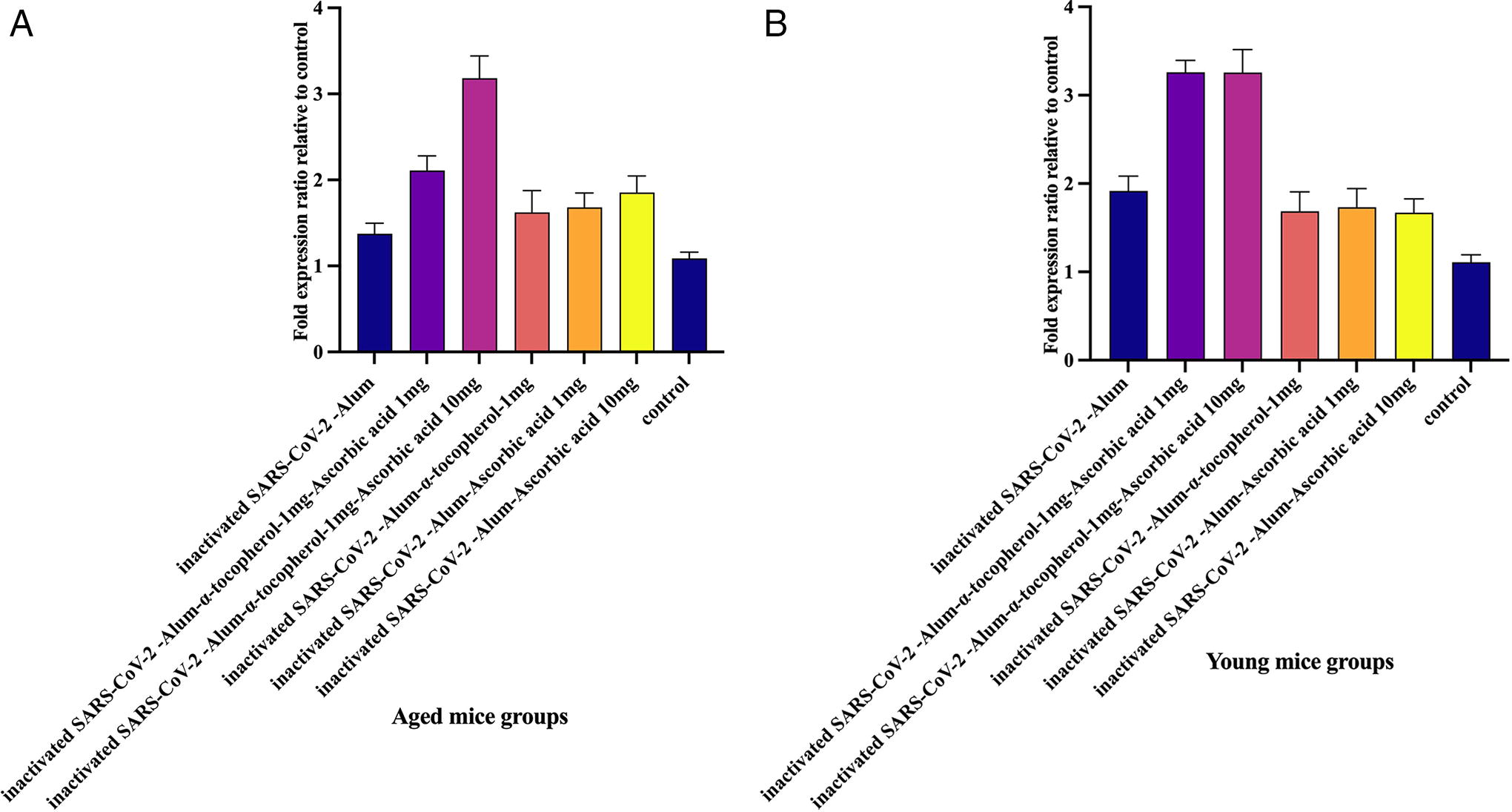
TERT gene expression in young mice
Results from TERT gene expression in young experimental groups demonstrated that immunization with inactivated SARS-CoV-2-Alum, inactivated SARS-CoV-2-Alum + α-tocopherol + VitC1mg, inactivated SARS-CoV-2-Alum + α-tocopherol + VitC10mg, inactivated SARS-CoV-2-Alum + VitC1mg, and inactivated SARS-CoV-2-Alum + VitC10mg led to a significant increase in TERT gene expression compared with the PBS control group (p < 0.0155), whereas immunization with inactivated SARS-CoV-2-Alum + α-tocopherol showed a borderline increase in TERT gene expression compared with the PBS control group (p = 0.0602). In addition, mice immunized with inactivated SARS-CoV-2-Alum + α-tocopherol + VitC1mg and inactivated SARS-CoV-2-Alum + α-tocopherol + VitC10mg showed a significant increase in TERT gene expression compared with those immunized with SARS-CoV-2-Alum (p = 0.0004 and p = 0.0078, respectively) (Fig. 10B).
Discussion
According to official reports on COVID-19, approximately 22% of mortality is attributed to the elderly. In fact, the mortality rate from COVID‐19 is associated with age, and elderly patients with COVID‐19 are more vulnerable. The cause of high mortality in aged people can be attributed to the decline in the function of the immune system (Blagosklonny, 2020; Ciabattini et al., 2020; Dadras et al., 2022; Kang and Jung, 2020; Li et al., 2021; Park et al., 2021). However, various methods such as increasing the vaccine dose, changing the injection route, and developing new adjuvants have been introduced in vaccinology to increase vaccine efficacy in the elderly (Li et al., 2021). In the present study, we hypothesized that the use of α-tocopherol as an adjuvant along with VitC formulated in the inactivated SARS-CoV-2 vaccine might increase vaccine potency in aged mice. Evidence has shown a dramatic decline in T cell function and cytokine release, especially the IFN-γ response, in aged people, which in turn explains why the mortality rate increases in aged people, compared with young people, following infections with viruses and intracellular pathogens (McElhaney et al., 2016; Ouyang et al., 2000). Therefore, restoring T cell function and the IFN-γ cytokine response in aged people may improve immune responses to vaccines. Our results demonstrated that the vaccines formulated in VitC alone or VitC plus α-tocopherol showed an increase in IFN-γ levels compared with the Alum-based vaccine in aged and young mice, and this effect was more evident in aged mice than in young mice. In addition, no synergistic effect was found between VitC and α-tocopherol on the IFN-γ cytokine response. While the VitC-formulated vaccine showed a significant effect only in the aged group, α-tocopherol exhibited no positive effect on the IFN-γ cytokine response in the aged and young groups. Accumulating evidence has shown that VitC and α-tocopherol have immunomodulatory effects. Our previous study on the adjuvant effect of α-tocopherol on the influenza vaccine showed a significant increase in the IFN-γ cytokine response in the aged mice, but not in the young mice (Eshraghi et al., 2022). In addition, a study on α-tocopherol in an influenza DNA vaccine model showed a significant increase in IFN-γ cytokine response (Karlsson et al., 2017a). Nevertheless, we did not obtain this finding in this study, which may be related to the nature of the antigen used in this study.
Studies have shown that the level of IFN-γ is lower in patients with COVID-19 hospitalized in the ICU than in those with milder symptoms (Kwon et al., 2020). In addition, the level of IFN-γ has been shown to decrease with age, which multiplies the sensitivity of the elderly to infectious diseases, such as COVID-19 (Lin et al., 2022; Palacios-Gutiérrez et al., 2022). Studies have shown that vaccine efficiency seems to be lower in the elderly than in others because of the low level of IFN-γ; notably, improved release of the IFN-γ cytokine in response to vaccines may circumvent this problem (Pawelec, 2018; Zhang et al., 2021; Zhou et al., 2020a). A study conducted in mice showed that the use of VitC at the initial time of infection could inhibit and control the viral responses to influenza A in the lungs of Gulo (-/-) mice through increased production of IFN-γ (Kim et al., 2013). In addition, the use of VitC intravenously or as a supplement in COVID-19 and acute respiratory syndrome leads to a reduction in inflammation and accelerates the healing process in elderly patients (Bae and Kim, 2020). Our findings showed no significant effect of the IL-2 cytokine response in the aged and young groups. In contrast, an increase in the IL-4 cytokine response was found in aged mice immunized with VitC alone or VitC plus α-tocopherol compared with the Alum-based vaccine. However, VitC10mg showed a significant increase in IL-4 cytokine levels in young mice compared to the Alum-based vaccine. In a study conducted by Noh et al., mice were intraperitoneally injected with 5 mg VitC to respond to dinitro-1-infuorobenzene. Their results showed an increased level of cytokine IFN-γ (Th1) after activation, while a lower proportion of cytokine IL-4 (Th2) was discovered after injection in mice (Noh et al., 2005). It seems that VitC formulated in the vaccine improved both IFN-γ and IL-4 cytokine responses, which are critical for controlling viral infections (Mahdavi et al., 2011). As another Th1 parameter judgment, the IFN-γ/IL-4 ratio in the SARS-CoV-2-Alum + α-tocopherol + VitC1mg and SARS-CoV-2-Alum + α-tocopherol + VitC10mg groups of aged mice showed a 46.94% and 37.95% increase, respectively, compared with that in the SARS-CoV-2-Alum group. This finding is further confirmation of Th1 polarization (Haghighi et al., 2022).
Results from the specific IgG response in the aged and young groups showed a suppression in humoral immune response when VitC was used alone or in combination with α-tocopherol in the vaccine formulation; however, the vaccine formulated with α-tocopherol showed a slight increase in the humoral immune response in the aged group, which was not significant. Our previous study on the H1N1 influenza vaccine model showed a positive effect of α-tocopherol in the improvement of humoral immune response, especially in the aged mice group (Eshraghi et al., 2022). In addition, α-tocopherol in HBsAg vaccine formulations improved specific IgG responses (Rahimkhani et al., 2021). Nonetheless, α-tocopherol exhibited no positive effects in the present study, which may be due to the nature of the antigen used. Furthermore, VitC and VitC plus α-tocopherol in the vaccine formulation suppressed the IgG response, which seems to have no synergistic effect, and the suppression appears to be due to VitC in the vaccine formulation. Parallel to specific total IgG, a decrease was found in the IgG1 and IgG2a isotypes in these groups compared with the SARS-CoV-2-Alum vaccine. All of these findings regarding the humoral immune response show suppression, which may be due to VitC in the vaccine formulation. Several studies have shown that VitC has a positive effect on humoral immune responses (Qi et al., 2020; Wu et al., 2000). While used as an oral supplement in other studies, VitC was used as an adjuvant in our study; as a result, the results from these studies were not in parallel with our findings. Some parameters, such as VitC dose, antigen nature, administration dose, and route may be involved in achieving these results. An oil adjuvant model showed that antioxidants could inhibit the enhancement of corresponding immune responses against an immunogen, indicating the critical role of oxygen radicals in the immune response, and inhibition of oxygen radicals during an immune response may not have positive effects on the immune response (Di Gianni et al., 1996). However, the dose of VitC seems to play an important role in the immunomodulatory effect; as a result, antioxidant agents such as VitC may decrease the adjuvant effect of a vaccine formulation by capturing free radicals. Furthermore, mice immunized with vaccines formulated in VitC and VitC plus α-tocopherol showed a significant increase in the IgG2a/IgG1 ratio in the aged and young groups, which is another confirmation of Th1 polarization.
In addition, we checked the expression levels of the genes involved in aging, which play a critical role in vaccine efficacy. Our results showed that the inactivated SARS-CoV-2-Alum vaccine formulated in α-tocopherol + VitC decreased PD1 gene expression in aged and young mice. In addition, the expression of the TERT gene, which shows telomerase activity, increased in both old and young mice immunized with inactivated SARS-CoV-2-Alum formulated with α-tocopherol and VitC. Studies have shown that aging is accompanied by high expression of immune checkpoint receptors. PD1 is highly expressed on the immune cells of aged people, which may play a critical role in the decline of the immune response in aging (Garcia et al., 2022; Onorati et al., 2022; Wang et al., 2022). Therefore, decreased expression of PD1 in immune cells may reinforce the immune responses of aged people. In addition, expression of the TERT gene decreases with age; studies have shown that this condition is correlated with immune system function, which shows a decline in the immune response (Weng et al., 1997; Zhdanov et al., 2018a; Zhdanov et al., 2018b; Zhdanov et al., 2018c). The results of the present study showed an increase and decrease in the TERT and PD1 levels, respectively. These results suggest that the vaccine formulations (α-tocopherol and VitC) developed in this study may affect the components of the immune system by changing the cell physiology, which was more in the elderly population. Different researches showed that simultaneous use of vitamin E and VitC supplements improves the function of the immune system in the elderly, controls infections, and reduces the treatment duration (De la Fuente et al., 2008; De la Fuente et al., 2020; Kim et al., 2013; Lee and Han, 2018; Meydani et al., 2018). In a study carried out by Fuente et al., the use of vitamin C and vitamin E supplements simultaneously by elderly people was demonstrated not only to have positive effects on patients but also to decrease the severity of the infection and the duration of the treatment. Furthermore, a study on these people indicated that the simultaneous use of these two supplements can be useful for rejuvenating the immune system (De la Fuente et al., 2020). In this study we showed that α-tocopherol + VitC in the vaccine formulation shifted a Th1 pattern. But the nature of infiltrating cells in the injection site and depot effect of α-tocopherol + VitC in the vaccine potency and also the vaccine efficacy are some limitations of this study that remain to be clarified.
Conclusion
Our study demonstrated that α-tocopherol + VitC in inactivated SARS-CoV-2 vaccine formulation led to a shift toward Th1, which may be due to affecting the physiology of cells, especially aged ones, and changing their phenotype toward young cells. However, the efficacy of this strategy in preventing mortality in elderly individuals should be assessed in the future.
Footnotes
Acknowledgments
The authors thank the staffs of Vaccine Formulation Laboratory from Department of FMD vaccine production of Razi Vaccine and Serum Research Institute of Iran for technical supports. The authors also are grateful to Borna Zist Pazhohan Knowledge Company and their staff for the technical supports of this project.
Authors’ Contributions
N.A.: Methodology, software, formal analysis, investigation, data curation, and writing—original draft. P.P.: Resources, conceptualization, and visualization. A.K.: Conceptualization, methodology, validation, investigation, writing—review and editing, and visualization. M.T.: Resources, methodology, validation, investigation, and writing—review and editing. Z.A.: Writing—review and editing. M.H.Y.: Funding acquisition, investigation, and writing—review and editing. A.R.S.: Resources, conceptualization, methodology, validation, investigation, and writing—review and editing. M.M.: Conceptualization, methodology, software, validation, formal analysis, investigation, resources, data curation, writing—review and editing, visualization, supervision, and project administration.
Author Disclosure Statement
The authors declare that they have no conflict of interest.
Funding Information
This study has received grant from Recombinant Vaccine Research Center of Tehran University of Medical Sciences (Grant No. 64044).