
Abstract
To develop polyomavirus VP1 recombinant protein-based immunoassay, the expression of two polyomavirus (Karolinska Institute Polyomavirus; KIPyV, and Washington University Polyomavirus; WUPyV) VP1s in insect cells was investigated using an improved baculovirus system (BacMagic). The reliability of the purified VP1 to serve as antigens in serological tests was confirmed by the establishment of an enzyme-linked immunosorbent assay (ELISA). Two panels of serum samples were used, with Panel I comprising 60 sera (20 KIPyV-positive, 20 WUPyV-positive, and 20 negative) and Panel II consisting of 134 sera with unknown status. The seroprevalence of KIPyV and WUPyV in the study population was determined to be 62% and 50%, respectively. Antibody-negative sera exhibited low reactivities in both ELISAs, whereas antibody-positive sera displayed high reactivity with median optical density values of 1.37 and 1.47 in the KIPyV and WUPyV ELISAs, respectively. The differences in seroreactivities between antibody positive and negative for each virus were statistically significant (p < 0.0001; with 95% confidence interval). The study suggests that seroconversion for KIPyV and WUPyV occurs in childhood, with KIPyV seropositivity reaching 70% and WUPyV seropositivity reaching 60% after the age of 5 years. Adult seroprevalence for polyomaviruses was high, with more than 64% and 51% of the adult population being seropositive for KIPyV and WUPyV, respectively. The constant prevalence of KIPyV and WUPyV antibody in the age groups suggested that this antibody persists for life. The fact that antibody titers were generally stable over time revealed a persistent infection of polyomaviruses in the human population. The insect cell-derived recombinant VP1-based ELISA has been demonstrated to be valuable as a serological assay, offering a valid, reliable, fast, nonlaborious, and economical procedure.
Introduction
Various studies have clearly demonstrated the presence of Karolinska Institute Polyomavirus (KIPyV) and Washington University Polyomavirus (WUPyV) in samples from patients suffering from respiratory illnesses (Abed et al., 2007; Abedi Kiasari, 2021; Gaynor et al., 2007). The current understanding of KIPyV and WUPyV, primarily derived from polymerase chain reaction (PCR) detection of viral DNA sequences, is still in its early stages, leaving much to learn about their prevalence, primary infection, disease associations, and diagnostic methods. Despite the progress in polyomavirus molecular diagnostics (Abedi Kiasari et al., 2008; Assis and Carvalho, 2017), serology remains an important alternative method for the diagnosis of viral infections. Rapid serological diagnosis of acute viral infection can be achieved by the detection of virus-specific antibodies. For patients with compromised immune systems, such as those undergoing cancer chemotherapy, the immune response may be absent or delayed. In these cases, it is essential to use highly sensitive serological assays. Enzyme immunoassays (EIAs) utilizing polyomavirus VP1 as the antigen have revealed high immunogenicity of the major capsid protein across BK, JC, and SV40 polyomaviruses. In addition, human serum has been found to effectively detect the recombinant VP1 protein (Hirsch et al., 2013; Moens et al., 2013; Tolstov et al., 2009). In this regard, Chang and colleagues devised an enzyme-linked immunosorbent assay (ELISA) test utilizing recombinant JCPyV VP1, synthesized in Escherichia coli, as the antigen for conducting serological investigations (Chang et al., 1997). Stolt et al. introduced an EIA that utilized VP1 major capsid proteins of JCPyV, as well as BK virus strains AS and SB, and SV40 expressed in yeast. This approach indicates that VLP-based EIAs could benefit epidemiological investigations of these viruses (Stolt et al., 2003). Moreover, Viscidi et al. developed an EIA aimed at detecting serum antibodies against SV40, JC, and BK polyomaviruses. They highlighted the utility of expressing VP1 capsid proteins in insect cells to produce highly purified proteins that mimic many immunological properties of native virions (Viscidi et al., 2003). Nguyen et al. documented approximately 80% seropositivity for WUPyV and around 70% for KIPyV, whereas Neske et al. reported WUPyV IgG seroprevalence at 89% (median age 31 years, 49.4% male) and 67% for KIPyV (Neske et al., 2010; Nguyen et al., 2009).
In this study we aimed to develop a recombinant VP1-based ELISA for the detection of KIPyV and WUPyV. To streamline the availability of appropriate antigens for diagnostic assays and accelerate the production of recombinant polyomavirus VP1 proteins, we investigated the expression of VP1s from these two polyomaviruses in insect cells utilizing an improved baculovirus system. The suitability of the purified VP1 obtained from this system to serve as antigens in serological assays was evaluated by developing an immunoassay specifically designed for detecting polyomavirus-specific antibodies in human sera.
Materials and Methods
Patient characteristics and human sera
This study utilized two distinct panels of serum samples. Panel I consisted of 60 serum samples, categorized into 20 KIPyV positive, 20 WUPyV positive, and 20 negative samples. The positive samples were obtained from patients confirmed to have KIPyV and WUPyV through PCR, within 2–4 weeks after symptom onset. The negative samples were collected from healthy individuals with no respiratory infections in the past year and were PCR confirmed to be negative for both KIPyV and WUPyV.
Panel II comprised 134 serum samples with unknown status. These samples were randomly selected from various age groups based on the age-specific distribution of human polyomavirus antibodies. The median age of these patients was 35 years, with a mean age of 30 years, ranging from 1 week to 65 years (Table 1). The serum samples were collected from April 2017 to September 2018 and stored at −70°C until use. Full informed consent was obtained from all participants, and to maintain confidentiality, the samples were anonymized by renumbering and removing all patient identifiers before analysis.
Age Distribution of Patients
DNA fragment production
The alignment of gene sequences of KIPyV and WUPyV VP1 from multiple isolates previously published in GenBank (http://www.ncbi.nlm.nih.gov/sites/entrez?db=Nucleotide&itool=toolbar) was performed for each virus using BioEdit Sequence Alignment Editor (Ibis Biosciences, Carlsbad, California, USA), and highly conserved regions were identified for each virus from which primer sequences were selected. To identify any homology between primer sequences and all published sequences in GenBank, a NCBI BLAST (Basic Local Alignment Search Tool) (http://www.ncbi.nlm.nih.gov/BLAST/) search was performed. For generation of specific vector overhangs, the sense primer incorporated a 5′-CAGGGACCCGGT sequence, and the antisense primer incorporated a 5′-GGCACCAGAGCGTT sequence. Primers are detailed in Table 2. The DNA region of KIPyV and WUPyV encoding the capsid protein VP1 was amplified through PCR, employing the KIPyV and WUPyV DNA extracted from a clinical sample as the target.
Details of KI and WU Polyomaviruses VP1 Primers. To Generate Specific Vector-Compatible Overhangs, the Sense Primer Incorporated a 5′-CAGGGACCCGGT Sequence and the anti-Sense Primer a 5′-GGCACCAGAGCGTT Sequence. The Tails Are Underlined
Construction of recombinant plasmid
Recombinant plasmids were created by annealing the treated insert with the PIEx/Bac-3 3C/LIC vector (Novagen, USA). Transformation of Escherichia coli (NovaBlue GigaSingles™ Competent, K-12 strain; Novagen, USA) was performed using the annealed complex. Purification of recombinant plasmid was performed using Mobius™ 200 Kit (Novagen, USA). Analysis of purification was done by 1% agarose gel electrophoresis.
Protein expression
The recombinant mixture (1 mL BacVector medium, 500 ng recombinant plasmids, 5 µL Insect GeneJuice, and 100 ng BacMagic DNA) was added to insect cells (1 × 106 cells/dish) and incubated at 28°C. Recombinant baculovirus was harvested and stored at 4°C. A plaque assay was performed to determine the titer of the recombinant baculovirus. For protein expression, suspension cultures of Sf9 cells (1 × 108 cells seeded in 50 mL medium) were infected at an Multiplicity of Infection (MOI) of 10 and incubated at 28°C with shaking at 150 rpm for 72 h.
Protein purification
For 1 × 108 infected cell pellets extraction, 20 mL of CytoBuster reagent (Novagen, USA) was used and then incubated for 5 min at room temperature followed by centrifugation for 10 min at 5000 g. Ni-NTA His•Bind Resin (Novagen, USA) was added to the extracts and loaded on the column, and the retained resin was washed. The protein was eluted with Ni-NTA Elute Buffer. UV spectrophotometrically was used to determine the yield of protein.
Immunofluorescence
Immunofluorescence assay was performed as previously described (Abedi Kiasari, 2021) using 1:10 dilution of each virus positive plasma sample and a dilution of fluorescein-conjugated goat-anti-human-IgG (Invitrogen).
SDS-PAGE, western blot, and dot immunoblot analysis
The expressed and purified proteins were analyzed using Sodium Dodecyl Sulfate Polyacrylamide Gel Electrophoresis (SDS-PAGE), western blot, and dot immunoblot methods as previously described (Abedi Kiasari, 2021). In blotting assays, the membrane was probed with a 1:1500 dilution goat polyclonal to human IgG antibody conjugated to horseradish peroxidase (Abcam, USA), goat polyclonal to human IgG antibody conjugated to alkaline phosphatase (Abcam, USA), or with a 1:1500 of goat polyclonal to rabbit IgG antibody conjugated to alkaline phosphatase (Abcam, USA).
Recombinant VP1-based ELISA
Flat-bottomed 96-well polystyrene microplates (Immulon 2, Dynex Technologies, Chantilly, USA) were coated with the 50 μl volumes containing purified recombinant VP1 protein (0.2 μg) in the coating buffer (Phosphate-Buffered Saline; PBS, pH = 7.2). After incubation at 4°C overnight, the ELISA plate was washed 6 times with PBS-T (0.1% Tween-20) and then blocked with 300 μl volumes of a blocking solution (5% skim milk powder in PBS-T) for 1 h at 37°C. The plates were washed six times; 100 μl sera (diluted 1:40) was added and incubated at 37°C for 1 h. After washing six times with PBS-T, 100 μl volumes of horseradish peroxidase conjugate anti-human IgG (gamma chain specific; Sigma, UK) produced in goat (diluted 1:4000) were added and the plates were incubated for half an hour at 37°C followed by washing six times with PBS-T. Thereafter, 100 μl of 2,2-azino-bis-3-ethylbenzothiazoline-6-sulfonic acid (ABTS) peroxidase substrate tablet (Sigma) dissolved in ABTS buffer solution (10 mL) was added to each well. The plates were incubated in a dark room for 30 min at room temperature. The developed color was detected at wavelength 405 nm by an ELISA reader (Titertek Multiscan MCC/340 MKII, Labsystems). The cutoff to define a positive sample was calculated by subtracting the mean absorbance value of the negative control from the mean row absorbance value read at 405 nm.
Application of the optimized ELISA for serum specimen
Panel I of sera consisting of 20 positive [for each virus] and 20 negative sera were used for validation of ELISA. To examine the presence of KIPyV and WUPyV antibodies in serum samples of patients (Panel II consisting of 123 sera), the optimized VP1-based ELISA was used as described above.
Competitive VP1 binding assay
To determine sera cross-reactivity, samples were preadsorbed using purified VP1 proteins. Initially, various amounts of VP1 protein (0.25, 0.5, 1, 2, 3, and 4 μg/mL) were used to determine the concentration needed to efficiently adsorb cross-reactive antibodies. VP1 proteins (3 μg/mL) were added to sample diluted (1:40) in blocking buffer and incubated for 30 min at 37°C prior to incubating samples with test antigen. Then, sample (100 μl) was added to antigen-coated plate, and the assay was completed as described above. The percent competition was defined as 1 minus the mean optical density (OD) value for wells containing diluent with added VP1 divided by the mean OD value for well with buffer alone.
Statistical analysis
All statistical analysis was performed using SPSS 14.0 software (SPSS Inc., Chicago, USA), including determination of the mean, median, standard deviation (SD), ranges, p-values, and Pearson’s correlation coefficient.
Results
Expression of VP1 in insect cells culture
The major capsid proteins of KIPyV and WUPyV were expressed in the Sf9 insect cells. An indirect immunofluorescence assay was applied to examine Sf9 cells infected with recombinant baculovirus containing the VP1 gene. The fluorescence intensity of the infected Sf9 cells was observed (Panels A-B; Fig. 1). No fluorescence signal was observed in immunofluorescence assay of noninfected insect cells. Furthermore, western blot and dot immunoblotting assays confirmed the antigenicity and identity of the recombinant VP1 proteins. A protein band with an approximate molecular mass of 45 KDa was observed in cells infected with all recombinants. There was no reactivity of sera with the negative control (noninfected Sf9 cells), indicating the specificity of the reaction.

Immunofluorescence assay:
Purification of VP1 protein
Ni-NTA resin columns were used to purify the expressed recombinant proteins from the Sf9 lysate. A high degree (95%) of purification was achieved using this system as observed in SDS-PAGE. The recombinant proteins of the expected size (45 KDa) were achieved (Fig. 2). Western blotting analysis of the lysates of Sf9 cells and purified proteins revealed a band of approximately 45 KDa, indicating the presence of the target protein (Fig. 3). The results of dot blot analysis showed that the expressed proteins were found to react with the virus-specific antibodies (Fig. 4).

Protein expression and purification analyzed by SDS-PAGE. KIPyV (left; Lanes 2 to 12) and WUPyV (right; Lanes 12 to 16) VP1: Lane 1: Mw Marker (EasyLadder, Bioline); Lane 2 KIPyV VP1 flow-through; Lanes 3 to 6: Wash. Lanes 7 and 10 Elute 1; Lanes 8 and 11 Elute 2. Lane 12 Elute 3 WUPyV VP1; Lane 13 Elute 2; Lane 14 Elute 1; Lane 15 Wash; Lane 16 Flow-through.

Western blot analysis of expressed proteins in insect cells. 1: Marker; 2: KIPyV VP1 protein (72 h p.i.); 3. WUPyV VP1 protein (72 h p.i.).
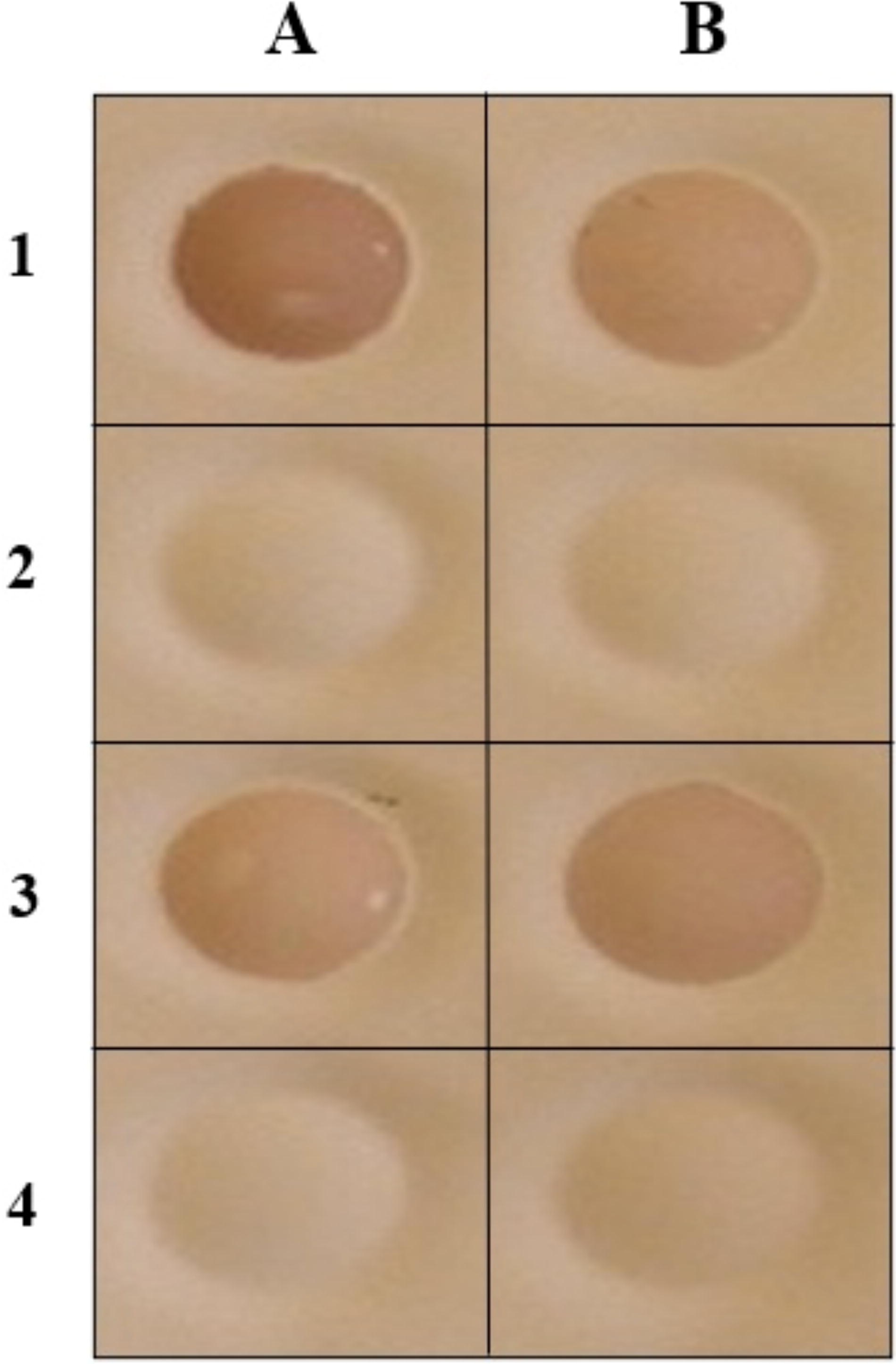
The immunoreactivity of the expressed proteins was examined using dot blot.
Protein yields determination
The yields of purified VP1 proteins from the insect cells (10 mL culture; 107 cells) were 1.67 mg/mL and 1.86 mg/mL for KIPyV and WUPyV, respectively, as analyzed spectrophotometrically.
ELISA development and validation
The purified polyomavirus VP1 proteins were utilized to develop ELISAs for the detection of polyomavirus-specific IgG antibodies. The optimal concentration of polyomavirus VP1 was determined by titration using positive and negative controls, establishing that 200 ng/well was appropriate for the assay. Using a polyomavirus-positive human serum (1:40 dilution), as little as 25 ng/well of purified VP1 protein yielded an ELISA signal at least twofold above the background reading (A405 < 0.2). The standard ELISA protocol was performed with various serum and anti-human IgG conjugate dilutions, and the working system was determined to be a 1:40 serum dilution with a 1:4000 conjugate dilution. Samples with OD values equal to the cutoff were retested, and each plate included duplicates of positive controls, negative controls, and a blank. For validation, a panel of 60 known serum samples (20 KIPyV positive, 20 WUPyV positive, and 20 negative) was used. A violin plot with a box plot overlay (Fig. 5) was created to visualize the OD value distributions from the ELISA for these 60 known serum samples. The KIPyV Positive group had a median OD value of 1.42, whereas the WUPyV Positive group exhibited a median OD value of 1.48, both indicating high seroreactivity. Conversely, the Negative group had a median OD value of 0.24, reflecting low seroreactivity. The violin plots illustrated the full distribution of OD values for each group, with the superimposed box plots highlighting the median, interquartile range, and potential outliers. The individual data points emphasized the clear separation between the OD values of antibody-positive and negative sera for both KIPyV and WUPyV.

Figure displays the distribution of OD values for three groups of panel I of serum samples (n = 60), categorized as KIPyV Positive (n = 20), WUPyV Positive (n = 20), and Negative (n = 20), based on the results of the polyomavirus VP1 recombinant protein-based ELISA. The violin plots illustrate the density of the OD values for each group, whereas the box plots within the violin plots highlight the median, interquartile range, and potential outliers. Individual data points are overlaid for additional clarity. OD, optical density.
The results validated the reliability of the purified VP1 protein as an antigen in serological tests, confirming the assay’s effectiveness in distinguishing between antibody-positive and negative sera. One negative serum was considered positive by ELISA, whereas two positive sera were negative. The sensitivity and specificity of the ELISA were 93.93% and 97.95%, respectively, with statistically significant differences in seroreactivities between antibody-positive and negative sera for each virus (p < 0.0001; 95% confidence interval [CI]).
Cutoff value determination
Cutoff values were determined as follows: the mean value and SD were calculated from 8 replicates of the negative controls. The cutoff value between positive and negative samples was set to be the mean OD value of 8 replicates of the negative control plus three SD values of the mean. Table 3 shows the OD value mean (M) and SD of 8 replicates of KIPyV and WUPyV negative controls and the calculated cutoff value for each antigen. The variability data presented in this table indicate intrarun variability, with observed coefficients of variation (CV) for KIPyV and WUPyV negative controls being 12% and 14.8%, respectively. Although a CV of 12% is considered elevated for intrarun variability, these values provide important context for the consistency and reliability of our assay measurements.
Optical Density Value M and Standard Deviation of Eight Replicates of KIPyV and WUPyV Negative Controls, Representing Intrarun Variability and the Cutoff Value for Each Antigen
OD, optical density.
Antibody cross-reactivity determination using competitive VP1 binding assay
To determine sera cross-reactivity between two VP1 proteins, serum samples were preadsorbed using purified VP1 proteins to investigate potential cross-reactivities in the specific ELISA. To determine the concentration of VP1 protein needed to effectively inhibit antibody reactivity in the VP1-based ELISA, VP1 proteins were titrated. The result of the titration experiment showed that 3 µg/mL of competing VP1 was sufficient to effectively inhibit antibody reactivities. Virus-specific positive sera samples were preadsorbed using virus-specific purified VP1 proteins and then applied to a VP1-coated plate. The plates were processed, and the assay was performed.
For each virus, one positive serum sample that exhibited coincident seroreactivity with other polyomaviruses was tested. No cross-reactivity was observed between the VP1 protein of KIPyV and WUPyV (Panels A and B; Fig. 6).

Seroreactivity to virus specific VP1 antigen after competition with other VP1 antigens.
Comparative age seroprevalence study
Sera from the 134 children and adults were tested for VP1-reactive antibodies using recombinant VP1 from KIPyV and WUPyV. Each sample was tested in duplicate, and samples with an OD value equal to the cutoff values were repeated. Duplicates of positive controls, negative controls, and blank were included in each plate. A seroprevalence of 61.9% for KIPyV and 70.1% for WUPyV was determined (Table 4). The negative sera gave low reactivities in two ELISAs, with mean OD values of 0.23 and 0.25 in the KIPyV and WUPyV ELISAs, respectively. By contrast, the antibody-positive sera gave high levels of reactivity in all five ELISAs, with median OD values of 1.42 and 1.48 in the KIPyV and WUPyV ELISAs, respectively. Statistically significant differences (p < 0.0001; with 95% CI) were found in seroreactivities between antibody positive and negative for each virus.
Age Specific Seroprevalence of Antibody to Polyomaviruses
Discussion
Previous studies, including our own, have identified KIPyV and WUPyV in nasopharyngeal samples from individuals exhibiting respiratory symptoms (Abed et al., 2007; Abedi Kiasari et al., 2008; Gaynor et al., 2007; Han et al., 2007). Despite these findings, the clinical implications of KIPyV and WUPyV infections in humans remain unclear. However, there have been cases where KIPyV was found in patients with severe respiratory symptoms without the presence of other viruses, and instances of WUPyV being linked to bronchitis have been documented (Babakir-Mina et al., 2013; Dehority et al., 2017; Teramoto et al., 2011).
Despite progress in molecular diagnostic techniques, serology continues to be an essential alternative for identifying viral infections. Rapid serological diagnosis of acute viral infections is possible by detecting virus-specific antibodies. Developing sensitive and specific immunoassays may enhance our understanding of the epidemiology and pathogenesis of KIPyV and WUPyV across different clinical specimens. To streamline the production of suitable antigens for ELISA and accelerate the expression of polyomavirus VP1 recombinant proteins, this study examined the expression of VP1 proteins from two polyomaviruses (KIPyV and WUPyV) in insect cells using an enhanced baculovirus system. The recombinant VP1 proteins were characterized through immunofluorescence, SDS-PAGE, western blot, and dot blot analyses.
Analysis of infected cells by immunofluorescence revealed that KIPyV and WUPyV VP1 protein was successfully expressed in insect cells. The fluorescent intensity of the baculovirus infected Sf9 cells showed the immunoreactivity of the expressed protein in insect cells.
The KIPyV and WUPyV VP1 proteins, produced in insect cells, exhibited a molecular mass of approximately 45 kDa as observed in SDS-PAGE analysis. This size is consistent with the molecular weights reported for these recombinant proteins in earlier studies (An et al., 1999; Chang et al., 1997; Gillock et al., 1997).
Western blot analysis of the lysates revealed a 45 kDa band corresponding to VP1. Both western blot and dot blot analyses demonstrated that human serum effectively reacted with the recombinant VP1 proteins.
The VP1 protein produced in the baculovirus system was utilized as an antigen for developing a polyomavirus VP1-based ELISA. To determine the optimal VP1 concentration for plate coating, titrations were conducted with positive and negative controls. The purified VP1 protein was effectively detected using polyomavirus-positive human serum at a 1:40 dilution. As little as 25 ng/well of purified VP1 protein generated an ELISA signal at least twice the background level. These results align with previous studies (Stolt et al., 2003; Viscidi et al., 2003), which indicated that 25–200 ng/well of VP1 protein achieved appropriate saturation of ELISA plate wells. Higher concentrations led to oversaturation, with excess antigen removed during washing. Consequently, 200 ng VP1/well was selected as the optimal concentration for detecting minimal antibody concentrations following positive serum control titration.
To determine the optimal dilutions for human serum and anti-human IgG conjugate, an assay was conducted using serial dilutions of control serum and conjugate. It was found that a serum dilution of 1:40 and a conjugate dilution of 1:4000 provided the best differentiation between positive and negative controls. Therefore, a serum dilution factor of 1:40 and a conjugate dilution factor of 1:4000 were chosen as the optimum dilutions for screening serum samples. Samples were considered positive when the OD reading was greater than the mean OD of eight negative control replicates plus three SDs (Ridge and Vizard, 1993). Using this method, the best discrimination between the negative and positive controls was obtained. Both ELISAs showed perfect specificity when the cutoff value was calculated. The results showed that polyomavirus VP1, when used as antigen in an ELISA, can distinguish between polyomavirus-positive patients and controls. It demonstrated that the antigenic configuration of the VP1 proteins was recognized by specific virus antibodies. For the study of polyomaviruses, the produced VP1 purified proteins should play an important role in the development of immunoassays.
In a preliminary study, the presence of antibodies against the KIPyV and WUPyV VP1 antigens was examined across the general population in Iran, spanning from infancy to 66 years of age. Previous seroprevalence investigations for JC and BK polyomaviruses have used a variety of methods, such as hemagglutination inhibition (HI) (De Stasio et al., 1979; Nastac et al., 1982) and a virus-like particle (VLP) ELISA-based test (Stolt et al., 2003; Viscidi et al., 2003). The HI assay typically requires intact virions or VLPs and may exhibit lower sensitivity compared with EIAs in detecting BK and JC VP1 seroreactivity (Hamilton et al., 2000). Moreover, not all polyomaviruses demonstrate hemagglutination (e.g., SV40), and HI has not yet been assessed for KIPyV and WUPyV. Conversely, a VP1-based ELISA can detect antibodies reactive to all VP1 proteins.
In our study population, the seroprevalence of KIPyV and WUPyV was observed to be 61.9% and 70.1%, respectively. The ELISA tests showed low reactivity in antibody-negative sera, whereas antibody-positive sera exhibited significantly higher reactivity with median OD values of 1.42 and 1.48 for KIPyV and WUPyV, respectively. These differences in seroreactivity between antibody-positive and-negative samples for each virus were statistically significant (p < 0.0001; 95% CI). Previous research by Nguyen et al. reported a seropositivity rate of approximately 80% for WUPyV and around 70% for KIPyV. In addition, Neske et al. documented a seroprevalence of 89% for WUPyV IgG (median age 31 years, 49.4% male) and 67% for KIPyV (Neske et al., 2010; Nguyen et al., 2009).
The findings suggest that antibody acquisition for KIPyV and WUPyV occurs during childhood, with seropositivity rates reaching 75% for KIPyV and 80% for WUPyV by the age of 5 years. In adults, the seroprevalence of these polyomaviruses remains notably high, with over 65% seropositive for KIPyV and over 75% for WUPyV. The consistent presence of KIPyV and WUPyV antibodies across various age groups indicates that these antibodies persist throughout an individual’s lifetime. The stability of antibody titers over time further suggests a persistent infection of polyomaviruses in the human population.
Primary infections with KIPyV and WUPyV are generally asymptomatic or associated with mild respiratory illnesses. These infections typically occur during childhood or early adulthood and are followed by the virus spreading to sites where it can establish lifelong persistent infections (Katona et al., 2022; Motamedi et al., 2012). The cell types and sites of persistence differ among polyomaviruses, with several distinct locations identified for their long-term presence (Imperiale and Jiang, 2016; Katona et al., 2022; Motamedi et al., 2012). It has been suggested that infection of these cells may be involved in dissemination or persistence. The mechanisms responsible for persistent infection and how the immune responses control these persistent infections remain unknown.
In this study, a preabsorption assay was conducted to identify potential cross-reactivities between KIPyV and WUPyV VP1 proteins. To determine the optimal VP1 protein concentration needed to inhibit antibody reactivity in the VP1-based ELISA, a titration study was performed. The titration results indicated that preincubation with 3 µg/mL of VP1 was sufficient to effectively inhibit antibody reactivity in the ELISA.
No cross-reactivity was observed between the VP1 proteins of KIPyV and WUPyV. Similarly, Nguyen et al. conducted preincubation experiments to determine if cross-reactivity exists between antibodies against WUPyV VP1 and those against KIPyV, SV40, BK, and JC in human serum. Their results showed that VP1 proteins from BK, JC, KIPyV, and SV40 only minimally reduced absorbance signals, indicating no significant cross-reactivity with antibodies against WUPyV VP1 (Nguyen et al., 2009). Another study also found no cross-reactivity between the VP1 proteins of lymphotropic polyomavirus or MCPyV and between WUPyV and KIPyV (Kean et al., 2009).
A more detailed investigation into the seroepidemiology of these viruses would likely yield valuable insights into their infection patterns and pathogenesis. The recombinant VP1-based ELISA developed in this study could prove useful for examining the seroprevalence and pathogenesis of polyomavirus infections, developing serodiagnostic tests, and further elucidating the antigenic relationships among members of the polyomaviridae family. However, the limited sample size in this study, due to difficulties in obtaining sufficient positive samples for KIPyV and WUPyV, affected the evaluation of the assay’s specificity and sensitivity. Recognizing this limitation, we have planned future research with larger cohorts to validate our findings.
Footnotes
Authors’ Contributions
B.A.K. and F.H.F. designed the study. B.A.K., M.G., and A.H.A. performed the experiments. A.H.A., and M.G. analyzed the data. B.A.K., and M.G. collected the study data. B.A.K. and A.H.A. wrote the manuscript. B.A.K., M.G., and F.H.F. reviewed the manuscript. F.H.F. supervised the study. All authors read and approved the final manuscript. B.A.K. and M.G. contributed equally to this work and are designated as co-first authors.
Author Disclosure Statement
The authors declare that they have no competing interests.
Funding Information
No funding was received for this article.