
Abstract
Following viral infection, antigen-restricted T lymphocytes are activated and recognize infected cells to eliminate them. A subset of T cells differentiates into memory lymphocytes able to counteract viral rechallenge in a faster and enhanced way. SARS-CoV-2 can escape immune responses leading to a poor clinical outcome. Immune escape can be associated with the failure of the development of T cell memory compartments. The aim of this study is to characterize the T memory subsets and to test the immune response against class I- and II-restricted immunodominant epitopes shared by ancestral and SARS-CoV-2 variants strains. T memory subsets and recognition of SARS-CoV-2S Spike-specific epitopes were analyzed by flow cytometry on 14 fully vaccinated healthy donors (HDV) and 18 COVID-19 recovered patients (CD). The results obtained showed that CD8+ T naïve subset numbers decreased in association with a significant increase of the effector memory T cell subset whereas there was a small increase in the percentage of SARS-CoV-2 antigen-restricted T clones in both CD4+ and CD8+ subset in the CD compared to HDV sample. Collectively, these features may reflect a broader cytotoxic T cell repertoire stimulated by the virus during the natural infection compared to the spike-restricted response activated during vaccination.
Introduction
Following the antigenic challenge, the immune system develops the memory compartment that allows to rapidly activate and expand specific T cell clones against the returning pathogen (Martins et al., 2023; Turner et al., 2021). Memory T cells are organized in various populations to maintain a balance among proliferation, activation, naïve T (TN) cell formation, and time elapsed from the last encounter of the pathogen and to minimize energies usage (Mahnke et al., 2013; Zhan et al., 2017). Memory T cells can be defined by the expression of molecular markers both in CD4+ and CD8+ T subsets (Turner et al., 2021). The central memory T cells (TCMs) express CD45RA–CCR7+CD62L+, they are already primed for the pathogen epitope recognition and they circulate along the secondary lymph nodes to finally find the relevant epitope and rapidly expand in number (Pepper and Jenkins, 2011; Turner et al., 2021). The effector memory T cells (TEMs) express CD45RA–CCR7–CD62L+/– markers, are located inside the peripheral organs, and cannot proliferate. TEMs are immediately able to recognize and attack the specific pathogen (Turner et al., 2021). Effector memory recently activated T (TEMRA) population shows features comparable with effector T cell but it expresses homing markers similar to the TEM subset (CD45RA+CCR7–CD62L+) (Salumets et al., 2022).
SARS-CoV-2 disease (COVID-19) is a pathology caused by the infection of SARS-CoV-2 virus that firstly occurred on Earth at the end of 2019 (Coronaviridae Study Group of the International Committee on Taxonomy of Viruses, 2020). It is characterized by infection through the respiratory tracts, induction of a wide range of symptoms starting from cough, fever, flu, and leading up to pneumonia, and in the most severe forms, cytokine storm and eventually death (Hu et al., 2021). The global spreading of the pandemic produced a rapid evolution of the virus with the rise and fall of different variants that were challenging for the effectiveness of the vaccination (Chavda et al., 2022; Telenti et al., 2022). The antibody production against SARS-CoV-2 worked well along the variants, even if the induction of a sterilizing humoral immunity has not been achieved yet. Some progress has been made in understanding the CD4+ and CD8+ memory response in relation with the time elapsed from the vaccination and the difference between recovered vaccinated subjects and vaccinated subjects (Sette and Crotty, 2022). Nonetheless, the knowledge of the kinetics and interrelationships among types of memory in humans is limited.
Hence, we analyzed memory T cell subpopulations in a cohort of subjects sorted by the time elapsed from the extinction of SARS-CoV-2 infection or from the last dose of vaccine to deeply understand the differences on memory T cells subpopulations distribution.
Materials and Methods
Studying design
In total, 32 subjects have been enrolled for the study: 14 vaccinated healthy donor (HDV) and 18 COVID-19 recovered patients (CD). According to pentamer and tetramer used (see below), all the enrolled subjects were both HLA-A0201+ and HLA-DRB1+ and were stratified depending on the severity of the disease, in the case of CD, and time elapsed from the last vaccination or the negativization following SARS-CoV-2 infection, in HDV and CD, respectively. Characteristics of participants are reported in Figure 1.
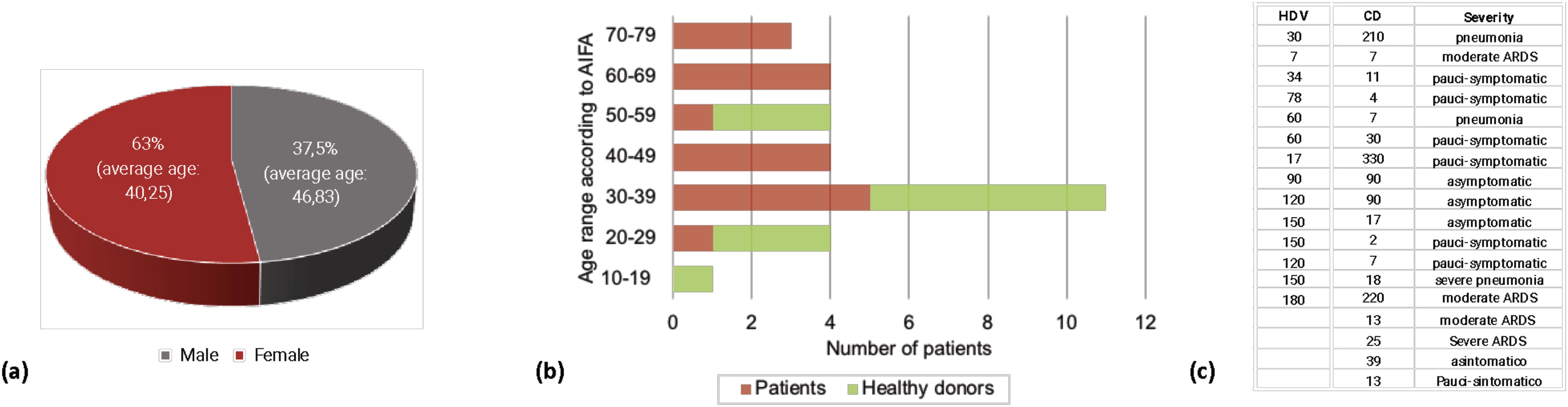
Donors stratification
Immunotyping and memory T CD8+ and CD4+ characterization from the whole blood samples
All subjects gave fasting morning blood samples collected in Ethylenediaminetetraacetic acid (EDTA). A volume of 100 μL of whole blood was incubated for 25 min on ice in the dark with 5 μL of following directly conjugated antibodies (all from Biolegend, San Diego, CA): for Class I and Class II HLA typing, fluorescein-5-isothiocyanate (FITC)-labeled antihuman HLA-A2.1 (clone BB7.2), and phycoerythrin (PE) antihuman HLA-DRB1 (clone NFLD.D2), respectively; for memory subset identification: Peridinin-Chlorophyll-protein (PerCP) antihuman CD4 (clone RPA-T4), PerCP antihuman CD8A (clone RPA-T8), FITC antihuman CD45RA (clone HI100), allophycocyanin (APC) antihuman CD62L (clone DREG-56), and APC/Cyanine 7 antihuman CD197 (CCR7, clone G043H7). In all cases, at the end of the incubation, erythrocytes were lysed by adding 1 mL of lysis buffer (BD Pharm Lyse, Becton Dickinson, Mountain View, CA) to each sample for 15 min at room temperature in the dark. Finally, samples were acquired by using a FACs ARIA II cell sorter (Becton Dickinson, Mountain View, CA) equipped with a 488 nm solid state laser and a 633 nm HeNe laser using FACs DiVa Software (v6.3.1, Becton Dickinson, Mountain View, CA), and analyzed by using FlowJo Software (v10.8.1, Becton Dickinson, Mountain View, CA).
Gating strategy of memory T cells
Lymphocytes were identified from forward scatter (FSC) vs. side scatter (SSC) plot, then CD4+ or CD8+ T lymphocytes have been gated based on FSC vs. PerCP dots. Finally, the memory compartment was analyzed by gating TN (CD45RA+CCR7+CD62L+), TCM (CD45RA-CCR7+CD62L+), TEM (CD45RA-CCR7-CD62L+/−), and TEMRA (CD45RA+CCR7-CD62L+) lymphocytes on both CD4+ and CD8+ T subsets. See Supplementary Figure S1 for the complete depiction of the gating strategy.
Pentamer and tetramer staining
The Pro5 MHC Class I Pentamer-PE allows to recognize HLA-A2.1 restricted CD8+ T cells specific for the SARS-CoV-2 S-protein FIAGLIAIV epitope (residues 1220–1228), whereas the ProT2 MHC Class II Tetramer-PE allows to recognize HLA-DRB1 CD4+ T cells specific for the SARS-CoV-2 S-protein QLIRAAEIRASANLAATK epitope (residues 993–1010, Proimmune, Oxford, UK). We chose these epitopes according to scores obtained using the SYFPEITHI tool (www.syfpeithi.de), following the manufacturer advices. In particular, the following score were attributed: FIAGLIAIV score 30; QLIRAAEIRASANLAATK score 18.
Once established the HLA subtype of the sample as already described, a volume of 100 μL of whole blood was analyzed with 10 μL of Pro5 MHC Class I Pentamer-PE by incubating for 60 min at 4°C, whereas 5 μL of ProT2 MHC Class II Tetramer-PE was incubated for 120 min at 37°C on another 100 μL of whole blood of the same sample. The red blood cells lysis procedure, acquisition, and analysis were performed as already described.
Gating strategy of pentamer and tetramer
Total lymphocytes were identified by Forward/Side Scatter proprieties. T lymphocytes were further characterized as CD4+ and CD8+ cells based on FSC vs. PerCP dots. Finally, CD4+ lymphocytes able to recognize ProT2 MHC Class II Tetramer-PE and CD8+ lymphocytes able to recognize Pro5 MHC Class I Pentamer-PE have been enclosed into the CD4 CoV+ or CD8 CoV+, respectively. As negative control and to correctly set the gate corresponding to CoV+ positive events we used Pro5 MHC Class I Pentamer-PE and ProT2 MHC Class II Tetramer-PE specific for HIV-1 peptides, being all the enrolled subjects seronegative. See Supplementary Figure S1 for the complete depiction of the gating strategy.
Statistical analysis
Results were presented as median values. The difference between two groups has been calculated using the nonparametric Mann–Whitney U test. When comparing more than two groups, the nonparametric Kruskal–Wallis test followed by the Dunn post-test was used to compare all pairs of data. GraphPad PRISM v5.0 (GraphPad Software, Inc., San Diego, CA, USA) and Excel v16.43 (Microsoft, Redmond, WA) were used for the statistical analysis. p < 0.0001(***); p < 0.01 (**); p < 0.05 (*). Error bars in the graphs represent the standard deviation. To compare CD4+ and CD8+ memory cell subsets as a function of the time, we performed a descriptive of the number of CD4 and the CD8 cells according to each type of cells (LN, TCM, TEM, and TEMRA) calculating their medians and interquartile ranges (IQR) in each study group, COVID and non-COVID. We considered the measurements collected from a minimum of 0.5 to a maximum of 6 weeks since the start of convalescence for COVID, and since the date of the 3rd dose for non-COVID group, respectively. We assessed the differences between the two groups (COVID vs. non-COVID) by nonparametric Mann–Whitney U test (Supplementary Fig. S2).
Results
Experimental design and recruitment of the patients
This study was conducted by enrolling 32 subjects of which 63% were female and 37% male. The average ages were 40 and 47 years old for female and male subjects, respectively (Fig. 1a). By sorting the subjects by their ages, most of the subjects fell in the 30–39 years age range (Fig. 1b). All the subjects were mainly characterized for the elapsed time from the last vaccine dose (in the case of the HDV) or the day of the absence of SARS-CoV-2 virus counts (in the case of the COVID-19 recovered subjects, Fig. 1c). The elapsed time from hospital dismission ranged from 2 to 330 days (median: 17.5 days; IQR: 69.25), whereas in the case of vaccinated individuals ranged from 7 to 180 days (median 120 days: IQR: 116). Contextually, all the patients were defined based on the severity of the COVID-19 disease. Clinical conditions varied from asymptomatic to severe pneumonia COVID-19 disease (Fig. 1c).
COVID-19 convalescents display expanded CD8+ effector memory T cell and decreased CD8+ naïve T subset compared to HDV
It has been reported that cell mediated immune responses against SARS-CoV-2 include all the viral products, especially M and N proteins (Grifoni et al., 2020). Then, we sought to compare the percentage of both CD4+ and CD8+ T lymphocyte memory subsets in recovered subject (CD) to HDV. The features of T memory response could be, indeed, representative for the difference between HDV and CD. As shown in Figure 2a, CD4+ and CD8+ T lymphocytes were identified from all of the lymphocytes. Subsequently, both CD4+ and CD8+ T lymphocytes were clustered following CD45RA, CCR7 and CD62L expression in four memory T cells populations: NT CD45RA+CCR7+CD62L+, TCM CD45RA-CCR7+CD62L+, TEM CD45RA-CCR7-CD62L+/-, and TEMRA CD45RA+CCR7-CD62L+. CD4+ memory T cells populations did not show difference between healthy donors and recovered subjects (i.e., TN HDV: 7.53 ± 12.07% vs. CD: 5.27 ± 7.09%; TCM HDV: 2.34 ± 3.87% vs. CD: 2.54 ± 4.64%; TEM HDV: 57.76 ± 19.57% vs. CD: 61.45 ± 19.1%; TEMRA HDV: 32.54 ± 19.13% vs. CD: 26.99 ± 15.33%), whereas CD8+ Memory T cells showed downmodulation of NT population (i.e., HDV: 49,76 ± 16,49% vs. CD 19,72 ± 14,76%, p < 0,0015) and upregulation of TEM population in CD compared to HDV subjects (i.e., HDV: 24,87 ± 13,72% vs. CD 46,19 ± 17,51%, p < 0,0074, Fig. 2b).
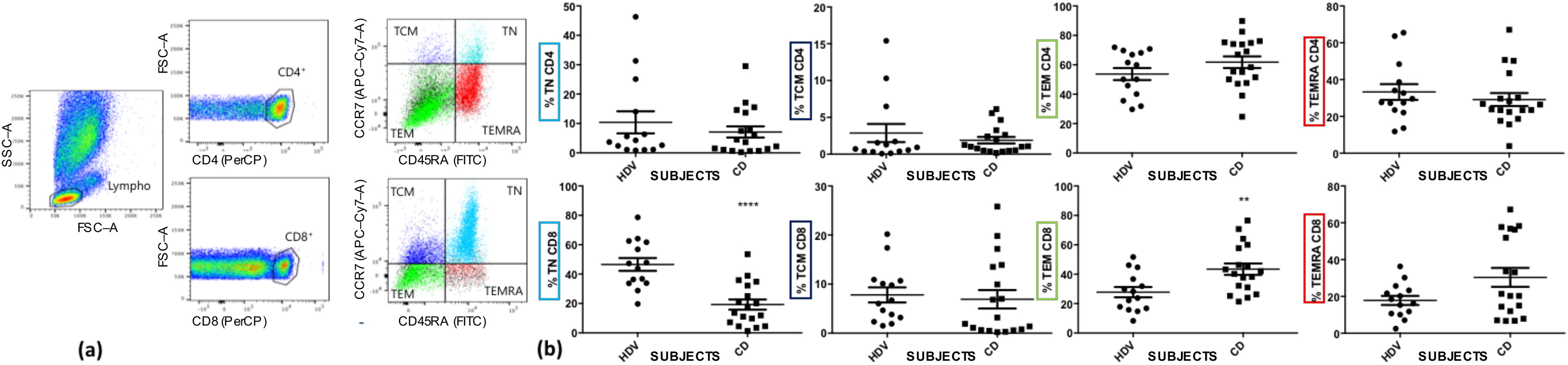
Memory T cells characterization
When we stratified the data looking at the absolute cell number of both CD4+ and CD8+ subsets and subpopulations, we did not observe any statistically significant difference between COVID vs. non-COVID group. Of note, we found the greatest differences when comparing CD8+ cells of COVID with non-COVID group, with higher medians in the COVID group for the three type of cells: TCM, TEM, and TEMRA (Supplementary Fig. S2).
CD4+ and CD8+ derived SARS-CoV-2 antigen restricted T clones similarly respond in both CD and HDV
SARS-CoV-2 virus infects target cells mainly through the interaction between its S-protein and the ACE2 receptor on the surface of pulmonary mucosa (Hoffmann et al., 2020). The pivotal role of S-protein in the initial phase of the viral cycle is confirmed by the generation of different types of vaccines against this protein. S-protein degradation by immunoproteasome generates a great number of immunological epitopes (Chatterjee et al., 2020). The most immunogenic and conservative epitope was selected to be assayed by cytometric analysis (see Material and Methods for details). Starting from the peripheral blood of all the previously described subjects, a protocol of CD4 or CD8 T cell restricted against the S-protein chosen epitope selection was executed (CoV+ CD4 and CoV+ CD8) (Fig. 3a). By analyzing the percentage of CoV+ CD4 it is interesting to observe that 6 over 17 recovered subjects we analyzed showed an increase higher than 0,25% while none of the vaccinated healthy donors were able to pass this threshold (Fig. 3b). Concerning the CoV+ CD8 just 1 patient showed levels higher than 0,75% while all the other subjects did not move significatively from zero (Fig. 3b). Collectively, these results suggest that, even if CD4 T cells seems to more easily recognize selected SARS-CoV-2 epitopes than CD8 T cells, such behavior is not significantly different between recovered patients and vaccinated healthy donors.

Donors stratification and SARS-CoV-2 epitope recognition.
Vaccination and recovering from disease induce superimposable T memory response
To define whether the vaccinated control subjects and recovered patients could be effective to develop T memory response difference, a principal component analysis (PCA) was performed by considering the elapsed time from vaccination or recovery from COVID-19 disease, the type of vaccine, age, and gender, comparing HDV and CD subjects and demonstrating that no differences could be determined (Fig. 4a). In Figure 4b, the comparison between HDV and CD subjects in relation to CD4 CoV+ and CD8 CoV+ values is shown, and a strong superimposition between the two type of subject can be observed. Considering all of the values regarding the abundance of the different subpopulations of CD4+ and CD8+ memory T cells, an expansion of variance along PC1 and PC2 is evident, although the superimposition among HDV and CD subjects is maintained (Fig. 4c).

Principal component analysis (PCA) The PCAs have been performed to evaluate the variance among HDV and CD derived data
Discussion
It is already established that a permanent neutralizing humoral immunity is non achievable after natural infection as well as vaccination with Spike-based SARS-CoV-2 vaccines (Beukenhorst et al., 2023), much less is known about the development of the T cell memory compartment during both the natural infection and the vaccination procedures. Since the beginning of the SARS-CoV-2 of pandemics it has been already demonstrated that a SARS-CoV-2 restricted response is detectable in both CD4+ and CD8+ T lymphocytes, especially against peptides derived from M and Spike proteins (Grifoni et al., 2020). It has also been reported that, in the case of CD4+ cells, a cross-reactivity exists between SARS-CoV-2 and common cold coronaviruses (Grifoni et al., 2020). Our work tried to elucidate the development of memory subsets during the recovery phase in naturally infected people as well as in the vaccinated subjects after the completion of the vaccination schedule. In our cohorts no differences were observed in the CD4+ memory subsets in terms of overall percentage as well as spike-specific clones measured by epitope-restricted response through the use of tetramers, as discussed below. This suggests that the vaccination procedure is able to evoke a memory immunity superimposable to the natural infection. On the other side and most remarkably, when we looked at the cytotoxic CD8+ T lymphocyte, a clear difference appeared when we measured the percentage of circulating TEM cells: in the recovered patients, indeed, this cellular subset is greatly amplified compared to vaccinated individuals. When compared to the percentage of TN and TEM cells measured in only three subjects, we were able to enroll, which were uninfected and unvaccinated, the percentage of TN cells ranged between 64 and 76% and TEM between 14% and 15% (data not shown), indicating that TEM subset is amplified in vaccinated individuals and even more in convalescent subjects. Dan et al. have been reported that in COVID-19 recovered patients, the majority of memory CD8+ cells had a TEMRA phenotype with smaller populations of both TEM and TCM, with no differences between hospitalized and asymptomatic subjects (Dan et al., 2021). This might lead us to hypothesize that in an early phase of the T response TEM CD8+ subset appears amplified. Indeed, in our study all the infected subjects were hospitalized and most of them suffered from moderate to severe COVID-19 whereas in Dan et al., the subjects were mostly asymptomatic or paucisymptomatic without requiring hospitalization.
The difference in the percentage of CD8+ TEM cells in convalescent subject vs. vaccinated individuals, could be also ascribed to the ability that other viral proteins than Spike are able to activate specific cytotoxic T lymphocytes. It has been reported, indeed, that despite the immunodominance of T cell reactivity vs. the Spike protein (about 26%), a sizeable response of 22%, 15%, 10% and 7% could be measured from M protein, nsp6, ORF8, and ORF3A, respectively (Grifoni et al., 2020). Conversely, the fact that the immunodominant reactivity of CD4+ cells is mainly directed against M, N, nsp3, nsp4, and ORF8 rather than Spike protein (Grifoni et al., 2020), might explain why no differences between convalescent vs. vaccinated subjects were observed in TEM cells as well as in the other memory subsets.
Unbiased PCA based on immunophenotypic characterization of memory T cells was already used to discriminate from acute, moderate, or severe infection in convalescent subjects and healthy individuals (Sekine et al., 2020). We then tried to test if differences in PCA analysis could be evident in vaccinated vs. convalescent subject finding no neat differences. These results confirmed the ability of the vaccination to induce a really similar even if not completely superimposable immune response in term of induction of T cell memory subsets. The lack of a sizeable unvaccinated and uninfected group did not allow us to conclude if differences exist between naïve individuals (i.e., people neither exposed to virus nor vaccinated) vs. subjects immunologically “experienced” by either natural infection or vaccination.
We also analyzed the presence of spike-restricted CD4+ and CD8+ T cell clones. Again, no statistical differences were observed between vaccinated vs. convalescent individuals whereas no spike-restricted T cell clones were detected in the sample from the three unvaccinated and uninfected subjects available. This suggest that, despite the presence of cross-reactive T cell clones to other Coronavirus species (Grifoni et al., 2020), spike-derived immunodominant epitopes are SARS-CoV-2 specific and could be presented by APC even following the vaccination.
In conclusion, our results suggest that even if largely superimposable, the T cell memory response evoked by natural infection differs especially for CD8+ lymphocytes in naturally infected patients compared to vaccinated subjects.
Nonetheless, some limitations in our study should be underlined: (1) the small number of subjects in our cohorts; (2) the lack of a statistical relevant group composed of unvaccinated and unexposed individuals, due being to the sampling performed starting from late 2021; and (3) the use of only immunodominant peptides derived from Spike and conjugated to fluorescent-labeled tetramers/pentamers, rather than a pool of tetramers/pentamers associated with peptides covering also other viral proteins and patients with other haplotypes than A2.1 and DRB1 HLA-I and -II.
Footnotes
Authors’ Contributions
M.I., R.M.M.: Methodology, Software, Formal analysis, Investigation, Data Curation, Writing–original draft, Writing–review & editing; A.P.: Methodology, Investigation, Writing–original draft; B.K., A.C., R.M., Giu.M., C.D.B.: Methodology, Data Curation, Writing–original draft; L.G.: Writing–review & editing; M.D.: Software, Formal analysis, Data Curation, Writing–original draft, Writing–review & editing; G.R., G.M., M.L.: Conceptualization, Funding acquisition, Supervision, Writing–original draft, Writing–review & editing. All authors have read and agreed to the published version of the manuscript.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
Supported by
Supplementary Material
Supplementary Figures S1
Supplementary Figures S2
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.