
Abstract
Respiratory syncytial virus (RSV) is one of the most important etiologies of acute respiratory infections that cause bronchiolitis in children under 5 years of age. Treatments are expensive, no vaccine is available, and this is an important cause of hospitalization. Costimulatory molecules have been reported to be good inducers of antiviral type 1 immune response. This study aimed to generate a recombinant vaccine with the RSV F protein fused to 4-1BBL to evaluate the activation of an antiviral response in vitro and the production of neutralizing antibodies in vivo. The codon-optimized F gene was subcloned into an expression vector as follows: streptavidin core, gene F, and costimulatorytumor necrosis factor receptor superfamily member 9 -TNFRS9- ligand (4-1BBL). After the induction of expression in Escherichia coli C43, the recombinant protein (SA-F3x-4-1BBL, denominated SF4) was purified and verified by western blotting. Cultured RAW264.7 macrophages were stimulated with SF4 protein, then tumor necrosis factor (TNF) receptor-associated factor 2 (TRAF2), p38, and nuclear factor κ-light-chain-enhancer of activated B cells (NF-κB) were analyzed by western blot, and mRNA cytokines were analyzed by RT-qPCR. Finally, male C57BL/6 mice were inoculated with SF4, and the generation of anti-RSV neutralizing antibodies and serum cytokines was examined. SF4 had a size of 84.4 kDa with a 5.6% yield. SA-F-4-1BBL upregulated TRAF2, TNF-α, and interferon (IFN)-γ expression levels and activated p38 mitogen-activated protein kinase and NF−κΒ pathways in RAW264.7 cells. Importantly, antibodies capable of neutralizing RSV infection and producing type 1 cytokines were detected in the sera of immunized animals. These results suggest that the fusion protein SF4 activates the 4-1BBL signaling pathway, resulting in an effective antiviral response mediated by neutralizing antibodies and antiviral cytokines.
Introduction
Respiratory syncytial virus (RSV) is one of the most frequent causes of acute respiratory infections in children under 5 years of age and in immunocompromised adults. This inflammatory response predisposes patients to allergic sensitization and reinfections during childhood (Lotz et al., 2013; Piralla et al., 2023). The first vaccine consisted of a formalin-inactivated RSV (Burnett’s strain). However, the immune response worsened inflammation in vaccinated children (Chin et al., 1969). There is no vaccine or treatment available for RSV (Sun et al., 2024).
RSV is a member of the family Pneumoviridae. Its genome is a nonsegmented negative-sense RNA encapsulated and lipidic enveloped with transmembrane glycoproteins: the fusion (F) protein, attachment (G) protein, and small hydrophobic (SH) protein (Borchers et al., 2013; Boyoglu-Barnum et al., 2019). The F protein is relatively conserved, highly immunogenic, and displays two molecular conformations, prefusion and postfusion, with almost equal antigenicity that includes the antigenic sites I–IV (Ruckwardt et al., 2019). Of particular interest, antibodies targeting site II exhibit a strong neutralizing character (Andreano et al., 2021). The monoclonal antibody Palivizumab (Synagis®) binds to antigenic site II and is the mainstay therapeutic and prophylactic approach in high-risk patients (Narayan et al., 2020; Ye et al., 2018).
During antigen presentation, costimulatory signals facilitate crosstalk between T cells and antigen-presenting cells to elicit adaptive immunity (Hussell et al., 2004). Among these signals, the 4-1BB (tumor necrosis factor receptor superfamily member 9—TNFRS9—or CD137)–4-1BBL pathway has been shown to activate monocytes/macrophages (Stoll et al., 2021). The receptor 4-1BB is broadly expressed on dendritic cells, macrophages, and B lymphocytes (Melero et al., 2008; Wang et al., 2009), and the binding of 4-1BBL results in the aggregation of TNF receptor-associated factor 1 (TRAF1) and TRAF2 proteins, which activates the nuclear factor κ-light-chain-enhancer of activated B cells (NF-κB) and mitogen-activated protein kinase (MAPK) pathways, resulting in the secretion of type 1 cytokines (Sharma et al., 2014).
The pathogenesis of RSV and airway hyper-responsiveness are associated with the excessive production of type 2 cytokines, such as interleukin (IL)-4, IL-5, and eotaxin, leading to an imbalance among type 1/2 cytokines (Becker, 2006; Qin et al., 2019; Román et al., 1997; Russell et al., 2017). Therefore, type 1 immune response may attenuate severe bronchiolitis. Currently, the use of vaccines fused to ligands is a promising alternative. For instance, the use of the core subunit of streptavidin (SA) fused to 4-1BBL promotes antitumor type 1 responses (Moyle, 2017; Garza-Morales et al., 2019; Powell et al., 2015; Martinez-Perez et al., 2021).
In the present study, we aimed to generate a recombinant protein with the SA core, a pre-F protein with a tandem of three repeats of site II, and 4-1BBL. This recombinant protein (denominated SF4) was tested in vitro to evaluate the activation of the 4-1BBL pathway in macrophages and in vivo for the generation of type 1 cytokines and RSV-neutralizing antibodies.
Materials and Methods
Cells, virus, and plasmid construction
Vero and RAW 264.7 cells (kindly provided by Dr. Loera-Arias from the Autonomous University of Nuevo Leon) were cultured in Dulbecco's Modified Eagle Medium (DMEM) supplemented with 10% heat-inactivated fetal bovine serum and 0.1% penicillin-streptomycin solution at 37°C with 5% CO2. RSV strain A2 (VR-1540) was purchased from ATCC (Manassas, VA, USA) and was propagated in Vero cells. The pUC57-F3x plasmid with a codon-optimized F sequence with three tandems of three repeats of site II was obtained from GenScript Biotech (Piscataway, NJ, USA). pET-6xHN-SA-F3x-4-1BBL was constructed as shown in Figure 1.
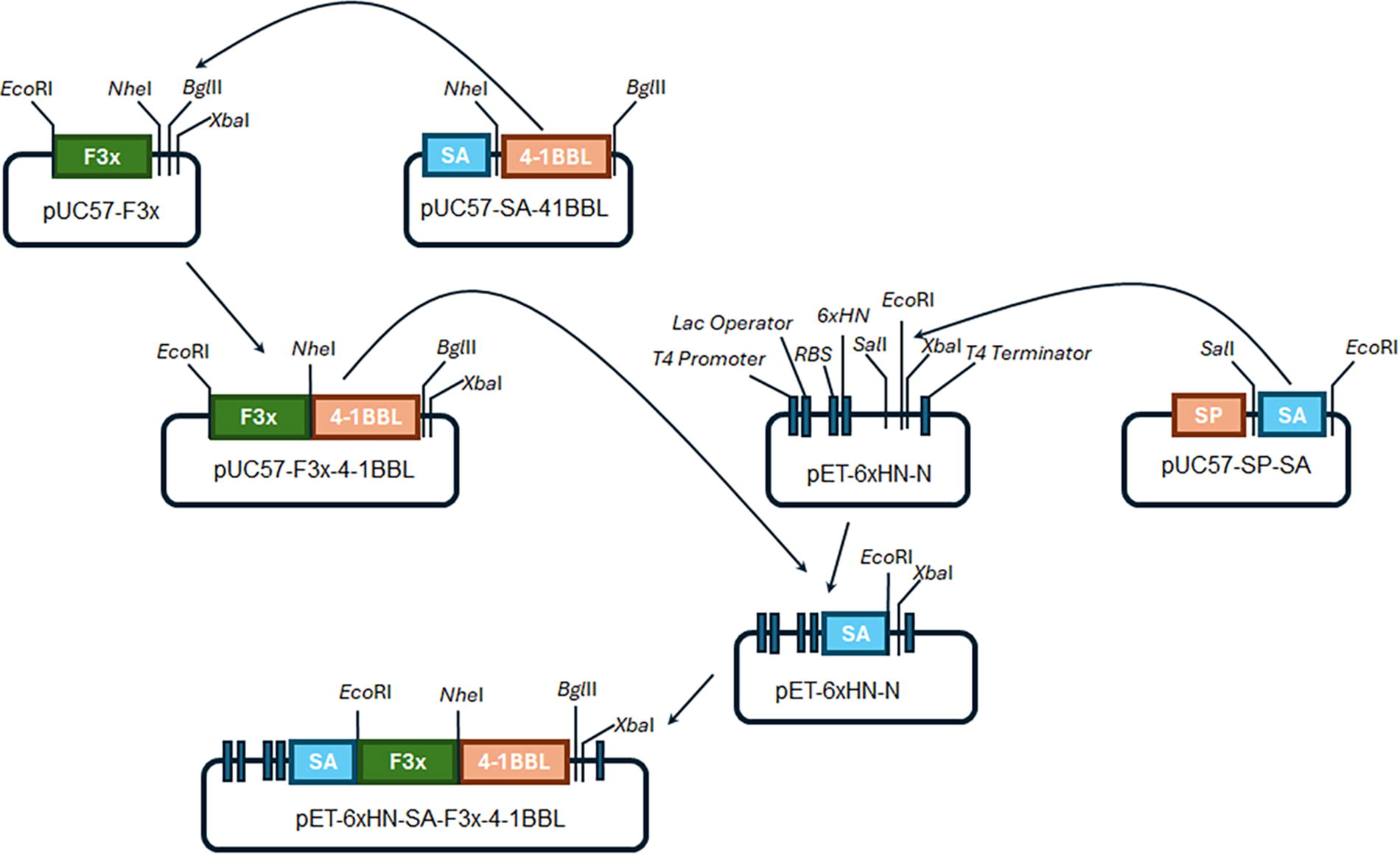
Design of the expression plasmid pET-6xHN-SA-F3x-4-1BBL. The 4-1BBL sequences (669 pb) was obtained after digestion of the plasmid pUC57-SA-4-1BBL with the restriction enzymes NheI and BglII and inserted in the same sites on the pUC57-F3x using the T4 DNA ligase to generate the pUC57-F3x-4-1BBL plasmid. Besides, the SA sequence (381 pb) was obtained from the restriction of pUC57-SP-SA with the enzymes SalI and EcoRI (and ligated in the same sites of the pET-6xHN-N) to generate the pET-6xHN-SA. Finally, the pET-6xHN-SA-F3x-4-1BBL was constructed by ligation of fragment F3x-4-1BBL, which is obtained by digestion of the pUC57-F3x-4-1BBL with EcoRI and XbaI enzymes, into the same site in the pET-6xHN-SA.
Protein production and purification
For the bacterial expression of recombinant SF4, the plasmid pET-6xHN-SA-F3x-4-1BBL was used to transform Escherichia coli C43 (DE3, a gift from Dr. Rocha-Pizaña, Tecnológico de Monterrey). Heterologous expression was induced with 1 mM IPTG (Zymo Research, Irvine, CA, USA). Protein purification was performed using His-Spin Protein Miniprep (Zymo Research). Proteins were separated using 13.5% sodium dodecyl-sulfate polyacrylamide gel electrophoresis (SDS-PAGE) and stained with Coomassie Brilliant Blue solution. The protein production yield was calculated by densitometry using the Gel Analyzer tool of ImageJ software [(intensity of purified SF4 band/intensity of total protein bands) × 100] (Alonso Villela et al., 2020). The purified protein was confirmed using western blot analysis.
Western blot
The proteins were transferred onto polyvinylidene difluoride membranes (Merck Millipore, Darmstadt, Germany) and blocked with bovine serum albumin 5% in tris-buffered saline (BSA-TBS). The corresponding primary and secondary antibodies (Table 1) were incubated. The peroxidase activity was detected using Clarity Western ECL Substrate (Bio-Rad, Hercules, CA, USA) in a Microchemi 4.2 chemiluminescence system (DNR Bio-Imaging Systems, Modi’ in-Maccabim-Re’ut, Israel).
Antibodies Used in This Study
Santa Cruz Biotechnology, Santa Cruz, CA, USA; Sigma-Aldrich, St. Louis, MO, USA; Abcam, Cambridge, UK.
Cell viability assay
RAW 264.7 cells were plated at 1 × 105 cells/well in a 96-well plate and exposed to 0–10 µg/mL of SF4 for 24 and 48 h, and cell viability was quantified by MTT (3-(4,5-dimethylthiazol-2-yl)-2,5-dipheniltetrazolium bromide) assay (Sigma-Aldrich, St. Louis, MO, USA) by measuring at 595 nm in a SmartSpec PLUS (Bio-Rad) and calculated as follows: Abs595 treated cells/Abs595 control cells × 100%.
In vitro activity of SF4
To analyze the response of RAW 264.7, cells after stimulation with the SF4 protein, 5 × 106 cells were cultured in 6-well plates and stimulated with 0.1, 0.5, and 1 µg/mL of SF4 for 24 and 48 h and then processed for protein or RNA extraction.
Total and microsomal protein purification
RAW 264.7 cells were resuspended in 50 µL RIPA buffer (50 mM Tris, 150 mM NaCl, 0.5% deoxycholate, 0.1% SDS, 1% Triton X-100, and 1:50 protease inhibitor cocktail), incubated for 30 min at 4°C with shaking, and centrifuged at 8,800×g for 20 min at 4°C. The supernatant containing total protein was stored at −20°C. To obtain the microsomal fraction, the cell pellet was resuspended in 500 µL of sonication buffer (50 mM Tris-HCl, 1 mM EGTA, 2 mM PMSF, and pH 7.2), sonicated for two periods of 10 s, and ultracentrifuged at 150,000×g for 30 min at 4°C in an Optima MAX-XP Ultracentrifuge (Beckman Coulter, Brea, CA, USA). Proteins were separated using 13.5% SDS-PAGE for western blot analysis.
Analysis of gene expression by RT-qPCR
Total RNA was extracted from RAW 264.7, using the TRI reagent (Sigma-Aldrich), and evaluated using the NanoDrop 2000 (Thermo Scientific, Waltham, MA, USA). cDNA was synthesized using the First Strand cDNA Synthesis Kit (Thermo Scientific). RT-qPCR was performed using Maxima SYBR Green/ROX qPCR Master Mix (2X) (Thermo Scientific) in a StepOne Real-Time PCR system (Thermo Scientific). The oligonucleotides that were used are listed in Table 2. The mRNA expression levels were analyzed using the 2−ΔΔCt method (Schmittgen and Livak, 2008).
Oligonucleotides Used in This Study
Animal model
All animal procedures were approved by the Research Ethics Committee of the
For mouse immunization, 10, 20, and 50 µg of recombinant purified SF4 in 100 µL of elution buffer (50 mM PBS pH 7.7, 300 mM NaCl, and 250 mM imidazole) was instilled intranasally on days 0 and 7. The control animals received elution buffer. The mice were euthanized with overdoses of sevoflurane (PiSA, CDMX, and MX) on day 14, and blood was obtained by intracardiac puncture to determine neutralizing antibodies, inhibition of viral replication, and cytokine concentrations.
Neutralizing antibodies
The titer of neutralizing antibodies was determined following the method described by Bergeron et al. (2023) and Huang et al. (2010) with modifications. Heat-inactivated serum samples were diluted with PBS (1:2, 1:4, 1:8, and 1:16). Then, each sample was incubated with 10,000 PFU of RSV in DMEM at 37°C for 2 h and added to a monolayer of 10,000 Vero cells for 2 h for a final multiplicity of infection of 1. Mix was replaced with supplemented DMEM and incubated for 5 days. The titer of neutralizing antibodies was determined as the highest dilution without cytopathic effects and was evaluated by two blinded observers.
Inhibition of viral replication
After viral titer evaluation, total RNA was extracted using the ZR Viral DNA/RNA kit (Zymo Research). RNA was reverse transcribed using the First Strand cDNA Synthesis Kit (Thermo Scientific). RT-qPCR was performed using Maxima SYBR Green/ROX qPCR Master Mix (2×) (Thermo Scientific) in a StepOne Real-Time PCR system (Thermo Scientific). Oligonucleotides were hybridized in NS1 and NS2 genes (Table 2). For absolute quantification, the plasmid pJET1.2-NS1-NS2 with amplicon NS1-NS2 (270 bp) was constructed, and serial dilutions were prepared with 102–108 copies per reaction. The number of copies in each sample was calculated using the Ct values of the standard curve.
Serum cytokines
Multiplex measurement of cytokines TNF-α, IFN-γ, IL-5, IL-4, and IL-2 in serum samples was performed using the BD cytometric bead array mouse Th1/Th2 kit (BD Biosciences, Franklin Lakes, NJ, USA). The samples were acquired using a FACS CANTO II flow cytometer with a 4-2-2 configuration (BD Biosciences). FCS 3.0 files were recovered and analyzed in FCAP array software V 3.0 (BD Biosciences).
Statistical analysis
All data were analyzed with GraphPad Prism 8.0 software (GraphPad Software Inc., La Jolla, CA, USA) and presented as the mean ± standard error of the mean. Groups were compared using one-way analysis of variance (ANOVA) with Dunnett’s test and Student’s t test for cytokine evaluation. Two-way ANOVA with Tukey’s test was performed to analyze viral replication. p < 0.05 was considered statistically significant.
Results
The coding sequence of the F gene was obtained from GenBank entry U39661. The gene sequence was codon-optimized and analyzed to identify site II (Lee et al., 2022), in which two extra repeats were added in tandem. The codon adaptation index was adjusted to 0.96 for expression in E. coli (Fig. 2A) and %GC to 51.09% (Fig. 2B). The final optimized F sequence with the addition of two extra sites II is shown in Figure 2C. After IPTG induction, the recombinant protein production yield was 5.6% (Fig. 2D). Western blotting with anti-F RSV revealed a protein band of 85 kDa in the soluble fraction (Fig. 2E).
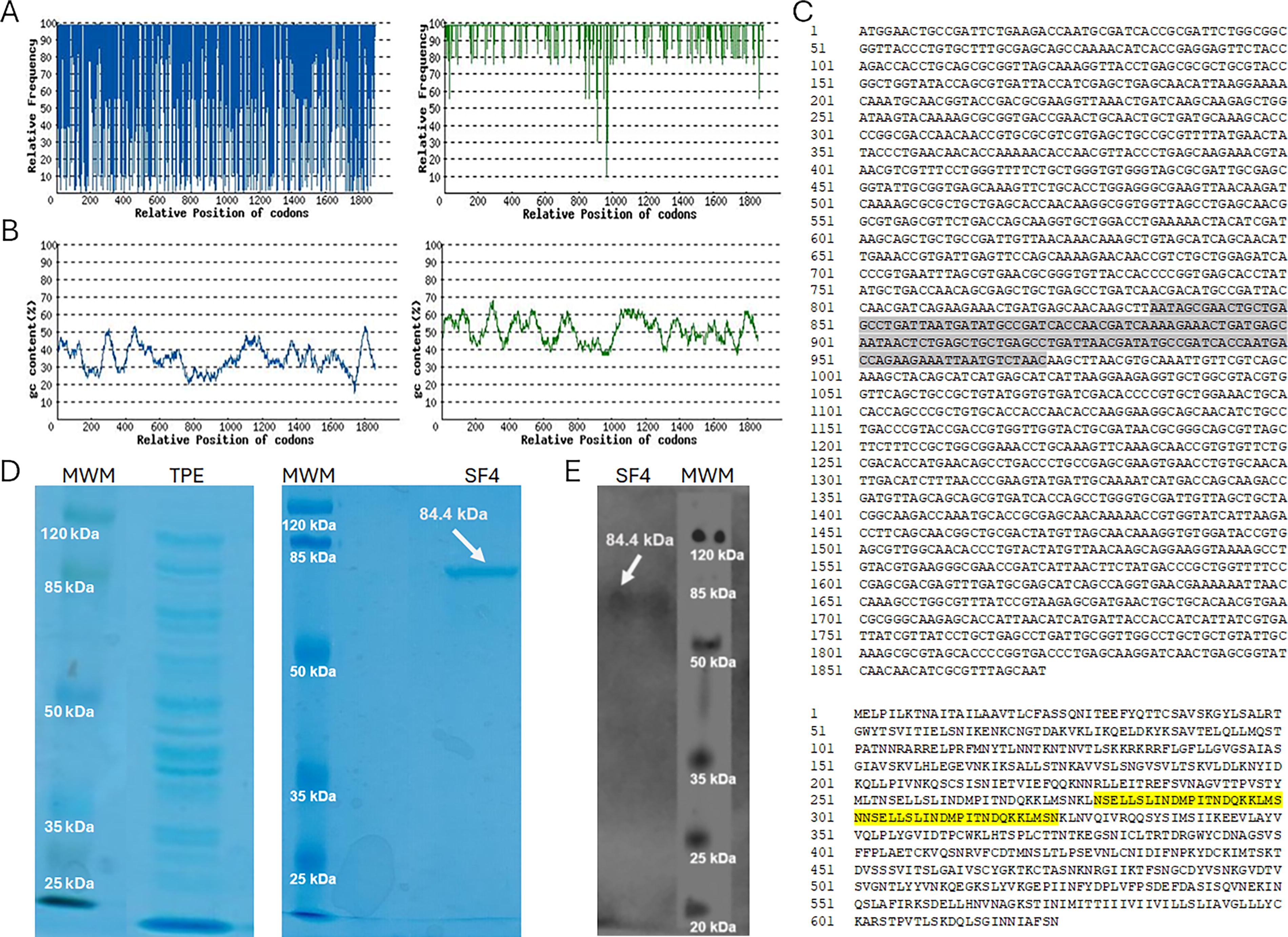
Design and production of SF4 protein.
To investigate the effect of SF4 on viability, RAW 264.7, cells were treated with SF4 protein for 24 and 48 h to identify the concentrations that maintained at least 80% of cell viability. At concentration from 0.1 to 2 µg/mL SF4 at 24 h, cell viability was comparable to that of the control, with at least 88% cell viability, while concentrations from 3 to 10 µg/mL were cytotoxic (Fig. 3A). After 48 h, viability was not affected with 0.1–1 µg/mL of SF4 (ranging from 86% to 93%), while concentration from 2 to 10 µg/mL exerted a cytotoxic effect (Fig. 3B). Stimulation of RAW 264.7 cells at concentrations below 1 µg/mL for 24 and 48 h appeared to be optimal for analyzing cell activation.
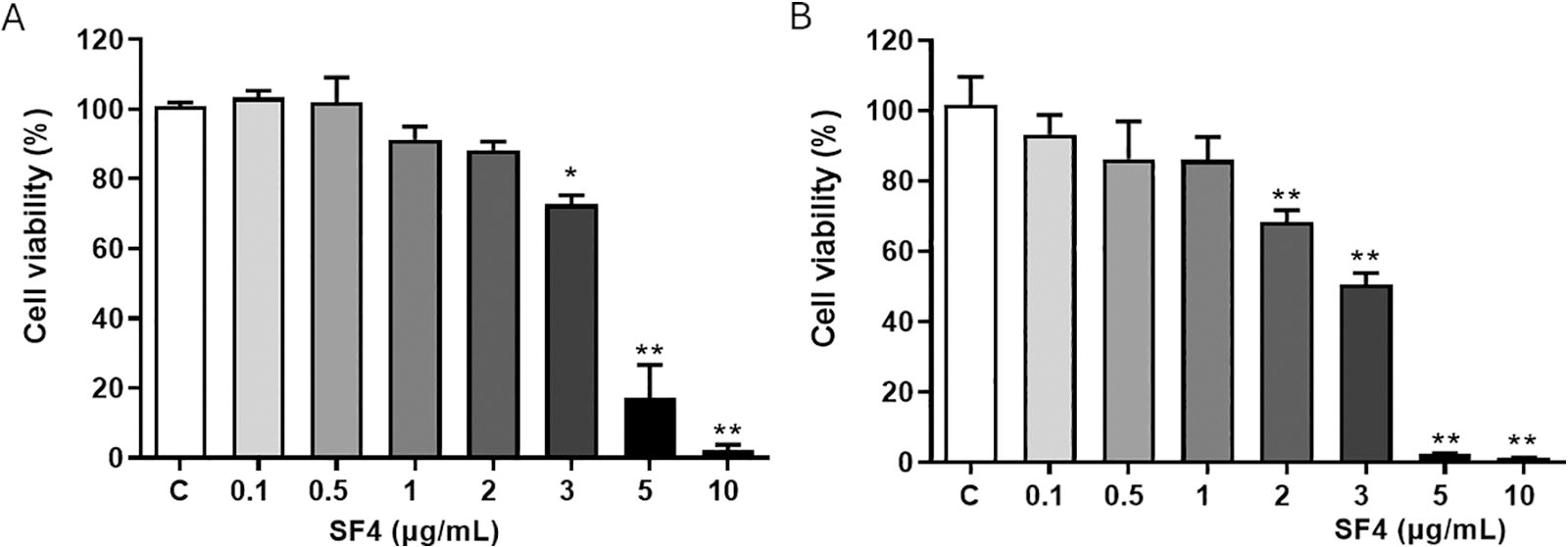
The viability of RAW 264.7 cells after the recombinant SF4 exposure at
First, we analyzed the expression of 4-1BB. After 0.1, 0.5, and 1 µg/mL SF4 stimulation, the mRNA levels of 4-1BB significantly increased at 24 and 48 h (Fig. 4A). Western blot analysis showed the expression of 4-1BB in unstimulated cells, with a significant increment in 0.5 µg/mL SF4 for 48 h (Fig. 4B). TRAF2 protein significantly increased accumulation after 24 h, although it was not significant at 48 h (Fig. 5A). Also, the p38 protein was significantly phosphorylated at 24 h with 0.1 and 0.5 µg/mL SF4. Notably, 48 h of SF4 stimulation increased the ratio of phosphorylated p38 to 0.1, 0.5, and 1 µg/mL of SF4 (Fig. 5B). The phosphorylation of IκB tended to increase after 24 h of stimulation, although it increased after 48 h at 1 µg/mL (Fig. 5C). These results suggest that SF4 activates the MAPK and NF-κB signaling pathways, which may lead to efficient costimulation during antigen presentation.

Expression of the 4-1BB in RAW 264.7. After stimuli of RAW 264.7 with SF4,

Stimulation of RAW 264.7 cells with SF4 triggers the MAPK and NF-κB pathway signaling.
To investigate whether SF4 induces the expression of type 1 cytokines by macrophages in vitro, we determined the mRNA expression levels of TNF-α, IFN-γ, and IL-12 in RAW 264.7 cells. Interestingly, 0.5 and 1 µg/mL SF4 stimulation significantly upregulated the expression of TNF-α at 24 h when compared with the control (Fig. 6A). Moreover, compared with the control, the expression of IFN-γ was significantly upregulated at 1 µg/mL at 24 h and with 0.1, 0.5, and 1 µg/mL SF4 at 48 h (Fig. 6B). Finally, IL-12 expression increased significantly only after 48 h of 1 µg/mL SF4 stimulation (Fig. 6C). Although an RSV-challenge model is needed, these in vitro results indicate that SF4 induces macrophages to express master cytokines in triggering a type 1 immune response.

Gene expression in RAW 264.7 cell after 24 and 48 h of stimuli.
We further examined the production of neutralizing antibodies in C57BL/6J mice immunized with SF4. Sera from control mice had no pre-existing neutralizing antibodies against RSV. Titers of neutralizing antibodies increased in the sera from immunized animals (Fig. 7A). RSV replication was significantly inhibited by 1.16, 0.99, and 1.97 logs when virions were preincubated with serum from SF4-immunized animals, compared with nondiluted sera from control animals (Fig. 7B). However, further studies are needed to evaluate its efficacy against challenge using an in vivo model of infection.

Neutralizing antibodies and inhibition of viral replication.
The concentrations of type 1 (TNF-α, IFN-γ, and IL-2) and 2 (IL-4 and IL-5) cytokines in the serum were determined in immunized mice with 20 µg SF4, since these animals presented a higher neutralizing antibody titer. We found that the concentrations of TNF-α, IFN-γ, IL-2, and IL-4 in the immunized groups were significantly higher than those in the control group (Fig. 8), with IL-2 being the most abundant cytokine. Finally, there was no detection of IL-5 in any of the study groups, and it is worth mentioning that the kit controls validated the test. Even in the presence of increased levels of IL-4, the type 1 response predominated, as there were no detectable levels of IL-5. Overall, these data indicate that the immune response generated by SF4 is mainly biased toward a type 1 response, which prevents adverse type 2 inflammation.

Analysis of the secretion of Th1 and Th2-related cytokines in serum of SF4-immunized mice. Th1 cytokines levels, TNF-α, IFN-γ, and IL-2, were increased after SF4 immunization, together with IL-4. Data are presented as mean ± standard error of the mean. *p < 0.05 versus control.
Discussion
Currently, RSV is one of the most important etiological agents of acute lower respiratory infections and is considered a serious burden to public health worldwide. Various studies have focused on searching for viral antigenic molecules. Among the candidates are the F and G proteins (Borchers et al., 2013; Boyoglu-Barnum et al., 2019). However, the G protein induces an exacerbated Th2 response and pulmonary eosinophilia (Johnson and Graham, 2004), which makes it unsuitable for a vaccine. Thus, in 2023, the U.S. Food and Drug Administration approved a vaccine against RSV based on pre-F for the geriatric population (Kingwell, 2023). In an animal study, vaccination of pregnant lambs with the F protein linked to an adjuvant (F/TriAdj) raised RSV F-specific immunoglobulin (Ig)G, which was passively transferred to the offspring, preventing lung pathology after RSV infection (Garg et al., 2015). Thus, F protein may stimulate protective immunity.
In some patients, RSV triggers a type 2 response that facilitates proasthmatic inflammation and viral persistence, while, in others, type 1 immune response can eliminate the virus (Gut et al., 2013). In this study, we evaluated the ability of SF4 to modulate a type 1 immune response and the generation of neutralizing antibodies against RSV. The SF4 protein was designed to contain the extracellular portion of 4-1BBL, as several studies demonstrated that costimulatory molecules and antigens may drive the immune response (Engelstein et al., 2016; Powell et al., 2015).
4-1BB is broadly expressed in diverse immune cells, although it is primarily expressed in activated cytotoxic CD8+ T cells and helper CD4+ T cells, in which it enhances T-cell response to antigens and expansion (Nielsen et al., 2020; Singh et al., 2024). Activation of the 4-1BB receptor leads to the association of the adapter proteins TRAF1 and 2, leading to the activation of NF-κB and p38 and production of TNF-α (Cannons et al., 2000; Kang et al., 2007; Saoulli et al., 1998; Youlin et al., 2013). Our results show that the SF4 protein upregulates TRAF2 expression and activates NF-κB and p38 in macrophages, which are associated with TNF-α, IFN-γ, and IL-12 gene expression. In addition, blocking 4-1BB/4-1BBL suppresses the activity of p38 and IκB, resulting in a decrease in TNF-α (Haga et al., 2009). Moreover, immunization of mice with 4-1BBL-expressing RM-1 cells induced secretion of IFN-γ, TNF-α, and IL-2, leading to cytotoxicity in prostate cancer (Youlin et al., 2012). Also, the increment of TNF-α, IFN-γ, and IL-2 influences the induction of IgG in mice immunized with an HBsAg vaccine (Rahimkhani et al., 2021). Moreover, the production of neutralizing antibodies IgG2a against HIV is accompanied by an increase in TNF-α, IFN-γ, and IL-2-secreting CD8 T-specific cells (Huang et al., 2017). Thus, it is likely that type 1 cytokines produced by SF4-activated macrophages promote the production of neutralizing antibodies against RSV. In contrast, although levels of IL-4 appeared to increase, it is likely that those of IL-2 and IFN-γ surpass the effect of IL-4 in a mechanism to maintain homeostasis and avoid saturation of the immune response (Zhang et al., 2024). One of the main concerns in the development of vaccines for the prevention of RSV is the ability to generate type 2 responses that favor adverse reactions. Therefore, the evaluation of type 2 cytokines after immunization is a critical point. Immunization with the F protein precipitated with aluminum hydroxide promotes the increase of IL-13 levels; however, the use of monophosphoryl lipid A and oligodeoxynucleotide CpG prevents its increase and consequently induces a decrease in inflammatory RSV disease after infection (Kawahara et al., 2023; Lee et al., 2019). It has been observed that recombinant adenoviruses that express the F protein have a low capacity to induce cytokines such as IL-4, IL-5, and IL-13 (Shao et al., 2009). In the present study, the evaluation of IL-13 could provide information on a possible type 2 response and is the aim of our current work.
In this study, stimulation with the SF4 protein induced overexpression of 4-1BB, TNF-α, IFN-γ, and IL-12 mRNA through the NF-κB and MAPK pathways, as an increase in the phosphorylation of IκB and p38 was observed. The upregulated expression of IFN-γ could be linked to an increase in TRAF2, enhancing the response to 4-1BB/4-1BBL activation. In INS-1E cells, stimulation with IFN-γ increases TRAF2 expression, which is associated with phosphorylation of p38 (Prause et al., 2016). It is known that 4-1BB signaling promotes activation of other MAPK, such as ERK and AKT, which leads to inactivation of glycogen synthase kinase-3 and β-catenin increased levels (Lee et al., 2013). Evaluation of the activation state of other MAPK pathways should provide more precise information about the mechanisms induced by SF4.
Monoclonal antibodies, such as Palivizumab, which bind to antigenic site II in the F protein, are highly effective in the prevention and treatment of RSV infection in high-risk patients (Pisuttinusart et al., 2024; Esposito et al., 2022). An effective humoral response to neutralizing antibodies can be achieved by increasing the expression of surface receptors, cytokines, and chemokines (Linnik and Egli, 2016). In the viral neutralization assay, we found that sera from immunized mice neutralized the RSV infection. When the animals received two doses of 20 µg, titers of up to 1/16 of neutralizing antibodies were obtained, which were associated with increased serum levels of TNF-α, IFN-γ, and IL-2.
Further studies are required to enhance the immunogenicity of SF4, determine its efficacy to prevent viral infection, and evaluate inflammation markers commonly associated with RSV infection and pathophysiological changes after RSV challenge.
Footnotes
Acknowledgments
H.-M.A. and B.-G.C.B. received a master fellowship from CONAHCYT (#803202 and #756640, respectively).
Authors’ Contributions
H.-M.A., B.-G.C.B., R.-A.J., C.-D.L.E., J.M., and F.-R.J.C.: methodology. C.-D.J.E. and M.d.O.-L.R.: resources and supervision. H.-M.A., B.-G.C.B., C.-D.L.E., E.S., and C.-G.D.: formal analysis, and writing—review and editing. E.S. and C.-G.D.: conceptualization. All authors reviewed and approved the final article.
Author Disclosure Statement
The authors have no competing interest.
Funding Information
This work was supported by the National Council of Humanities, Sciences, and Technologies (CONAHCYT, APN-5901 to C.-G.D.) and the Autonomous University of Aguascalientes (PIBB 19–9N and PIBB 23–4 to C.-G.D.).