
Abstract
Respiratory syncytial virus (RSV) is a significant cause of disease in the young and old. Recently, pre-fusion F protein vaccines for RSV have received food and drug administration (FDA) approval to protect adults aged 60 years and older; however, vaccines evaluated against RSV typically do not elicit complete or durable protective immunity. We previously showed that an RSV G protein central conserved domain (CCD) nanoparticle vaccine containing an S177Q mutation (NP-S177Q) induced favorable immunogenicity and RSV-neutralizing antibodies compared with RSV G protein vaccination alone in mice. Boosting BALB/c mice with NP-S177Q vaccines improved correlates of protection and reduced markers of immunopathology following RSV challenge. This study examined microparticle (MP) vaccines displaying the CCD with an RSV G S177Q mutation (MP-S177Q) adjuvanted with monophosphoryl lipid A (MPLA) in BALB/c mice. Our findings show that mice adjuvanted MP-S177Q vaccination develop effective viral neutralization compared with MP-WT and MP-S177Q vaccination and have improved bronchoalveolar Th1-type cytokine responses following the RSV challenge compared with MP-WT or vehicle-vaccinated mice. This study shows that a rationally mutated RSV G protein MP vaccine is safe, effective, and can advance precision RSV vaccines.
Introduction
Precision vaccination, or the ability to optimize vaccines specific to vulnerable populations, e.g., the young, elderly, or immunocompromised, has become reachable based on customized platforms that provide new vaccines and immunization strategies for different populations. For example, the recent approval of respiratory syncytial virus (RSV) vaccines is based on vaccination recommendations for specific risk groups to provide the safest and most effective use of vaccines (Ruckwardt, 2023). The FDA approval of Abrysvo™ was the first RSV vaccine approved for use in pregnant individuals to prevent lower respiratory tract disease (LRTD) caused by RSV in infants from birth through 6 months of age, and Abrysvo was approved for use at 32 through 36 weeks gestational age of pregnancy. Later, the FDA approved Abrysvo for preventing LRTD caused by RSV in individuals 60 and older (Kampmann et al., 2023). Further, Beyfortus™ (nirsevimab), an extended half-life prophylactic monoclonal antibody (mAb) targeting the pre-fusion F protein, was approved for the prevention of RSV in neonates and infants born during or entering their first RSV season and in children up to 24 months of age (Jorgensen, 2023; Zhaori, 2023).
Developing safe and effective RSV vaccines is essential because RSV is a leading cause of respiratory disease in infants, young children, the elderly, and the immunocompromised (Chatzis et al., 2018; Jain et al., 2024). Childhood RSV disease has a substantial global health and economic burden, and RSV is the primary cause of hospitalization due to LRTD among infants worldwide (Hall, 2012). Another advancement in RSV vaccine development has been the layer-by-layer (LbL) method of vaccine fabrication, which offers simplicity in the assembly of vaccine platforms and allows for the rapid growth of synthetic vaccine components, providing the foundation for optimizing vaccine antigens needed for precision vaccine development (Guzman et al., 2023). Additionally, LbL vaccines offer versatility and modularity, opening new paths for designing vaccination platforms with high efficiency and specific targeting and minimizing the economic impact associated with their fabrication. Microparticle (MP) vaccines utilizing LbL construction have been used to develop the RSV vaccines (Jorquera and Tripp, 2016). LbL vaccines facilitate how antigen payloads are presented to the immune system by antigen-presenting cells (APCs), they enhance antigen uptake by APCs, act as an antigen depot, and modulate the Th1/Th2 immune response when engineered to include an immune agonist (Jorquera and Tripp, 2016). We previously showed that immunizing mice with LbL nanoparticle vaccines containing an RSV G peptide with a CX3C chemokine motif (aa 169–198) and a CD8+ epitope from RSV M2 protein-induced antibodies that block CX3CL1 (fractalkine) chemotactic activity of RSV G protein and protected from infection replication post-RSV challenge (Jorquera et al., 2013; Jorquera et al., 2015).
The RSV G protein is 298aa, depending on the RSV strain or cell line used to propagate RSV, which is a viral surface attachment protein that contains cytosolic, transmembrane, and ectodomain regions (McLellan et al., 2013). The intact G protein is heavily glycosylated and diverse. However, the central conserved domain (CCD; aa 157–198) is highly conserved between RSV A and B, lacks glycosylation, and contains a CX3C chemokine mimic motif (aa182–186) that binds to the fractalkine receptor, CX3CR1, and can trigger fractalkine-like host cell responses that contribute to inflammatory mechanisms (Anderson et al., 2021; Bakre et al., 2017; Chirkova et al., 2013). The G protein CX3C motif interacts with CX3CR1 on ciliated respiratory epithelia and some immune cells (Chirkova et al., 2015; Jeong et al., 2015; Johnson et al., 2015; Zhang et al., 2002). Interestingly, the RSV G protein has an alternative translation site (Met 48), which results in an unstable transmembrane domain and the release of a soluble form of G protein (sG) protein (Johnson et al., 2015). sG contains the CX3C motif and, like membrane-bound G (mG), functions to interfere with host immunity (Liang et al., 2019). Previous work by our group and others has shown that the CX3C motif of RSV G protein impacts immunity by biasing toward a Th2-type cytokine response, reducing interferon gamma (IFNγ) and interleukin-6 (IL-6), altering trafficking of CX3CR1+ cytotoxic T lymphocyte and natural killer cells, inducing pulmonary eosinophilia, modifying host miRNAs, dampening type I IFN responses, impacting toll-like receptor 4 (TLR4) signaling, and reducing antibody production (Bergeron et al., 2023a; Bergeron and Tripp, 2021; Bergeron and Tripp, 2022). Notably, several studies have shown antibodies against RSV G protein are protective, interfere with RSV-mediated immune antagonism, improve protective Th1-type immunity, and block CX3C-CX3CR1 interactions (Bergeron et al., 2023b; Bergeron et al., 2023c; Boyoglu-Barnum et al., 2013; Boyoglu-Barnum et al., 2015; Caidi et al., 2018; Haynes et al., 2009; Lee et al., 2024; Tripp et al., 2018).
The RSV G protein is attractive for vaccine development because the G protein CCD is essential for infectivity in vivo, it mediates viral attachment to airway epithelial cells, and the CCD has a conserved CX3C chemokine motif implicated in the alteration of the host immune response (Tripp et al., 2018). It has been shown that mAb TRL3D3, which binds with low picomolar affinity to an epitope within the CCD, can neutralize soluble G (sG) and has antiviral activity (Fedechkin et al., 2020; Fedechkin et al., 2018; Haynes et al., 2009; Hijano et al., 2019; Tripp et al., 2018). Thus, safety, the induction of a balanced Th1/Th2 cytokine response, and reduced G protein reactogenicity are essential considerations in RSV vaccine development. Recent structural studies revealed conformational epitopes in a larger region of RSV G, i.e., between aa157–198 (Fedechkin et al., 2020; Fedechkin et al., 2018; Nunez Castrejon et al., 2022). Previously, we showed that RSV G with an S177Q mutation does not alter conformational epitopes but increases anti-G protein antibody responses and Th1-type cytokine responses, improves RSV neutralization, and does not mediate enhanced disease compared with the wild-type G protein (Bergeron et al., 2023d; Bergeron et al., 2021; Nunez Castrejon et al., 2022).
In this study, we compared rationally designed G protein mutants with wild-type G protein immune responses in both male and female BALB/c mice as it is known there are sex-dependent immune responses to vaccinations and infections (Ursin and Klein, 2021), and elucidating these similarities and differences improves preclinical modeling. These results showed that mice boosted with MP-S177Q developed better immunogenicity and neutralizing antibodies than MP-WT boosting. Importantly, MP-S177Q vaccination led to potent viral neutralization compared with MP-WT, and MP-S177Q vaccination improved bronchoalveolar lavage fluid (BALF) Th1-type cytokine concentrations following the RSV challenge compared with MP-WT and vehicle-vaccinated mice. This study shows that a rationally mutated RSV G protein MP vaccine is safe, effective, and can advance precision RSV vaccines.
Materials and Methods
Cells and viruses
Vero E6 cells (CRL-1586), A549 cells (CCL-185), and HEp-2 cells (CCL-23) all from American Type Culture Collection (ATCC, Manassas, VA), were maintained in 10% fetal bovine serum (FBS) in Dulbecco's Modified Eagle Medium (DMEM) (Hyclone, Logan, UT). RSV A2 (ATCC VR-1540) and RSV A2 expressing GFP (a kind gift from Dr. Marty Moore, Meissa Vaccines) were propagated in HEp-2 cells (ATCC CCL-23) as described (Murray et al., 2022).
MP construction
LbL-MP was fabricated as previously described (Powell et al., 2022; Powell et al., 2011; Powell et al., 2013). Poly-
Microparticle Vaccine Peptides
ELISA analysis of MPs with mAb 2D10
All antigens (MP-WT, MP-S177Q, MP-empty, and recombinant RSV G ectodomain) were examined using high-binding enzyme-linked immunosorbent assay (ELISA) plates (Costar, Corning, NY) at 10 µg/mL in phosphate-buffered saline (PBS). Recombinant RSV G ectodomain was generated as described previously (Nunez Castrejon et al., 2022). PBS was plated for negative control wells. Plates were left at 4°C overnight. Plates were washed four times with PBS-T (PBS + 0.1% Tween). Blocking buffer (PBS-T + 5% milk) was added to all wells and incubated for 2 h at room temperature. mAb 2D10 (Trellis Biosciences) was diluted to 5 µg/mL in blocking buffer and serially diluted 1:3 with blocking buffer. The blocking solution was decanted, and 150 µL of serially diluted mAb 2D10 was added to wells. Plates were incubated for 1 h at room temperature and then washed four times with PBS-T. In total, 50 µL of goat anti-human IgG Fc secondary antibody conjugated to horseradish peroxidase (HRP) (Invitrogen A18817, Carlsbad, CA) diluted 1:3000 in PBS-T + 1% milk was added to all wells. Plates were incubated for 1 h at room temperature and then washed four times with PBS-T. Each well was added 100 µL of tetramethylbenzidine (TMB) substrate (Sigma T0440, Burlington, MA). Plates were developed for 10 min, at which point the reaction was quenched by adding 100 µL 1N sulfuric acid. Absorbance at 450 nm was quantified using a plate reader. Plotting with standard error of the mean (SEM) values was performed using MS Excel.
Mice
Male and female BALB/c mice (10–12 weeks old; Jackson Laboratories, Bar Harbor, ME) were housed in micro isolator cages with 12 h light/dark cycle and fed ad libitum. The mice received a priming dose of 10 µg MP-WT, MP-S177Q, or empty MPs. All vaccines were adjuvanted with 10 µg monophosphoryl lipid A (MPLA; VacciGrade™ from S. Minnesota R595, InvivoGen, San Diego, CA; a TLR4 agonist) diluted in PBS. Mice were i.m. vaccinated in the left and right rear quadriceps with 0.05 mL/quadriceps. On day 21 post-prime, mice were boosted with the same dose of the homologous vaccine. Mice were bled on day 28 post-prime (day 7 post-boost). On day 35 post-prime (day 14 post-boost), mice were i.p. anesthetized with 2, 2, 2-tribromoethanol (Sigma Aldrich, St. Louis, MO), and i.n. challenged with 106 PFU RSV A2 diluted in PBS. Mice were monitored daily and euthanized on day 5 pi. Sera, BALF, and lungs were collected and stored in serum-free-DMEM on ice during organ processing.
ELISA
Sera were evaluated for anti-RSV IgG levels. Briefly, high-binding ELISA plates (Corning, Corning, NY) were coated with RSV A2 or B1 (5 µg/mL) overnight at 4°C. The next day, the wells were washed three times with 1× KPL wash buffer in distilled water (SeraCare, Milford, MA) and blocked with Blotto (5% non-fat dry milk) for 1 h at 37°C. Blotto was removed, and the sera were 1:3 diluted (starting at 1:50) in Blotto and added to wells for 1 h at 37°C. The wells were washed 3× with KPL wash buffer, and secondary goat-anti-mouse IgG-AP (ThermoFisher, Waltham, MA) or secondary subtype IgG1 or IgG2a antibodies (Southern Biotech, Birmingham, AL) were added. Plates were incubated for 1 h at 37°C, washed 3× with KPL wash buffer, and developed with p-nitrophenyl phosphate (pNPP; ThermoFisher) for 45 min, and read using a BioTek plate reader (Bio-Tek, Winooski, VT) at OD405. The area under the curve (AUC) was calculated using Prism 10 (GraphPad, La Jolla, CA).
Virus neutralization
To determine the level of antisera neutralization, a reporter-based microneutralization protocol was performed as previously described (Bergeron et al., 2023d). Briefly, sera were pooled and heat-inactivated at 56°C for 30 min. Sera were two-fold diluted (starting at 1:40) and pre-incubated with RSV A2/GFP (MOI = 0.1) for 1 h at 37°C ± 10% guinea pig complement (C’) (NovusBio, Centennial, CO). After pre-incubation, the mixture was added to confluent A549 cells in a 96-well plate for 24 h. At 24 hpi, cells were gently washed, fixed with 4% paraformaldehyde (PFA; Ted Pella, Redding, CA) for 20 min at room temperature, and counter-stained with 1 µg/mL 4’,6-diamidino-2-phenylindole (ThermoFisher). Plates were read using Cellomics ArrayScan (ThermoFisher), and fluorescent focus units (FFUs) were automatically enumerated. The AUC was calculated using Prism 10 (GraphPad, La Jolla, CA).
RSV plaque assays
Lungs were harvested on day 5 pi and homogenized in 1 mL DMEM using a GentleMACS tissue homogenizer (Miltenyi Biotec, Gaithersburg, MD). Homogenates were centrifuged at 500 ×g at 4°C for 8 min, the supernatant diluted 10-fold in DMEM (Hyclone), and overlaid onto 90% confluent Vero E6 cells in 24-well plates. After 2 h of absorption, cells were overlaid with 2% methylcellulose (Sigma Aldrich) and incubated at 37°C for 7 days. Following incubation, methylcellulose was aspirated, the cells washed with PBS, fixed with acetone:methanol (60:40, Sigma Aldrich), and air-dried overnight. Wells were washed 3x with KPL wash buffer (VWR, Radnor, PA) and blocked with Blotto (ThermoFisher) overnight at 4°C. The next day, Blotto was removed, and a mAb cocktail against RSV F and G proteins (clones 131-2A, 131-2G, Tripp Laboratory, UGA) was diluted in Blotto (ThermoFisher) and added overnight at 4°C. Wells were washed 3× with KPL wash buffer (VWR), and goat anti-mouse-AP (ThermoFisher) was added overnight at 4°C. Wells were washed 3× with KPL wash buffer (VWR), and RSV plaques were developed with KPL TrueBlue substrate solution (VWR) for 5 min, rinsed with distilled water, and enumerated using a dissection microscope (VWR).
BALF analysis
BALF was collected from terminally bled mice. The trachea was exposed, and the lungs were flushed 3× with 1 mL 0.5% BSA/PBS collected and centrifuged for 10 min at 500 ×g at 4°C to isolate cell-free fluid. The BALF was stored at −80°C until analysis. BALF was analyzed in the Milliplex MAP Mouse Cytokine/Chemokine Immunology Multiplex Assay as described by the manufacturer (Millipore Sigma, Rockville, MD) using standards and quality controls included in the kit. Individual BALF samples were duplicated from n = 3–5 mice/group/sex. Samples were analyzed on a Luminex 200 instrument (Luminex Corporation, Austin, TX) using Luminex xPONENT 3.1 software.
Statistics
Data were analyzed by one-way analysis of variance with Dunnett’s multiple comparison test, or Kruskal-Wallis was performed for non-parametric tests. p < 0.05 was considered significant. Data are presented as mean ± SEM. The vaccination study was performed once for male and female mice, while experiments were performed at least in duplicate with representative data shown.
Results
S177Q vaccination improves immunogenicity and neutralization
Previous results have shown that while the antibody response to RSV G protein is protective, vaccination with G protein containing an unmodified CX3C motif may not be optimal for inducing the greatest immunogenicity, a feature linked to the CX3C motif (Chirkova et al., 2013; Harcourt et al., 2004). We previously identified the serine-to-glutamine point mutation at aa177 (i.e., S177Q), which improves humoral responses to vaccination in intact G protein and G nanoparticle formulations (Bergeron et al., 2023d; Bergeron et al., 2021). We first evaluated MPs for proper folding and display of CCDs using an ELISA with mAb 2D10, which recognizes a conformational epitope that requires native disulfide linkages in the CCD (Fig. 1A). We found that mAb 2D10 bound to MP + wildtype G CCD (WT) and MP + G CCD S177Q (S177Q) but not empty MP (Fig. 1B). These data support that MPs display correctly folded and disulfide-bonded CCDs and that the S177Q mutation does not disrupt this folding.
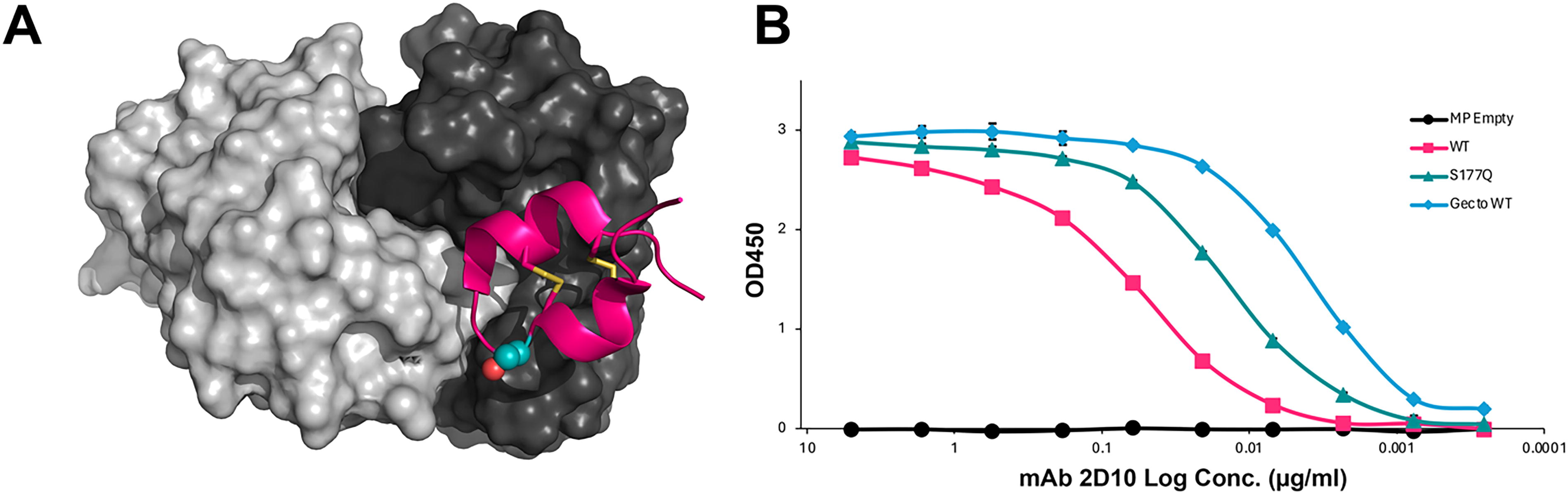
mAb 2D10 binds its conformational epitope on WT and S177Q MP constructs.
To determine if the mutation improved immunogenicity in a LbL-MP vaccine platform, male or female BALB/c mice were immunized on days 0 and 21 with 10 µg of either empty MP, MP + wildtype G CCD (WT) or MP + G CCD S177Q (S177Q), all adjuvanted with 10 µg MPLA. Sera from vaccines were examined for antibody responses on day 28 post-prime (day 7 post-boost). WT and S177Q vaccination induced vigorous antibody responses to RSV compared with empty MP or naïve controls (Fig. 2). Importantly, S177Q vaccination resulted in increased antibody responses compared with WT in female (p = 0.12) (Fig. 2A and C) and male (p = 0.074) mice (Fig. 2B and D). These findings are consistent with our previous study (Bergeron et al., 2023d). Although the magnitude of the serum antibody response was lower compared with responses to RSV A2 vaccination, S177Q antisera also reacted to RSV B1 by ELISA (Supplementary Fig. S1) from male (p = 0.0574) and female (p = 0.1954) vaccinees, while WT sera did not (p = 0.42 for males and p = 0.99 for females) compared with naïve control sera.
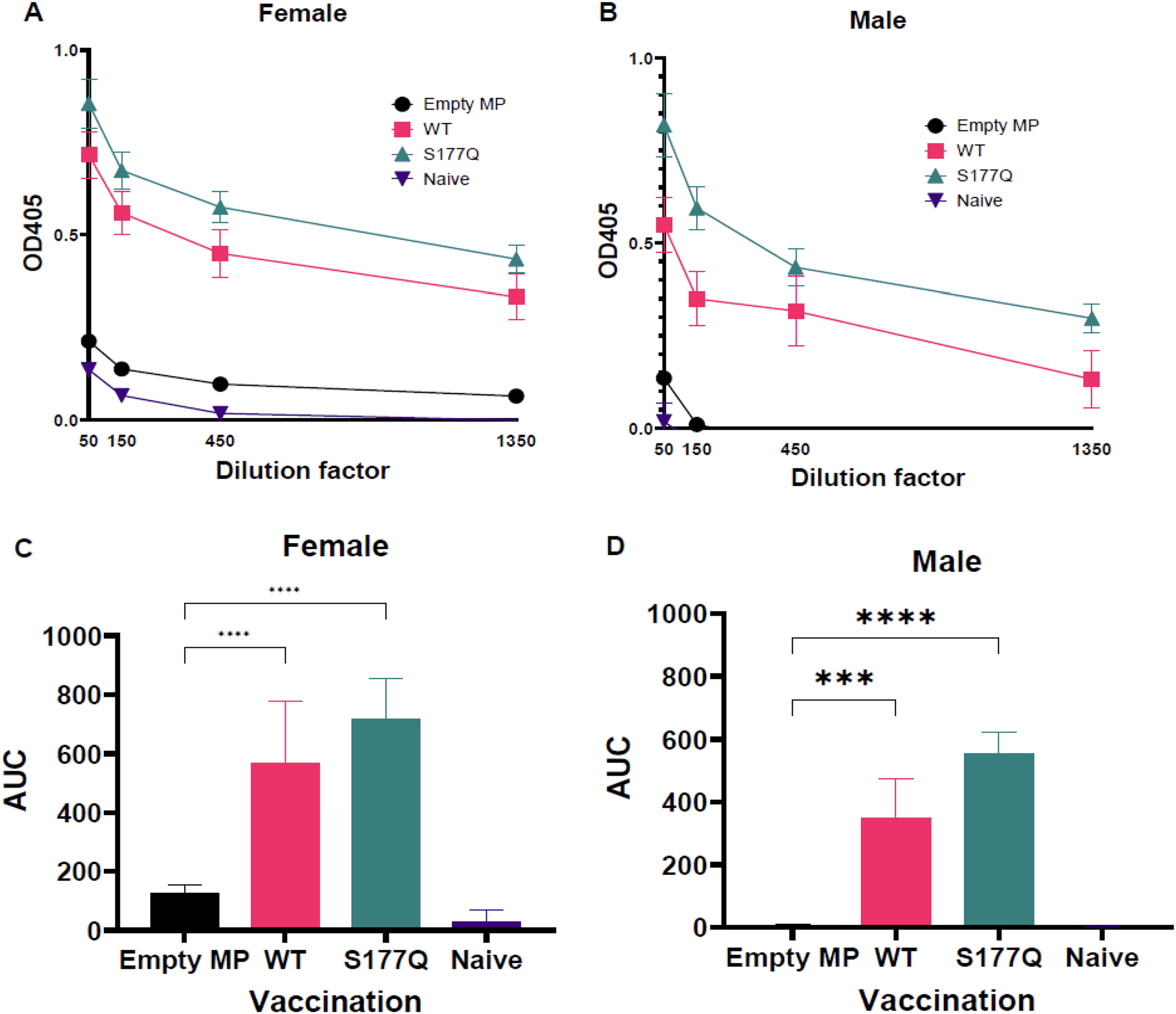
MP constructs are immunogenic, and S177Q improves immunogenicity. Female
Next, we determined the antisera-neutralizing ability generated by MP vaccination (Fig. 3). Sera from each group were heat-inactivated, pooled, and pre-incubated with RSV A2-GFP (MOI = 0.1) for 1 h in the presence or absence of 10% guinea pig C’. Sera/virus mixture was then added to A549 cells for 24 h to determine neutralization. Strikingly, S177Q vaccination induced neutralizing antibodies regardless of C’ addition in both female (Fig. 3A and C) and male (Fig. 3B and D) mice. WT G induced neutralizing antibodies in female mice in the absence or presence of C’ (p = 0.054, p = 0.062, respectively), although to a lower extent than S177Q. While empty MP vaccination did not induce substantial neutralizing antibody responses, it did elicit low levels of virus neutralization compared with naïve sera in the presence of complement. This result may be due to the induction of non-specific antibody responses that functioned with C’ or other effector mechanisms, resulting in low levels of a non-specific neutralization activity. These data show that S177Q vaccination induces a more significant neutralizing response than WT vaccination.

S177Q induces neutralizing antibodies. Female
The anti-RSV antibody response after RSV challenge
As expected, the WT and S177Q vaccines were immunogenic after priming and boosting (Fig. 2). We sought to evaluate the antibody response after RSV infection. WT and S177Q vaccinated mice were i.n. challenged with 106 PFU RSV A2, and on day 5 pi sera were collected and analyzed (Fig. 4). Male and female mice vaccinated with WT or S177Q vaccines had noteworthy increases in serum IgG compared with empty MP vehicle control or WT vaccines (Fig. 4A–D) (p < 0.05), but no significant differences between WT and S177Q vaccination. One metric for a safe RSV G vaccine is avoiding a Th2-type biased immune response following RSV infection, correlated with enhanced pulmonary RSV disease (Bergeron and Tripp, 2021). Antisera were analyzed for IgG subtypes to determine the Th1-type (IgG1) and Th2-type (IgG2a) antibody responses in vaccinated and challenged mice. IgG1 levels were significantly (p < 0.05) increased in both WT and S177Q vaccinations compared with empty MP with no significant differences between WT and S177Q (Fig. 4E and F). While there were significant (p < 0.05) but modest increases in female IgG2a responses in S177Q responses compared with controls, there were no considerable antibody increases compared with WT vaccinated mice (Fig. 4G). Interestingly, IgG2a levels were significantly (p < 0.05) increased in S177Q compared with all other groups of vaccinated male mice (Fig. 4H). These data indicate that S177Q vaccination can mediate a balanced recall Th1/2-type antibody response.

MP vaccination induces an antibody recall response after an RSV challenge. Female
Vaccination reduces the lung viral load
To determine the ability of vaccination to protect against RSV challenge, immunized mice were challenged with 106 PFU RSV A2 on day 5 pi, the lungs were collected, and viral titers were determined by plaque assay. Significant (p < 0.05) reductions in lung viral titers were observed in female (Fig. 5A) and male (Fig. 5B) mice vaccinated with WT or S177Q. Interestingly, vaccinated male mice had lower lung virus titers than female mice, and WT and S177Q vaccination was associated with reduced lung viral titers in both sexes. WT and S177Q vaccination in female mice reduced viral titers >2 logs compared with controls. Male mice vaccinated with WT had approximately a 2-log reduction of lung titers compared with controls. However, S117Q vaccinated mice had >4 log reduction, and no detectable infectious virus was recovered.
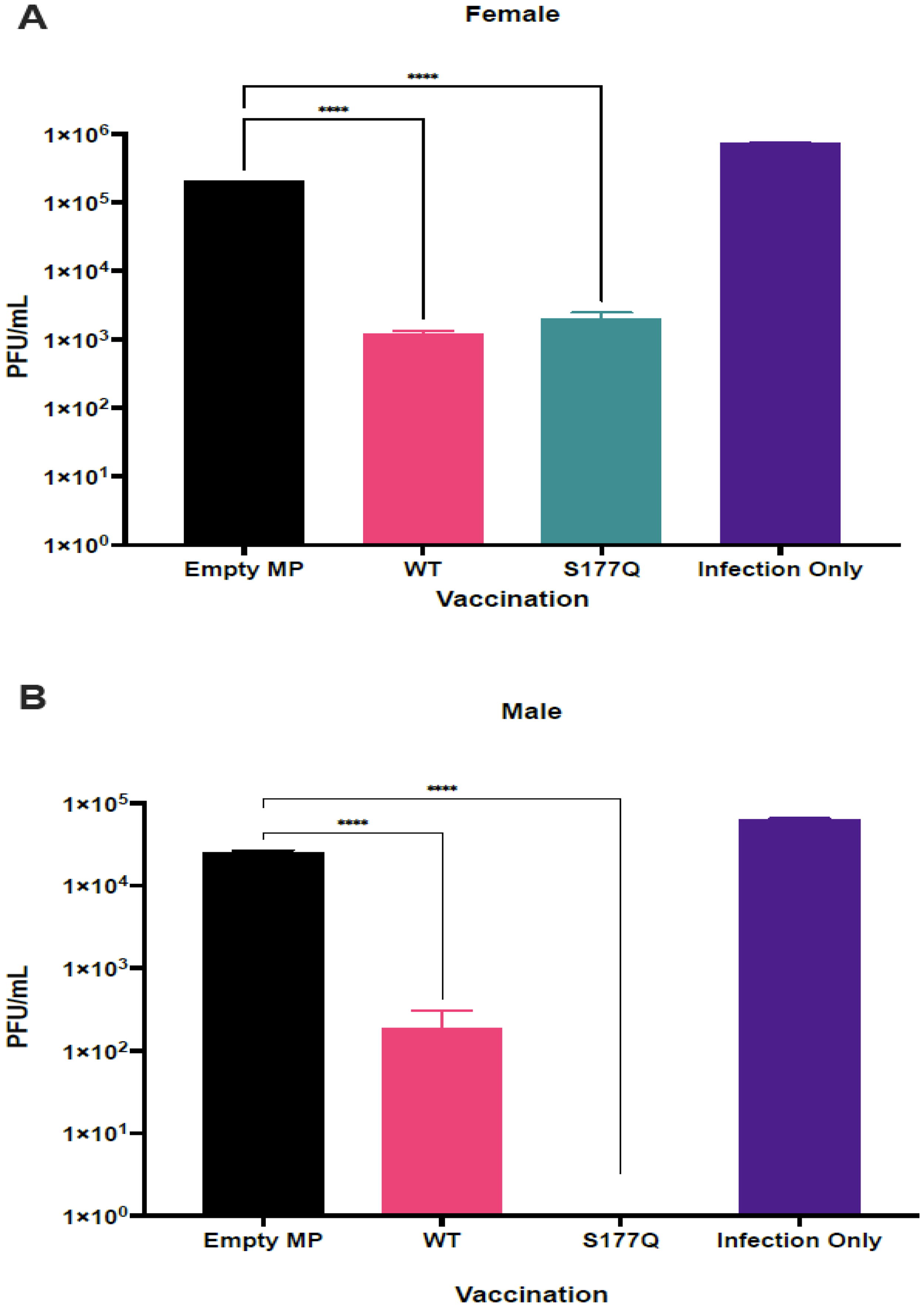
MP vaccination reduces lung titers. Female
Vaccination modifies the cytokine and chemokine responses to RSV challenge
An indicator of RSV disease is distorted cytokine or chemokine responses. To evaluate this, on day 5 pi, the BALF was collected from vaccinated and challenged mice. The concentrations of cytokines and chemokines were measured by bead-based multiplex cytokine assay. Multiple cytokines and chemokines were evaluated in BALF from vaccinated groups using bead-based multiplex assays (Supplementary Fig. S2). The results showed variation between sexes and some vaccination groups. However, for two cytokines, IFNγ, and IP-10, there were significant increases in the BALF from female and male mice vaccinated with S177Q (Fig. 6). IFNγ is a canonical Th1-type cytokine, and IP-10 is a chemokine affected downstream of IFNγ. Both are biomarkers for a Th1-type immune response. While the magnitude differed somewhat, female mice had significant (p < 0.05) increased IFNγ compared with control groups and were increased compared with WT BALF (p = 0.056) (Fig. 6A). Male mice had significant (p < 0.05) increases in BALF IFNγ levels compared with all other groups (Fig. 6B). IP-10 concentrations in BALF of female mice were increased in S177Q compared with empty MP and infection-only controls (Fig. 6C). In contrast, male BALF IP-10 was significantly (p < 0.05) increased compared with WT and the infection-only control (Fig. 6D). These data back improved Th1-type responses in S177Q vaccinated mice following infection. IgG subtype data (Fig. 4) suggest these vaccines induce a Th1/2 balanced response that varies subtly between males and females.
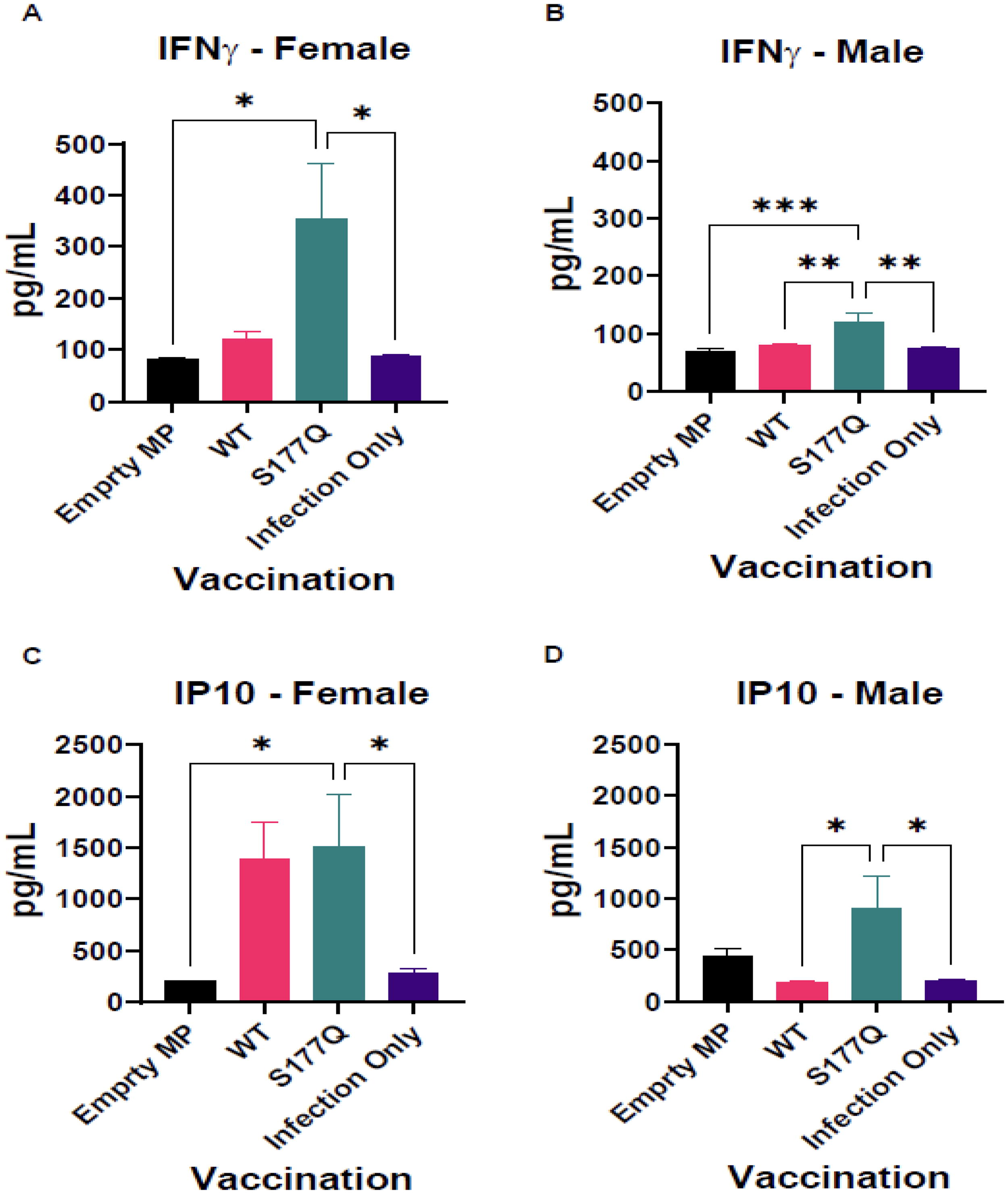
Cytokine/chemokine responses following RSV infection. Female
Discussion
When considering RSV vaccine platforms, vaccine safety is crucial because it has been shown that some viral vaccines may induce vaccine-enhanced respiratory disease (VERD), which is atypical but can occur when vaccination promotes aberrant immune responses that exacerbate the disease caused by subsequent infection with the associated pathogen. VERD has been observed in humans in three vaccine trials against RSV, dengue, and measles (Fulginiti et al., 1967; Kim et al., 1969; Sridhar et al., 2018). In 1966, RSV vaccine studies in infants and young children using a formalin-inactivated vaccine against RSV (FI-RSV) found that immunized children who were subsequently exposed to environmental RSV experienced an enhanced disease, and two immunized infants died (Kim et al., 1969). It was subsequently shown that the FI-RSV vaccine stimulated an unbalanced immune response. The induced antibodies were directed against nonprotective post-F conformation epitopes concomitant with substantial Th2-type immune responses (Acosta et al., 2015; Connors et al., 1994; Murphy and Walsh, 1988; Waris et al., 1996). Of particular note, some formulations of RSV G vaccines may prime for enhanced, Th2-type disease. This priming for vaccine-enhanced disease must be considered when developing RSV G protein-based vaccines (Acosta et al., 2015; Bembridge et al., 1998; Bembridge et al., 1999; Haynes et al., 2003; Johnson and Graham, 2004).
Significant advancements in RSV countermeasures have recently occurred. Currently, two RSV pre-fusion F vaccines are available, one for older adults and one for pregnant women, and a new immune prophylactic is available for all infants (Kampmann et al., 2023; Keam, 2023). Despite these successes, there remains a need for safe and effective RSV vaccines for young children, especially those 2–5 years of age, for whom none of the new countermeasures are approved. We have designed modalities targeting the RSV G protein to address these gaps and avoid potential reactogenicity. During infection, the G protein is responsible for initial virion attachment to CX3CR1 expressed on airway epithelial cells and interfering with host immunity to infection (Bergeron and Tripp, 2021; Bergeron and Tripp, 2022). Therefore, targeting the CX3C motif within the RSV G protein provides two biological mechanisms of protection—reducing initial virion attachment to host cells and preventing immune modulations mediated by G protein. We and others have previously demonstrated that RSV G protein countermeasures are safe and effective. Specifically, we communicated a G protein CCD NP vaccine that induced RSV neutralization in primary human airway epithelial cell cultures (Voorzaat et al., 2024), and our previous RSV G NP vaccines elicited antibody responses that were neutralizing in vivo and in vitro with the addition of complement (Bergeron et al., 2023d). This study demonstrates the promise of RSV G CCD MP vaccines. Notably, similar to the now-approved RSV pre-fusion F vaccines, these RSV G vaccines induce strong antibody responses that neutralize in vitro and reduce viral load in vivo (McLellan et al., 2013).
Another hurdle in developing RSV G protein vaccines is relatively poor immunogenicity. The G protein may prevent sufficient antibody responses, potentially due to infection and re-regulation of neonatal regulatory B cells (nBreg) (Zhivaki et al., 2017), soluble G protein antigenic decoy (Bukreyev et al., 2012), heavy glycosylation (Fuentes et al., 2015; Roe et al., 2022), or modification of T cell trafficking (Harcourt et al., 2006). Given that RSV G appears to leverage its CX3C motif to interact with CX3CR1 and elicit these immune-modulating effects, we hypothesized that we could mitigate this issue by mutating residues in the CCD (Chirkova et al., 2013; Chirkova et al., 2015). However, to consider the consequences of mutating residues in the CCD that might be important for the recognition of neutralizing antibodies, a panel of CCD mutant vaccine candidates was generated by structure-guided mutagenesis using high-resolution crystal structures of mAbs 3D3, 2D10, and 3G12 (Nunez Castrejon et al., 2022). Importantly, their epitopes revealed that mAbs 3D3 and 3G12 all use amino acids located on the N-terminal region of the CCD that were not included in our previous CCD-MP vaccine designs (Fedechkin et al., 2020; Fedechkin et al., 2018). Therefore, the new panel of CCD mutant vaccine candidates included a more extended range of amino acids in the CCD to fully encompass epitopes recognized by neutralizing antibodies (Nunez Castrejon et al., 2022). Our initial investigations into mutated CCDs identified the S177Q mutant, which prevented CX3CR1-mediated chemotaxis similar to the CX4C mutation (Bergeron et al., 2021). Surprisingly, mice vaccinated with the S177Q mutant had robust antibody responses with improved Th1-type responses. Moreover, despite the mutation, S177Q vaccination still mediated CX3C-blocking antibody responses. Finally, human anti-RSV reference sera bind the S177Q mutant with high affinity, similar to binding to WT, and S177Q retains high affinity binding to human and mouse mAbs (Nunez Castrejon et al., 2022). These previous studies demonstrated the translatable feasibility of the S177Q mutation as an immunogenic and safe antigen, which was recapitulated here in the MP vaccine platform.
Based on the previous structural studies which elucidated novel conformational epitopes spanning a larger G peptide region (aa157-198) (Fedechkin et al., 2020; Fedechkin et al., 2018) and our previous results using NP and MP vaccines, we designed RSV G CCD peptide MP vaccines that included the S177Q mutation to improve immunogenicity and correlates of protection (i.e., Th1-type immune responses). S177Q vaccination induced more significant IgG responses, including IgG2a, and there were increased concentrations of Th1-type analytes in the BALF on day 5 pi (i.e., IFNγ and IP-10) compared with WT vaccines. RSV G MP vaccines described here neutralized regardless of C’, and the S177Q mutation induces improved neutralization in vitro and in vivo. Finally, this study was performed in male and female BALB/c mice to improve robustness and because it is known that there can be sex-dependent immune responses to vaccinations and infections (Ursin and Klein, 2021). Our studies reveal that there are subtle intersex responses, but broadly, S177Q vaccination was best in both sexes. While these studies are promising, we are interested in precision medicine to design the most appropriate vaccine candidate for RSV in infants and young children. One limitation of this study is the use of the BALB/c mouse model. While this model suits the immunogenicity and viral challenge studies described here, the BALB/c mouse is semi-permissive to infection and does not fully recapitulate human disease (Taylor, 2017). Future studies should investigate refined vaccine candidates in the cotton rat model, considered a gold-standard RSV animal model for evaluating RSV pathogenesis and VERD. We intend to build upon the studies shown here to develop LbL-MP RSV G vaccines, as the benefits of this vaccine platform include improved uptake by APCs, resulting in greater immunogenicity. This immunogenic vaccine platform may be tailored to those who are ineligible for RSV vaccines and, therefore, close existing gaps in protection.
Conclusions
A significant gap in RSV G protein-based vaccines is the relatively modest immunogenicity and potential for reactogenicity that coincides with Th2-type biased immunity. Our findings show that vaccination with a rationally designed G protein CCD mutant (i.e., S177Q) is non-inferior to WT vaccines in the context of immunogenicity (Figs. 2 and 4) or neutralization (Figs. 3 and 5), which is important as other G protein mutations (including CX4C vaccines) fail to induce effective antibody responses. S177Q vaccination results in higher Th1-type cytokine responses than WT vaccination (Fig. 6), suggesting a reduced likelihood of VERD. Taken together, these data support the translational potential of RSV G S177Q MP vaccines as efficacious and safe RSV vaccines.
Footnotes
Acknowledgments
The authors gratefully acknowledge Edwin Cardenas, Andrea Jacobs, and Jie Tang for providing reagents and conducting mouse studies.
Authors’ Contributions
H.C.B. designed and carried out experiments and helped write and edit the article. H.C.B. and J.M. were involved in data acquisition. L.P.J. and T.J.P. provided reagents. R.M.D. and R.A.T. secured funding, guided the project, and wrote and edited the article. All authors contributed to the article and approved the submitted version. All authors have read and agreed to the published version of the article.
Institutional Review Board Statement
The Institutional Review Board of the University of Georgia, A2021 03–006-Y2-A0, approved the animal use protocol, Immunity to Respiratory Viruses and Virus Proteins in Mus Musculus, May 6, 2021.
Data Availability Statement
Data supporting reported results can be found in the Tripp laboratory at the University of Georgia and archived on the intranet in the Animal Health Research Center at UGA.
Author Disclosure Statement
The authors declare no conflicts of interest.
Funding Information
This research was funded by the Georgia Research Alliance (GRA) and from NIH, grant number 1R01AI166066. NIH, grant number 1R01AI166066 supported the APC.
Supplementary Material
Supplementary Figure S1
Supplementary Figure S2
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.