
Abstract
Interleukin-35 (IL-35) has an immunosuppressive function through the regulation of immune cells during infectious diseases, autoimmune disorders, and cancers. The modulatory role of IL-35 in T lymphocytes, which are involved in host immune responses during human immunodeficiency virus-1 (HIV-1) infection, has not been elucidated. The aim of the current study was to investigate the role of regulatory function of IL-35 to T-cell activity in patients living with chronic HIV-1 infection. Sixty-seven patients living with chronic HIV-1 infection and 17 controls were enrolled in the study. IL-35 levels were measured via an enzyme-linked immunosorbent assay. Purified CD4+ and CD8+ T cells were stimulated with recombinant human IL-35. The secretion of cytokines and cytotoxic molecules, the mRNA levels of IL-35 receptor subunits and transcription factors, the expression of immune checkpoint molecules, and cell proliferation were assessed to evaluate the effect of IL-35 on T lymphocyte function in vitro. Compared with controls, patients living with chronic HIV-1 infection presented increased plasma IL-35 levels. IL-35 stimulation did not affect either the expression of IL-35 receptor subunits or the proliferation of CD4+ and CD8+ T cells from either patients living with chronic HIV-1 infection or controls. IL-35 stimulation downregulated transcription factor mRNA expression and cytokine secretion by CD4+ T cells as well as cytotoxic molecule production by CD8+ T cells from both patients living with chronic HIV-1 infection and controls. This process was accompanied by increased expression of immune checkpoint molecules on CD4+ and CD8+ T cells. The addition of IL-35 also reduced perforin and granzyme B secretion by HIV-1-specific CD8+ T cells from patients living with chronic HIV-1 infection. Increased plasma IL-35 in patients living with chronic HIV-1 infection might dampen the activation of CD4+ and CD8+ T cells, leading to T-cell exhaustion.
Introduction
Human immunodeficiency virus-1 (HIV-1) infects CD4+ T cells and slowly obliterates the host immune response over time (Fenwick et al., 2019). Owing to the reduced numbers and functions of CD4+ T cells, their antigen-presenting ability and protective recall responses to memory CD8+ T cells by CD4+ T cells are impaired during chronic HIV-1 infection (Whitmire, 2011). Moreover, chronic HIV-1 infection can also lead to CD8+ T-cell dysfunction or exhaustion due to persistent HIV-1 viremia and sustained antigen stimulation (Jones and Walker, 2016).
The gene expression profiles of CD4+ and CD8+ T cells also change significantly during HIV-1 infection (Gao et al., 2018). Thus, the reprogramming or restoration of T-cell function, combined with the administration of antiretroviral therapy and other available immunomodulators, may promote the ability of adoptive cell therapy to potentially cure HIV-1 infection (Perdomo-Celis et al., 2022).
Interleukin-35 (IL-35) is a newly identified IL-12 cytokine family member, and is a heterodimeric protein that includes Epstein–Barr virus-induced gene 3 and the IL-12 p35 chain (Collison et al., 2007; Niedbala et al., 2007). The main sources of IL-35 include CD4+CD25+Forkhaed box protein P3 (FoxP3)+ regulatory T cells (Tregs), regulatory CD8+ T cells, and regulatory B cells (Sawant et al., 2015; Wang et al., 2018b). The IL-35 receptor also consists of two heterodimeric subunits, IL-12 receptor β2 (IL-12Rβ2) and gp130 (Collison et al., 2012).
The IL-35 receptor subunits are expressed on immune cells, including T and B lymphocytes, monocytes, and macrophages (Collison et al., 2012). Signaling through the IL-35/IL-35 receptor pathway mediates immunosuppression by inhibiting effector T-cell activity and enhancing of regulatory cell function (Whitehead et al., 2012; Zhang et al., 2019).
The IL-35 expression profile differs between acute and chronic viral infection (Teymouri et al., 2018). Enterovirus 71 and respiratory syncytial virus infection induce the downregulation of peripheral IL-35, leading to an imbalance between Tregs and Th17 cells during acute infection (Fan et al., 2018; Huang et al., 2017). In contrast, the serum IL-35 level is also increased in acute hepatitis B virus (HBV) infection (Ribeiro et al., 2020; Teng et al., 2019), which contributes to the suppression of liver inflammation through dampening Th17 cell activity (Teng et al., 2019). Furthermore, the increase in circulating IL-35 also reveals an important immunosuppressive function during chronic HBV and hepatitis C virus (HCV) infection (Cheng et al., 2018; Li et al., 2015; Liu et al., 2017; Shao et al., 2017; Xiang and Xie, 2015; Yang et al., 2019a; Zhou et al., 2015).
However, few studies have focused on the expression of IL-35 in patients living with HIV-1 infection. Owing to the immunosuppressive effect of IL-35 on T-cell activity, we hypothesized that IL-35 also regulates T-cell function during chronic HIV-1 infection. To test this hypothesis, we first analyzed the IL-35 expression profile in the peripheral blood of patients living with chronic HIV-1 infection. We then assessed the modulatory effect of the recombinant human IL-35 protein on CD4+ and CD8+ T cells from patients living with chronic HIV-1 infection and healthy individuals in vitro.
Methods
Institutional review boards
The study protocol was approved by Lanzhou Pulmonary Hospital Institutional Review Boards (Approval No. 2020–198-07[K]). Informed consent was obtained from each subject according to the guidelines for conducting clinical research.
Patients and controls
Sixty-seven patients living with chronic HIV-1 infection (45 males and 22 females; age: 42.96 ± 7.65 years) and 17 healthy individuals (12 males and 5 females; age: 42.76 ± 8.47 years) were enrolled between January 2021 and March 2023. All patients living with chronic HIV-1 infection were treatment-naïve. Patients who were coinfected with hepatitis virus or who were afflicted with autoimmune diseases or cancers were excluded from this study. The laboratory values for patients living with chronic HIV-1 infection were as follows: HIV-1 RNA load, 6.28 ± 2.01 log10 copies/mL; CD4+ T-cell count, 70 (51, 125)/μL; and CD8+ T-cell count, 226(151, 363)/μL.
Cell separation and culture
Peripheral blood samples were collected from all enrolled study individuals and centrifuged to separate the plasma. Peripheral blood mononuclear cells (PBMCs) were purified from each whole blood sample via Ficoll-Hypaque (Solarbio, Beijing, China) density gradient centrifugation. CD4+ T cells were positively selected with Human CD4 Microbeads (Miltenyi Biotec, Bergisch Galdbach, Germany), whereas CD8+ T cells were negatively selected with a Human CD8+ T-Cell Isolation Kit (Miltenyi Biotec). The CD4+ and CD8+ T-cell purity was >95%, as determined by flow cytometry analysis via fluorochrome-conjugated antibodies against CD3, CD14, and CD19.
In total, 105 of CD4+ or CD8+ T cells, which were separated from 21 patients living with HIV-1 infection and 12 controls, were cultured in complete Roswell Park Memorial Institute (RPMI) 1640 supplemented with 10% fetal calf serum (FCS) and antibodies against CD3/CD28 (final concentration: 100 ng/mL) in the presence or absence of recombinant human IL-35 protein [Peprotech, Rocky Hill, NJ, USA; final concentration: 1 ng/mL (Gao et al., 2022; Yang et al., 2019b)] for 48 h.
Enzyme-linked immunosorbent assay
The levels of IL-35 in the plasma samples as well as the levels of interferon-γ, IL-4, IL-9, IL-17A, IL-22, IL-10, perforin, granzyme B, and granulysin in the supernatants were measured via enzyme-linked immunosorbent assay Kits (CUSABIO, Wuhan, Hubei Province, China).
Cell proliferation
Cell proliferation was measured with a Cell Counting Kit-8 (CCK-8) (Beyotime, Wuhan, Hubei Province, China).
Quantitative real-time polymerase chain reaction
Total RNA was extractedvia TRIzol reagent (Invitrogen, Carlsbad, CA, USA) and reverse transcribed into cDNAvia the PrimeScript RT Reagent Kit (TaKaRa, Beijing, China). Real-time polymerase chain reaction was then performed using TB Green Premix Taq (TaKaRa). The primers for IL-12Rβ2 and gp130 were purchased from Bio-Rad (Hercules, CA, USA). The primers for the transcription factors of CD4+ T cells were obtained from published literature (Sun and Xing, 2021). The relative expression of target genes was determined via normalization to the expression of glyceraldehydes 3-phosphate dehydrogenase to calculate the fold change in expression. Gene expression was calculated via the comparative ΔΔCt method.
Flow cytometry
The cells were stained with anti-CD3-peridinin-chlorophyll-protein complex, anti-CD4 or CD8-allophycocyanin, anti-cytotoxic T lymphocyte-associated protein-4 (CTLA-4)-phycoerythrin, and anti-programmed death-1 (PD-1)-fluorescein isothiocyanate (BD Pharmagen, San Jose, CA, USA) antibodies. Isotype control antibodies were also used for the separation of positive and negative cells. Cell data were acquired on a fluorescence-activated cell sorting (FACS) Calibur flow cytometer (BD Biosciences, San Jose, CA, USA), and the FACS data were analyzed via FlowJo software.
Enzyme-linked immunospot assay
In total, 106 PBMCs from 12 human leukocyte antigen (HLA)-A*02-restricted patients living with chronic HIV-1 infection were stimulated with the HLA-A*02-restricted gag p17 epitope (SLYNTVATL) (Vali et al., 2011) (final concentration: 10 μg/mL) and the HLA-A*02-restricted gag p24 epitope (TLNAWVKVV) (Schaubert et al., 2007) (final concentration: 10 μg/mL) in the presence or absence of recombinant human IL-35 protein (final concentration: 1 ng/mL) for 24 h as we previously described (Zhou et al., 2024). The secretion of perforin and granzyme B by HIV-1-specific CD8+ T cells was measured via the Human Perforin (enzyme-linked immunospot assay, ELISPOT) Kit (Abcam, Cambridge, CA, USA) and the Human Granzyme B ELISPOT Kit (Abcam), respectively. The results are presented as the number of spot-forming cells.
Statistical analysis
The data were analyzed via SPSS 25.0 software. Student’s t test, paired t test, one-way analysis of variance (ANOVA), or least significant difference (LSD)-t test were used to assess the statistical significance of differences in normally distributed data, and these data are presented as the means±standard deviations. Relationships between variables were tested via the Pearson correlation test. Mann–Whitney tests, Wilcoxon ranked paired tests, Kruskal–Wallis tests, or Dunn’s multiple tests were employed to assess the statistical significance of differences in data that followed a skewed distribution, and these data are presented as the median (Q1, Q3). Relationships between variables were tested via the Spearman rank correlation test. All the tests were two-tailed, and p values of less than 0.05 were considered significant.
Results
The plasma IL-35 level was increased in patients living with chronic HIV-1 infection
The plasma IL-35 level was significantly increased in patients living with chronic HIV-1 infection than in healthy controls (0.87 ± 0.27 ng/mL vs 0.69 ± 0.12 ng/mL, p = 0.012, Fig. 1A). However, the plasma IL-35 level did not correlate with the HIV RNA load (r = −0.063, p = 0.613, Fig. 1B), CD4+ T-cell count (r = −0.093, p = 0.456, Fig. 1C), or CD8+ T-cell count (r = 0.067, p = 0.592, Fig. 1D) in patients living with chronic HIV-1 infection.
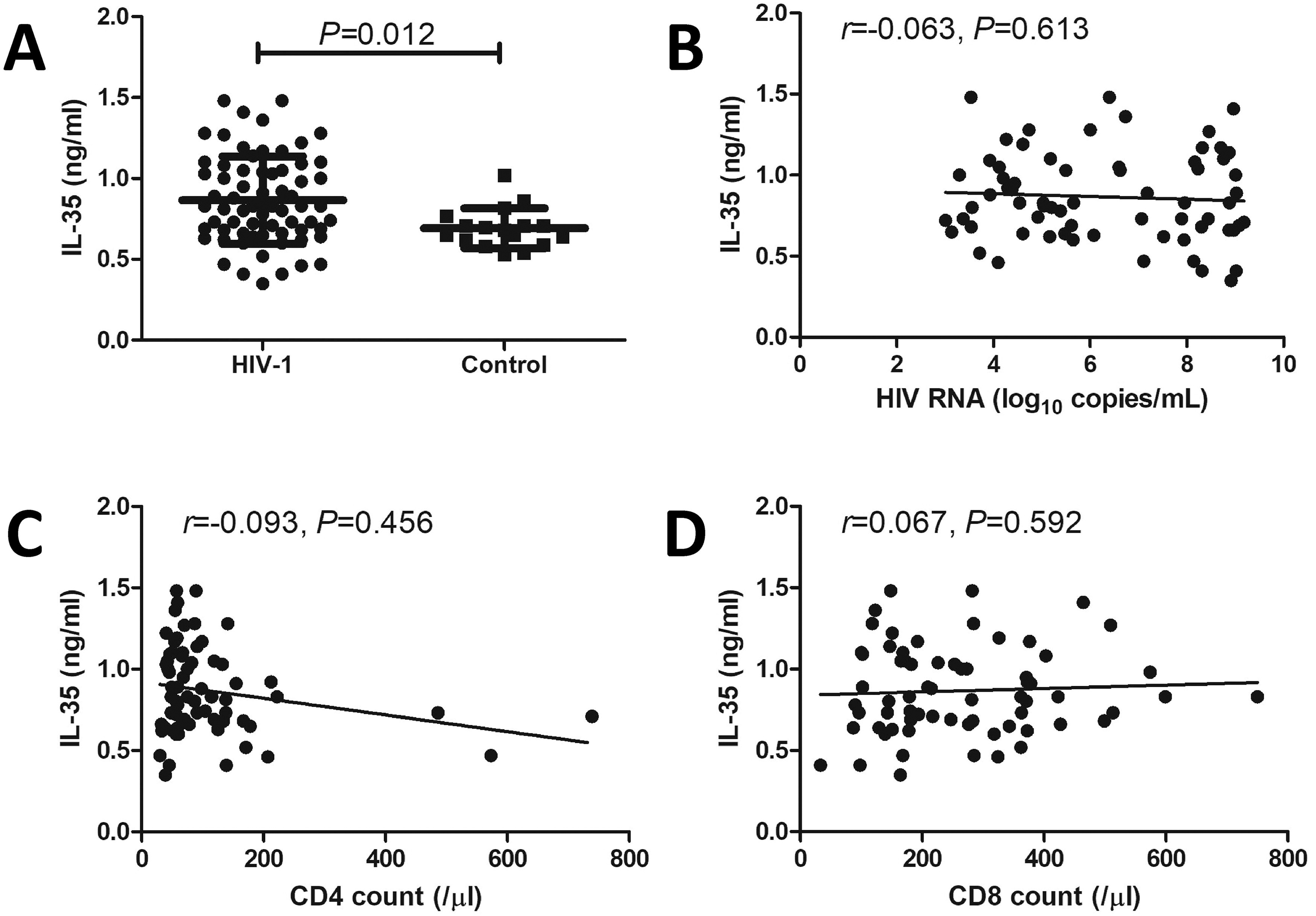
Expression of IL-35 and its correlation with clinical indices in patients living with chronic HIV-1 infection. Plasma IL-35 levels in patients living with chronic HIV-1 infection (n = 67) and controls (n = 17) were measured via ELISA.
IL-35 stimulation in vitro did not affect cell proliferation or IL-35 receptor subunit expression in CD4+ and CD8+ T cells
There was no remarkable difference in total cell count between cells with and without IL-35 stimulation in either the CD4+ T-cell culture (all p > 0.05, Fig. 2A) or the CD8+ T-cell culture (all p > 0.05, Fig. 2B). There were also no significant differences in the mRNA expression of IL-35 receptor subunits (IL-12Rβ2 and gp130) in CD4+ and CD8+ T cells between patients living with chronic HIV-1 infection and controls (all p > 0.05, Fig. 2C– F). IL-35 stimulation in vitro did not affect IL-35 receptor subunit mRNA expression in either CD4+ or CD8+ T cells (all p > 0.05, Fig. 2C–F).
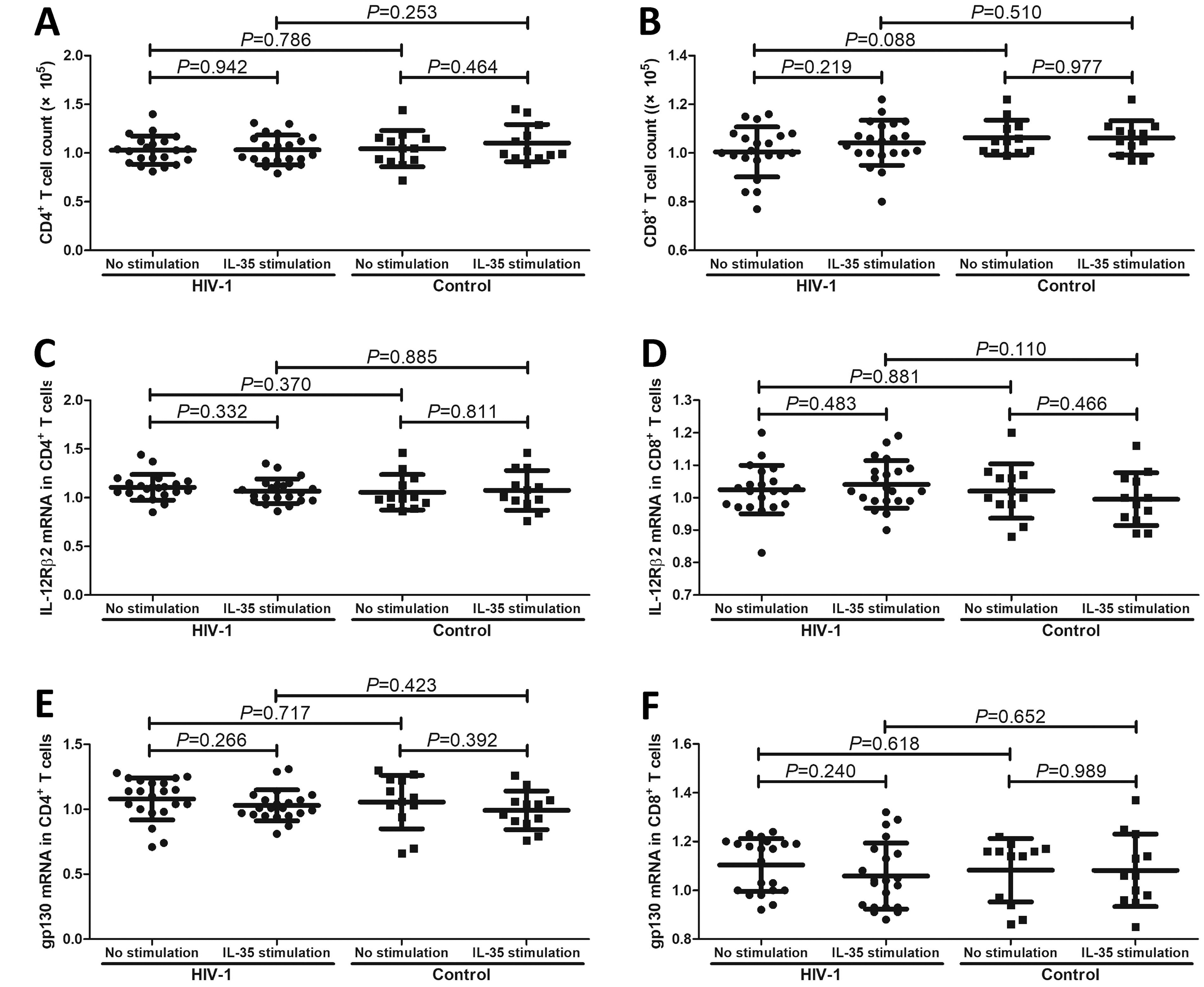
Influence of recombinant human IL-35 protein stimulation on cell proliferation and IL-35 receptor subunit mRNA expression in CD4+ and CD8+ T cells from patients living with chronic HIV-1 infection and controls. CD4+ T and CD8+ T cells were separated from patients living with chronic HIV-1 infection (n = 21) and controls (n = 12). In total, 105 of purified CD4+ T and CD8+ T cells were cultured with anti-CD3/CD28 antibodies (final concentration: 1 ng/mL) to maintain T-cell activity in the presence or absence of recombinant human IL-35 protein (final concentration: 1 ng/mL) for 48 h. Cell proliferation was measured with a Cell Counting Kit-8. The mRNA expression levels of IL-35 receptor subunits, including IL-12Rβ2 and gp130, were semiquantified via real-time PCR. There was no remarkable difference in
IL-35 stimulation in vitro promoted CTLA-4 and PD-1 expression on CD4+ and CD8+ T cells
Representative flow dot analyses of CTLA-4 and PD-1 expression on CD4+ or CD8+ T cells with or without IL-35 stimulation are shown in Figure 3A. The percentages of CTLA-4+ T cells among CD4+ T cells (p = 0.002, Fig. 3B), CTLA-4+ T cells among CD8+ T cells (p = 0.001, Fig. 3D), and PD-1+ T cells among CD8+ T cells (p = 0.012, Fig. 3E) were significantly higher in those from patients living with chronic HIV-1 infection than in controls. The proportion of PD-1+CD4+ T cells was also higher in those from patients living with chronic HIV-1 infection, but this difference just failed to achieve statistical significance (p = 0.059, Fig. 3C). IL-35 stimulation in vitro strongly promoted CTLA-4 and PD-1 expression on CD4+ and CD8+ T cells from both patients living with HIV-1 infection and controls (all p < 0.05, Fig. 3A–D).
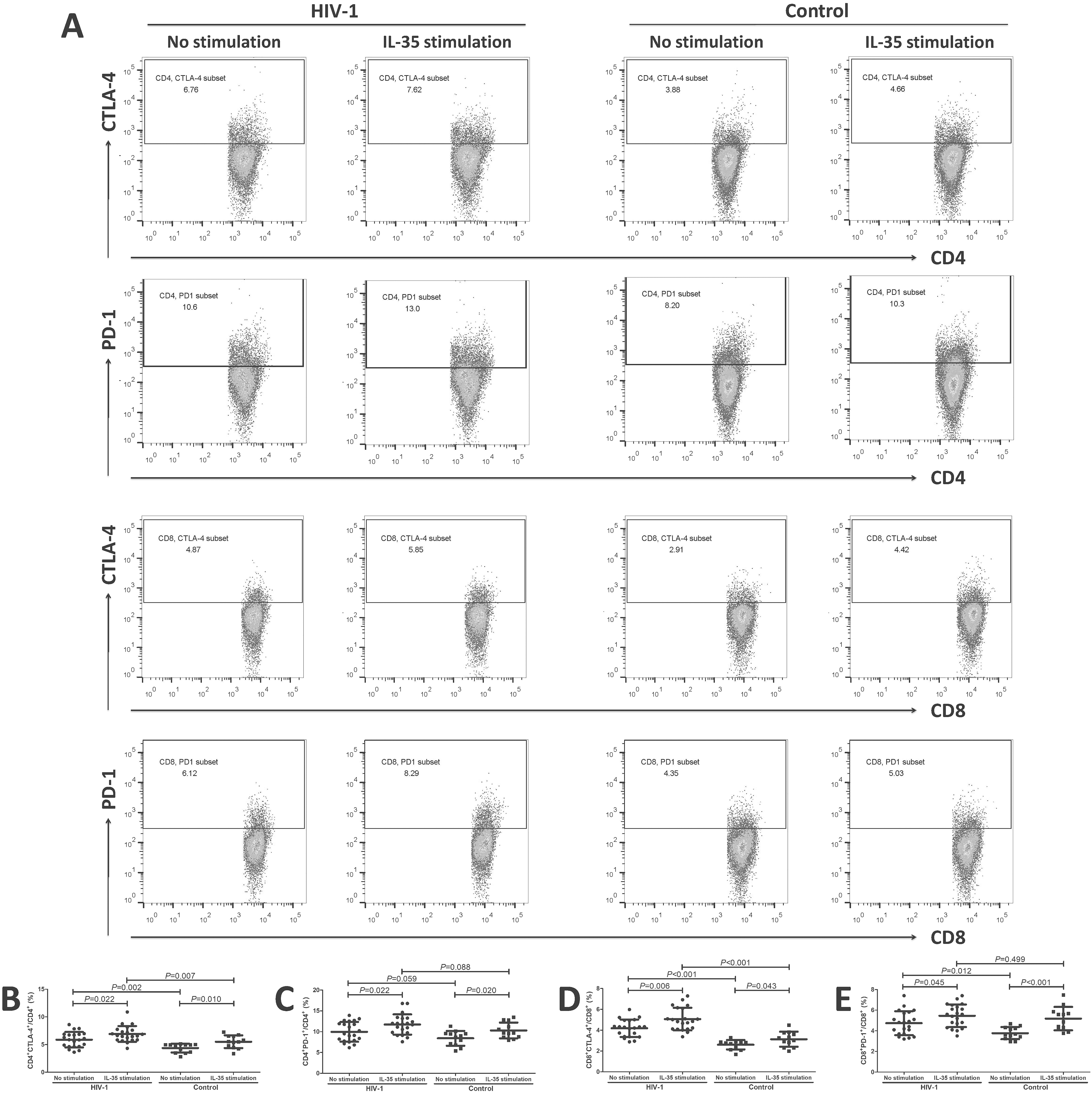
Influence of recombinant human IL-35 protein stimulation on CTLA-4 and PD-1 expression on CD4+ and CD8+ T cells from patients living with chronic HIV-1 infection and controls. CD4+ T and CD8+ T cells were separated from patients living with chronic HIV-1 infection (n = 21) and controls (n = 12). In total, 105 of purified CD4+ T and CD8+ T cells were cultured with anti-CD3/CD28 antibodies (final concentration: 1 ng/mL) to maintain T-cell activity in the presence or absence of recombinant human IL-35 protein (final concentration: 1 ng/mL) for 48 h. The expression of CTLA-4 and PD-1 on CD4+ or CD8+ T cells was measured via flow cytometry.
IL-35 stimulation in vitro downregulated transcription factor mRNA expression and cytokine secretion by effector CD4+ T cells
The mRNA expression of transcription factors in effector CD4+ T cells, including T-box expressed in T cell (T-bet) (Th1 transcription factor), GATA binding protein-3 (GATA-3) (Th2 transcription factor), purine-rich nucleic acid binding protein 1 (PU.1) (Th9 transcription factor), retinoid-related orphan receptor gamma t (RORγt) (Th17 transcription factor), and aryl hydrocarbon receptor (AhR) (Th22 transcription factor), was significantly lower in CD4+ T cells from patients living with chronic HIV-1 infection than in those from controls (all p < 0.05, Fig. 4A–E). IL-35 stimulation in vitro strongly downregulated T-bet, GATA-3, PU.1, RORγt, and AhR mRNA expression in CD4+ T cells from both patients living with HIV-1 infection and controls (all p < 0.05, Fig. 4A–E). In contrast, FoxP3 mRNA expression in Tregs was notably elevated in CD4+ T cells from patients living with HIV-1 infection compared with those from controls (p = 0.009, Fig. 4F). IL-35 stimulation in vitro promoted FoxP3 mRNA expression in CD4+ T cells from both patients living with HIV-1 infection and controls (all p < 0.05, Fig. 4F).

Influence of recombinant human IL-35 protein stimulation on transcription factor mRNA expression in CD4+ T cells from patients living with chronic HIV-1 infection and controls. CD4+ T cells were separated from patients living with chronic HIV-1 infection (n = 21) and controls (n = 12). In total, 105 of purified CD4+ T cells were cultured with anti-CD3/CD28 antibodies (final concentration: 1 ng/mL) to maintain T cell activity in the presence or absence of recombinant human IL-35 protein (final concentration: 1 ng/mL) for 48 h. The mRNA expression of transcription factors in CD4+ T cells was semiquantified by real-time PCR. mRNA expression of
The secretion of cytokines by effector CD4+ T cells, including the Th1 cytokine interferon-γ, the Th2 cytokine IL-4, the Th9 cytokine IL-9, the Th17 cytokine IL-17A, and the Th22 cytokine IL-22, were remarkably lower in the supernatants of cultured CD4+ T cells from patients living with HIV-1 infection than in those from controls (all p < 0.05, Fig. 5A–E). IL-35 stimulation in vitro significantly downregulated interferon-γ, IL-4, IL-9, IL-17A, and IL-22 secretion by CD4+ T cells from both patients living with HIV-1 infection and controls (all p < 0.05, Fig. 5A–E). In contrast, the Tregs cytokine IL-10 was remarkably increased in CD4+ T cells from patients living with HIV-1 infection compared with those from controls (p = 0.002, Fig. 5F). IL-35 stimulation in vitro increased IL-10 secretion by CD4+ T cells from both patients living with HIV-1 infection and controls (all p < 0.05, Fig. 5F).

Influence of recombinant human IL-35 protein stimulation on cytokine secretion by CD4+ T cells from patients living with chronic HIV-1 infection and controls. CD4+ T cells were separated from patients living with chronic HIV-1 infection (n = 21) and controls (n = 12). In total, 105 of purified CD4+ T cells were cultured with anti-CD3/CD28 antibodies (final concentration: 1 ng/mL) to maintain T cell activity in the presence or absence of recombinant human IL-35 protein (final concentration: 1 ng/mL) for 48 h. The cytokine levels in the supernatants were measured via ELISA.
IL-35 stimulation in vitro reduced cytotoxic molecule production in CD8+ T cells from patients living with chronic HIV-1 infection
The secretion of perforin, granzyme B, granulysin, and interferon-γ by CD8+ T cells was significantly lower in cells from patients living with chronic HIV-1 infection than in those from controls (all p < 0.01, Fig. 6A–D). IL-35 stimulation in vitro notably downregulated perforin, granzyme B, granulysin, and interferon-γ production by CD8+ T cells from both patients living with chronic HIV-1 infection and controls (all p < 0.05, Fig. 6A–D). Furthermore, a total of 106 PBMCs from 12 HLA-A*02-resctricted patients living with chronic HIV-1 infection were stimulated with HLA-A*02-restricted gag peptides and recombinant human IL-35 for 24 h. IL-35 stimulation strongly dampened both perforin and granzyme B secretion by HIV-1-specific CD8+ T cells (p < 0.05, Fig. 7A and B).

Influence of recombinant human IL-35 protein stimulation on cytotoxic molecule secretion by CD8+ T cells from patients living with chronic HIV-1 infection and controls. CD8+ T cells were separated from patients living with chronic HIV-1 infection (n = 21) and controls (n = 12). In total, 105 of purified CD8+ T cells were cultured with anti-CD3/CD28 antibodies (final concentration: 1 ng/mL) to maintain T-cell activity in the presence or absence of recombinant human IL-35 protein (final concentration: 1 ng/mL) for 48 h. The levels of cytotoxic molecules in the supernatants were measured via ELISA.

Influence of recombinant human IL-35 protein stimulation on perforin and granzyme B secretion by HIV-1-specific CD8+ T cells from patients living with chronic HIV-1 infection. In total, 106 PBMCs from 12 HLA-A*02-restricted patients living with chronic HIV-1 infection were stimulated with the HLA-A*02-restricted gag p17 epitope (SLYNTVATL) (final concentration: 10 μg/mL) and the HLA-A*02-restricted gag p24 epitope (TLNAWVKVV) (final concentration: 10 μg/mL) in the presence or absence of recombinant human IL-35 protein (final concentration: 1 ng/mL) for 24 h. Perforin and granzyme B secretion was measuredviathe ELISPOT assay. IL-35 stimulation dampened both
Discussion
To the best of our knowledge, this is the first report on the expression profile of IL-35 and the direct regulatory function of IL-35 in T cells during chronic HIV-1 infection. Our current results revealed that patients living with chronic HIV-1 infection had robustly increased peripheral IL-35 levels compared with healthy individuals, but this elevation failed to correlate with either viral replication or T-cell counts. Similar to our previous study (Zhou et al., 2024), we also showed that patients living with chronic HIV-1 infection presented reduced CD4+ T-cell activation and weaker CD8+ T-cell cytotoxicity.
IL-35 receptor subunits are expressed on both CD4+ and CD8+ T cells. Thus, IL-35 stimulation can directly modulate T-cell function. On the one hand, IL-35 stimulation suppressed effector T-cell responses but promoted the Treg response in cultured CD4+ T cells from both patients living with chronic HIV-1 infection and controls. On the other hand, IL-35 stimulation dampened both nonspecific and HIV-1-specific CD8+ T-cell cytotoxicity. These processes are accompanied by the upregulation of immune checkpoint molecule expression on T cells without affecting cell proliferation or IL-35 receptor expression. Taken together, the data obtained in the present study demonstrated an immunosuppressive effect of IL-35 on T cells, which might play an essential role in immune dysfunction and persistent viral activity during chronic HIV-1 infection.
IL-35 functions as a crucial immunosuppressive factor in the majority of immune-mediated diseases, including chronic parasitic, bacterial, and viral infections (Teymouri et al., 2018; Xiang and Xie, 2015). Increased serum IL-35 levels were detected in patients with Schistosoma haematobium or Schistosoma mansoni infection, whereas effective antiparasitic therapy robustly reduced the IL-35 concentration in chronic human Schistosoma infection (Marascio et al., 2023). Similarly, increased serum IL-35 and the IL-35-producing regulatory T-cell subset in the spleen increase the bacterial burden and lung lesions in chronic Mycobacterium tuberculosis-infected mice (Yu et al., 2022). Importantly, elevated IL-35 has been shown to be important for the progression of chronic HBV infection, the development of cirrhosis, and the transformation and metastasis of hepatocellular carcinoma (Li et al., 2021). The IL-35 level has been demonstrated to be strongly increased in patients with chronic hepatitis virus infection (Cheng et al., 2018; Liu et al., 2017; Shao et al., 2017), and IL-35 promoted HBV transcription and replication as well as antigen secretion in the an HBV-transgenic mouse model (Tao et al., 2018).
Herein, we showed that plasma IL-35 level was also remarkably increased in patients living with chronic HIV-1 infection. However, the IL-35 concentration did not correlate with HIV RNA load. This finding was partly consistent with previous findings that IL-35 had no effect on HIV-1 replication (Chen et al., 2013). Although there was also no statistical correlation between IL-35 levels and T-cell counts in patients living with chronic HIV-1 infection, the regulatory role of IL-35 in T-cell function during chronic HIV-1 infection still needs further elucidation.
Naïve CD4+ T cells differentiate into several effector and regulatory subsets, including Th1, Th2, Th9, Th17, Th22, and Tregs, in response to transcription factor and cytokine activation. The regulatory effects of IL-35 on CD4+ T-cell subsets under different pathological conditions remain controversial. Treatment of CD4+ T cells from Guillain–Barré syndrome patients with recombinant human IL-35 induced the upregulation of T-bet, GATA-3, and FoxP3 and inhibited RORγt expression, indicating that IL-35 promoted Th1, Th2, and Treg responses while dampening the Th17 response in Guillain–Barré syndrome pathogenesis (Liu et al., 2015). Recombinant human IL-35 treatment also induced Treg differentiation and suppressed the Th17 response in patients with systemic sclerosis (Yang et al., 2022). Exogenous IL-35 stimulation inhibited Th1/Th17 cell activation in only the tumor microenvironment, without affecting peripheral CD4+ T-cell proliferation, activation, or differentiation in nonsmall cell lung cancer patients (Wang et al., 2018a).
In the present study, we found that effector CD4+ T cells from patients living with chronic HIV-1 infection presented reduced activation, whereas Tregs seemed to be overactivated during chronic HIV-1 infection. Furthermore, in vitro IL-35 stimulation inhibited the effector CD4+ T-cell response in both patients living with chronic HIV-1 infection and healthy individuals. Tregs also respond to exogenous IL-35 stimulation in both patients living with chronic HIV-1 infection and healthy individuals, suggesting the potential positive feedback of IL-35 to Tregs, which are among the main sources of IL-35, under both pathological and physiological conditions. Our current results are consistent with findings in patients with Kawasaki disease (Sun and Xing, 2021). This finding further demonstrated the suppressive function of IL-35 in effector CD4+ T cells and that the ability of IL-35 to enhance Treg function contributed to immunotolerance in chronic infection, leading to viral persistence.
CD8+ T cells contribute to controlling HIV-1 infection through major histocompatibility complex-1-dependent processes and the perforin-granzyme signaling pathway (Gulzar and Copeland, 2004). Our previous study also demonstrated the exhausted phenotype of peripheral CD8+ T cells in patients with chronic HIV-1 infection (Zhou et al., 2024). Although we did not find a significant correlation between the IL-35 level and the CD8+ T-cell count in patients living with chronic HIV-1 infection, in vitro IL-35 stimulation also suppressed the cytotoxicity of both nonspecific and HIV-1-specific CD8+ T cells. CD8+ T cells exhibit cytotoxicity through two different mechanisms, including direct cell-to-cell contact dependent, cytotoxicity molecule (especially perforin, granzyme B, and granulysin)-mediated cell lysis and cytokine (mainly interferon-γ)-induced cell necrosis, or apoptosis in an indirect contact manner (Hay and Slansky, 2022; Iwahori, 2020; Saeidi et al., 2015). We showed that IL-35 stimulation inhibited the secretion of both cytotoxic molecules and interferon-γ from CD8+ T cells, indicating that increased peripheral IL-35 levels might dampen the direct cytolytic and cytokine-mediated noncytolytic activities of CD8+ T cells during chronic HIV-1 infection.
The potential mechanisms of IL-35 regulation in T lymphocytes were also investigated. First, although previous reports indicated the inhibitory effect of IL-35 on the proliferation of parenchymal cells and immune cells (Li et al., 2016; Osuna-Gómez et al., 2023), our current in vitro results revealed that IL-35 stimulation did not affect either CD4+ or CD8+ T-cell proliferation. Second, IL-35 stimulation did not affect IL-35 receptor subunit expression in CD4+ or CD8+ T cells, indicating that IL-35 might not amplify signaling pathway molecules. Third, IL-35 stimulation strongly promoted the expression of immune checkpoint molecules on CD4+ and CD8+ T cells, suggesting that IL-35 might contribute mainly to immunosuppressive activity through increasing the expression of inhibitory receptors on immune cells. Further in vivo studies are needed to ascertain the regulatory roles and mechanisms of IL-35 in T lymphocyte function during chronic HIV-1 infection.
There were limitations of the current study. First, the experiments were solely performed on CD8+ T cells due to the limited HLA-A*02-restricted patients enrolled. A large number of cases could be analyzed to obtain a suitable statistic number to compare. Second, Tregs and other cell types (including activated myeloid, endothelial cells, regulatory B cells) could secret IL-35. However, there was no commercial fluorescent-labeled antibody targeting IL-35 available for flow cytometry analysis. Thus, we cannot detect CD4+IL-35+ T cells by flow cytometry. Further in vivo experiments were needed to confirm the in vitro results.
Conclusion
In summary, the increased plasma IL-35 level in patients living with chronic HIV-1 infection might dampen the activation of CD4+ and CD8+ T cells, leading to T-cell exhaustion. IL-35 signaling might be a potential therapeutic target for the functional cure of chronic HIV-1 infection.
Footnotes
Author Disclosure Statement
No competing financial interest exists.
Authors’ contributions
N.L.: Conceptualization, data curation, formal analysis, investigation, methodology, resources, software, validation, visualization, funding acquisition, writing—original draft. C.T.: Conceptualization, data curation, formal analysis, investigation, methodology, resources, software, validation, visualization, writing—original draft. Y.C.: Conceptualization, data curation, formal analysis, investigation, methodology, resources, software, validation, visualization, writing—review and editing. Z.Y.: Conceptualization, data curation, formal analysis, investigation, methodology, resources, software, validation, visualization, writing—review and editing. Y.Z.: Conceptualization, data curation, formal analysis, investigation, methodology, project administration, resources, software, supervision, validation, visualization, writing—review and editing.
Funding Information
This work was supported by a grant from the Science and Technology Development Plan Project of Lanzhou City (No. 2024-4-43).