
Abstract
SARS-CoV-2 has evolved into several variants of concern, with Omicron and its subvariants currently being the most prevalent. Previously, we developed a mouse monoclonal antibody (m1E3H12 mAb) specific to the receptor binding domain of SARS-CoV-2 Omicron spike protein, and the mAb showed neutralizing activity against SARS-CoV-2 Omicron BA.1 and its subvariants BA.5, BQ.1.1, and XBB. Here, we showed that the mAb provided protection against SARS-CoV-2 Omicron infection in K18-hACE2 transgenic mice when administered intranasally. The mAb treatment reduced viral loads in both the brain and lungs. Additionally, the elevated levels of RANTES (CCL5) and MIP-3 alpha (CCL20) in the brain following SARS-CoV-2 Omicron infection showed a decreasing trend after mAb treatment. Therefore, we conclude that our mAb specific to SARS-CoV-2 Omicron spike protein has the potential to be applied as therapeutics against SARS-CoV-2 Omicron BA.1 and its subvariants BA.5, BQ.1.1, and XBB.
Introduction
On 5 May 2023, the World Health Organization announced the conclusion of the coronavirus disease 2019 (COVID-19) as a global health emergency (WHO, 2023). However, the Omicron variant (B.1.1.529) was first detected in November 2021, and this variant has spread rapidly across the world, evolved subvariants (CDC, 2024) and raised serious concerns about its effects on vaccine efficacy and therapeutic options (Ao et al., 2023).
Since the emergence and rapid spread of SARS-CoV-2, scientific efforts have been intensified to develop therapeutic solutions. The therapeutic monoclonal antibodies (mAbs), such as bamlanivimab—etesevimab, bebtelovimab, casirivimab—imdevimab, sotrovimab, and tixagevimab—cilgavimab, have gained significant attention due to FDA authorization or approval for their use in combating this formidable virus (Toussi et al., 2023). These antibodies, which are specifically designed to bind to the viral spike (S) protein, have proven effective in mitigating the severity of COVID-19 and reducing hospitalizations. However, the Omicron variant and its subvariants have presented a challenge for existing mAb treatments―except for bebtelovimab―because the Omicron variants have high transmissibility and the highly mutated receptor binding domain (RBD) of the S protein (Takashita et al., 2022; Zhou et al., 2022). Therefore, the development and application of mAbs capable of neutralizing recent Omicron variants are required.
Previously, we reported the development and characterization of a novel mAb (m1E3H12 mAb) that targeted the RBD of the S protein and exhibited potent neutralizing activity against the Omicron variants BA.5, BQ.1.1, and XBB in vitro (Kim et al., 2024b). Here, we elucidated the therapeutic effects of the mAb, depending on the route of administration. Our findings suggested that this mAb will be a valuable addition to the arsenal of therapeutics against COVID-19, especially given the ongoing generation of subvariants of SARS-CoV-2 Omicron.
Materials and Methods
Cell culture and viruses
The National Culture Collection for Pathogens of Korea provided the SARS-CoV-2 Omicron (hCoV-19/Korea/KDCA447321/2021, lineage B.1.1.529, GISAID accession no. EPI_ISL_6959993). The viruses were amplified in Vero E6 cells (American Type Culture Collection) as previously described (Park et al., 2023; Kim et al., 2022). The cell culture supernatants containing amplified viruses were collected, and the infectious viruses were quantified using a certified 50% tissue culture infectious dose (TCID50) assay as previously described (Lei et al., 2021; Baek et al., 2024b). SARS-CoV-2 amplification was performed at biosafety level 3 (BSL3) conditions in the Research Institute of Medical-Bio Convergence of Hallym University.
Animals
Seven-week-old male hemizygous K18-hACE2 transgenic mice (B6.Cg-Tg[K18-hACE2]2Prlmn/J, The Jackson Laboratory) and male C57BL/6 (Nara Biotech) mice were maintained in specific pathogen-free conditions at the Experimental Animal Center of Hallym University. All animal procedures were approved by the Institutional Animal Care and Use Committee of Hallym University (Hallym2022-46, Hallym2023-23). All efforts were made to minimize animal pain and suffering.
Virus challenge experiments
Virus challenge experiments were performed at animal BSL 3 conditions as previously described (Kim et al., 2023b; Kim et al., 2024a; Baek et al., 2024a). Eight-week-old male hemizygous K18-hACE2 transgenic mice were infected (intranasal route) with the SARS-CoV-2 Omicron variant at a dose of 2 × 104 TCID50. From 1 h after infection, the mice were intranasally treated with the m1E3H12 mAb once daily for 3 days (0.25 mg/kg) or intravenously or intraperitoneally treated with the m1E3H12 mAb once daily for 3 days (2.5 mg/kg). A mouse anti-human Mucin 1 monoclonal antibody (Wu et al., 2018) was used as a mouse IgG1 subclass control antibody (control mIgG). Body weight, clinical signs, and survival were recorded every day for 5 or 10 days. At 5- or 8-days post-infection (DPI), the turbinate, lungs, and brains of the infected mice were collected to measure virus levels.
qRT-PCR for measurement of SARS-CoV-2
To detect total SARS-CoV-2 Omicron virus particles in specific mouse tissues, we employed the quantitative reverse transcription PCR (qRT-PCR) method. The turbinate, lungs, and brains collected from the infected mice were homogenized, and the supernatants were harvested. The viral gene copy numbers were evaluated using qRT-PCR. Total RNA was extracted from the supernatants (100 μL), and cDNA (50 μL) was prepared using a Reverse Transcription System Kit (Promega). The primers utilized to quantify the RNA-dependent RNA polymerase (RdRP) gene expression were as follows (Kang et al., 2024): forward primer, 5′-GTGAAATGGTCATGTGTGGCGG-3′, reverse primer 5′-CAAATGTTAAAAACACTATTAGCATA-3′; and TaqMan probe, 5′-FAM-CAGGTGGAACCTCATCAGGAGATGC-BHQ1-3′. Ten microliters of Takara Premix Taq™ (TaKaRa Taq™ Version 2.0) (Takara) were added to a 10-µL reaction mixture containing 125 nM each of the forward and reverse primers, 62.5 nM probe, and 1 μL cDNA. After denaturation of cDNA at 95°C for 5 min, 45 cycles of PCR at 95°C for 15 sec and 60°C for 1 min were performed in a CFX Connect Real-Time PCR Detection System (Bio-Rad). The copy numbers of the RdRP gene were calculated using an RdRP cDNA standard curve.
Hematoxylin and eosin staining
Lung tissues were collected from the infected mice, and then the tissues were embedded in paraffin for histopathological examination as previously described (Baek et al., 2022). The tissues were sectioned into 5-µm-thick slices and then stained with Gill’s Hematoxylin V (Muto Pure Chemicals). After counterstaining with Eosin Y solution (Abcam), the stained sections were analyzed using an Eclipse E200 microscope (Nikon).
Measurement of cytokines and chemokines
Sera and homogenates of turbinate, lungs, and brains of K18-hACE2 transgenic mice (n = 5/group) were collected at 8 DPI. Sera and homogenates of tissues were serially diluted using an assay diluent buffer, and the amounts of cytokines and chemokines were measured using the cytometric bead array mouse Th1/Th2/Th17 cytokine kit (BD Biosciences) and a LEGENDplex™ Mouse Proinflammatory Chemokine Panel (BioLegend) according to the manufacturer’s instructions as previously described (Kim et al., 2024a). The samples were incubated with capture beads, followed by washing and treatment with detection reagents. The results were analyzed using FACSCalibur (BD Biosciences) and quantified with LEGENDplex™ software, version 7.0 (BioLegend).
Analysis of the biodistribution of m1E3H12 mAb in mice
To investigate the localization of m1E3H12 mAb in mice, the m1E3H12 mAb was conjugated with DyLight™ 755 NHS Ester (Thermo Fisher Scientific) according to the manufacturer’s instructions. DyLight 755-labeled m1E3H12 mAb (50 μg/mouse) was administered intranasally, intravenously, or intraperitoneally to 8-week-old male C57BL/6 mice (n = 3). The localization of m1E3H12 mAb was monitored by measuring fluorescence total flux (photons/s). The biodistribution of m1E3H12 mAb was monitored after 1 h using a VISQUE InVivo Smart-LF (Vieworks).
Pharmacokinetics study
The m1E3H12 mAb was conjugated with Z-Link™ Sulfo-NHS-Biotin (Thermo Fisher Scientific) according to the manufacturer’s instructions. The biotin-labeled m1E3H12 mAb (2.5 mg/kg mouse) was administered intranasally to 8-week-old male C57BL/6 mice (n = 5/group). Turbinate, lungs, and sera were collected after 30 min, 1 h, 3 h, 6 h, 12 h, 24 h, 48 h, and 72 h. Each organ was homogenized, and tissue lysates were prepared. ELISA was performed to measure the levels of m1E3H12 mAb in the tissue lysates and sera.
For the enzyme-linked immunosorbent assay (ELISA), 100 ng/well of recombinant SARS-CoV-2 Omicron RBD protein (Omicron B.1.1.529-RBD protein, Genscript) was coated in 96-well immunoplates at 4°C overnight. After blocking the plates with phosphate buffered saline (PBS) containing 1% bovine serum albumin for 1 h, tissue lysates and sera were added to each well and incubated for 2 h at room temperature. To create the standard curve for the calculation of IgG amounts in the samples, serially diluted biotin-labeled m1E3H12 mAb was added to each well. The plates were washed three times with PBS containing 0.1% Tween 20 (PBST) and then incubated with horse raddish peroxidase (HRP)-conjugated streptavidin (Sigma-Aldrich) for 1 h at room temperature. After washing three times with PBST, HRP activity was developed using the 3,3′,5,5′-tetramethylbenzidine (TMB) peroxidase substrate. The amounts of m1E3H12 mAb were determined by measuring absorbance at 450 nm with a Multiskan GO ELISA reader (Thermo Fisher Scientific).
The pharmacokinetics (PK) parameters of the intranasally administered m1E3H12 mAb were calculated using the non-compartmental model in the R package “PK” v1.3 (Jaki and Wolfsegger, 2011). A two-phase linear regression fitting procedure was used to perform the elimination half-life (t1/2) in tissues as previously described (Kim et al., 2023a). The elimination half-life (t1/2), estimated by linear regression, represents the time required for antibody concentration to decrease by 50%. AUClast is the area under the time—residual amount curve from 0 h to 24 h. AUCinf represents the area under the concentration-time curve extrapolated from 0 h to infinity, and mean residence time (MRT) indicates the average time the antibody molecules remain in the system.
Statistical analysis
The results were presented as mean ± standard error of the mean values. Differences between two samples were evaluated using Student’s t-tests; p < 0.05 was the threshold for statistical significance.
Results
Therapeutic effects of m1E3H12 mAb via intranasal treatment against SARS-CoV-2 Omicron infection in a K18-hACE2 mouse model
To estimate the protective effect of the m1E3H12 mAb based on route of administration, we first intranasally infected K18-hACE2 transgenic mice with SARS-CoV-2 Omicron (Fig. 1A). When we infected SARS-CoV-2 Omicron at a dose of 2 × 104 TCID50 in mice, no severe clinical signs, such as weight loss or lethality, were observed for 5 days. However, a gradual increase in mouse mortality was observed from 8 DPI, resulting in a 50% death rate at 10 DPI (Fig. 1B and C). When the mice were intranasally administered with m1E3H12 mAb once daily for 3 days (0.25 mg/kg) after infection, the survival rate was 100% (Fig. 1C). Viral titers in the turbinate (Fig. 1D), lungs (Fig. 1E), and brains (Fig. 1F) were highly elevated at 5 DPI and significantly reduced by intranasal administration of the m1E3H12 mAb. When the viral titers were measured at 8 DPI in survived mice (n = 5), we did not find viral loads in the turbinate (Fig. 1G) or lungs (Fig. 1H). However, measurable viral titers were detected in the brains, and intranasal administration of the m1E3H12 mAb almost completely reduced the increased viral loads (Fig. 1I). H&E staining of lung tissues at 5 and 8 DPI revealed no significant structural or functional impairment in the lungs of mice infected with SARS-CoV-2 Omicron (Fig. 1J). To evaluate whether inflammation contributed to the pathogenesis of SARS-CoV-2 Omicron infection, levels of cytokines and chemokines were estimated in the turbinate, lungs, blood, and brains using a multiplex cytokine array. At 8 DPI, no significant changes were observed in most of the tested cytokines or chemokines in the turbinate or lungs (Supplementary Fig. S1). A similar trend was seen in the brain; however, the expression levels of two chemokines, RANTES (CCL5) and MIP3-Alpha (CCL20), were significantly increased following Omicron infection (p < 0.05, Supplementary Fig. S2). Their expression levels showed a decreasing trend after intranasal administration of the m1E3H12 mAb, though the reduction did not reach statistical significance (p = 0.0578 and p = 0.0664, respectively; Supplementary Fig. S2). These results suggest that SARS-CoV-2 Omicron infection induced no severe pathophysiology in the lungs but might cause brain inflammation during the later days after infection. These findings also suggest that intranasal administration of the m1E3H12 mAb could provide a protective effect against SARS-CoV-2 Omicron infection in a K18-hACE2 mouse model.
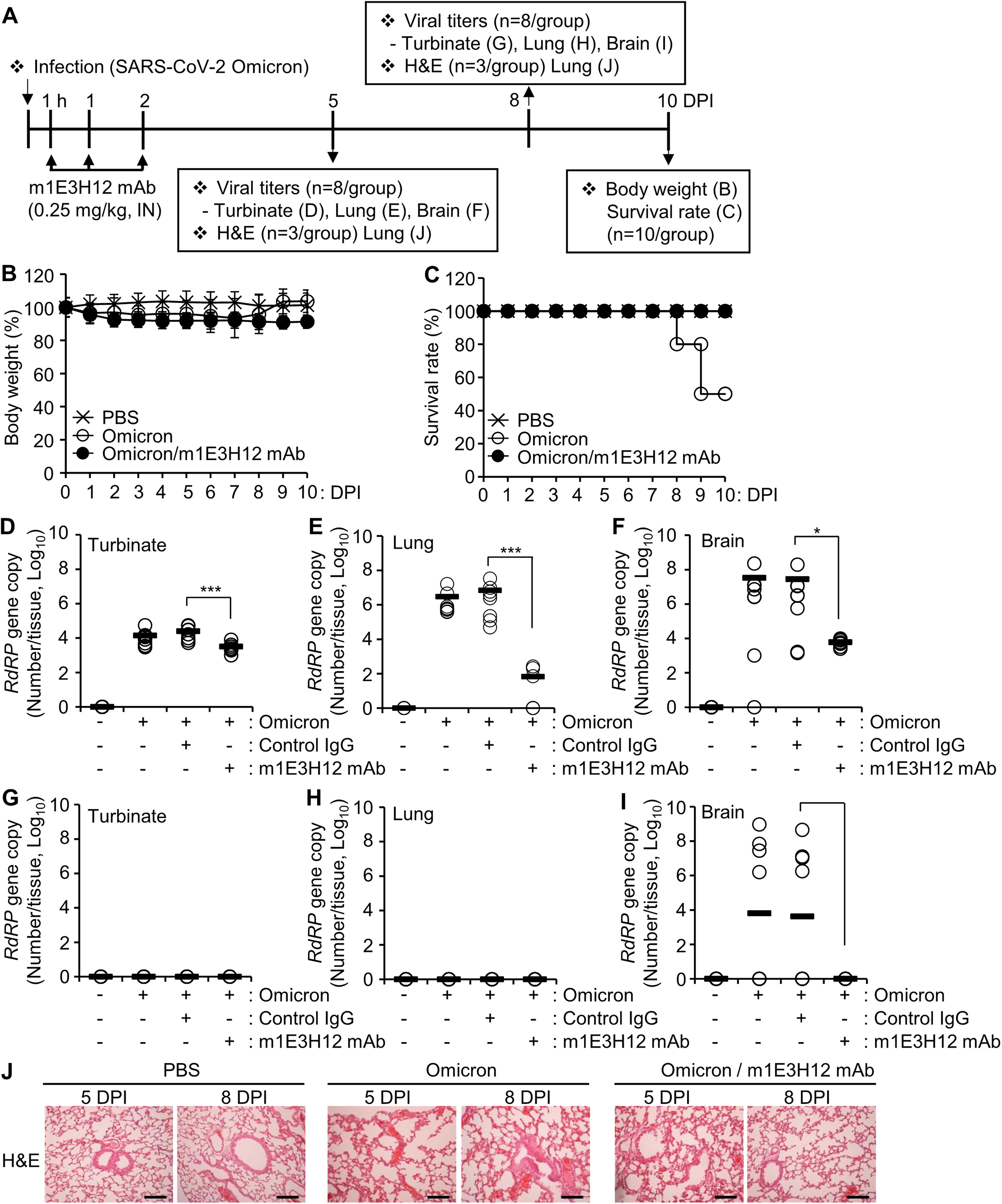
Therapeutic effect of m1E3H12 mAb against SARS-CoV-2 Omicron infection after intranasal administration in K18-hACE2 transgenic mouse model.
Efficacy of m1E3H12 mAb by intravenous and intraperitoneal treatment against SARS-CoV-2 Omicron infection in a K18-hACE2 mouse model
To further estimate the effect of administration route for m1E3H12 mAb treatment, the antibody was injected intravenously (Fig. 2) or intraperitoneally (Fig. 3) into the mice intranasally infected with SARS-CoV-2 Omicron. Importantly, we used the antibody at a 10-fold higher concentration (2.5 mg/kg) compared to the intranasal treatment. Despite this, the survival rate was only marginally reduced at 10 DPI following intravenous (Fig. 2C) or intraperitoneal (Fig. 3C) administration of the m1E3H12 mAb. At 5 DPI, viral loads in the lungs showed a significant reduction, though the efficacy was lower than that of intranasal treatment (Fig. 1E vs. Fig. 2E and Fig. 3E), while no statistically significant differences were observed in the turbinate (Fig. 2D and Fig. 3D). These results suggest that the intranasal route is the most effective for m1E3H12 mAb treatment.

No significant therapeutic effect of m1E3H12 mAb against SARS-CoV-2 Omicron infection by intravenous administration in K18-hACE2 transgenic mouse model.

No significant therapeutic effect of the m1E3H12 mAb against SARS-CoV-2 Omicron infection with intraperitoneal administration in K18-hACE2 transgenic mouse model.
Biodistribution of m1E3H12 mAb in mice based on different routes of administration
To investigate the in vivo distribution of m1E3H12 mAb, DyLight 755-labeled m1E3H12 mAb was administered to C57BL/6 mice via intranasal, intravenous, or intraperitoneal routes. Antibody localization was then examined using an in vivo imaging system (Fig. 4). At 1 h after intranasal administration, fluorescent signals were detected in the upper respiratory system, whereas no fluorescent signals were observed following intravenous and intraperitoneal administration (Fig. 4A). In dissected mice and isolated organs, intranasal administration resulted in fluorescent signals in the upper respiratory system, lungs, and stomach. In contrast, no fluorescent signals were detected in the upper respiratory system or lungs after intravenous or intraperitoneal administration (Fig. 4B and C). Further examination of vital organs revealed that, after intravenous and intraperitoneal administration, m1E3H12 mAb fluorescence was primarily localized in the liver and pancreas, respectively (Fig. 4C). These results suggest that intranasal administration effectively targets the m1E3H12 mAb to the upper respiratory system and lungs in this animal model.

Differential biodistribution of m1E3H12 mAb depending on route of administration in C57BL/6 mice. DyLight 755-labeled m1E3H12 mAb (50 µg/mouse) was intranasally, intravenously, or intraperitoneally administered into C57BL/6 mice.
Pharmacokinetics
An ELISA method was developed to quantify the residual amounts of intranasally administered m1E3H12 mAb in the turbinate, lungs, and sera as shown in Figure 5A. The standard curve analysis demonstrated linear regression was statistically validated up to 0.625 μg/mL of m1E3H12 mAb (Fig. 5B). If ELISA absorbance results fell outside the optimal range, all m1E3H12 mAb samples from tissues or sera were serially diluted to fit an optimal standard curve analysis. Analysis of time—residual amount curves in both the turbinate and lungs showed a biphasic PK profile, consisting of a rapid distribution phase followed by an extended terminal elimination phase (Fig. 5C and D). The elimination half-life (t1/2) values of the biotin-labeled m1E3H12 mAb after intranasal administration were 1.2 h in the turbinate and 0.4 h in the lungs during the initial phase. Conversely, in the terminal phase, the m1E3H12 mAb was eliminated more rapidly from the turbinate (t1/2, 26.4 h) than from the lungs (t1/2, 56.0 h). During intranasal drug delivery, drugs can directly enter the systemic bloodstream through the nasal mucosa or be cleared by the mucociliary action of ciliated epithelial cells in the turbinate (Gould et al., 2018). Typically, larger proteins like mAbs (approximately 150 kDa) have limited ability to reach systemic circulation. As expected, only trace amounts of m1E3H12 mAb (less than 0.1 μg/mL) were detected in serum after intranasal administration, indicating limited systemic exposure (Fig. 5E). The lungs also showed a significantly longer MRT of 48.6 h compared to 26.7 h for the turbinate. Therefore, the PK profiles suggest that intranasal administration of m1E3H12 mAb effectively delivers the antibody to the lungs and provides extended local retention.
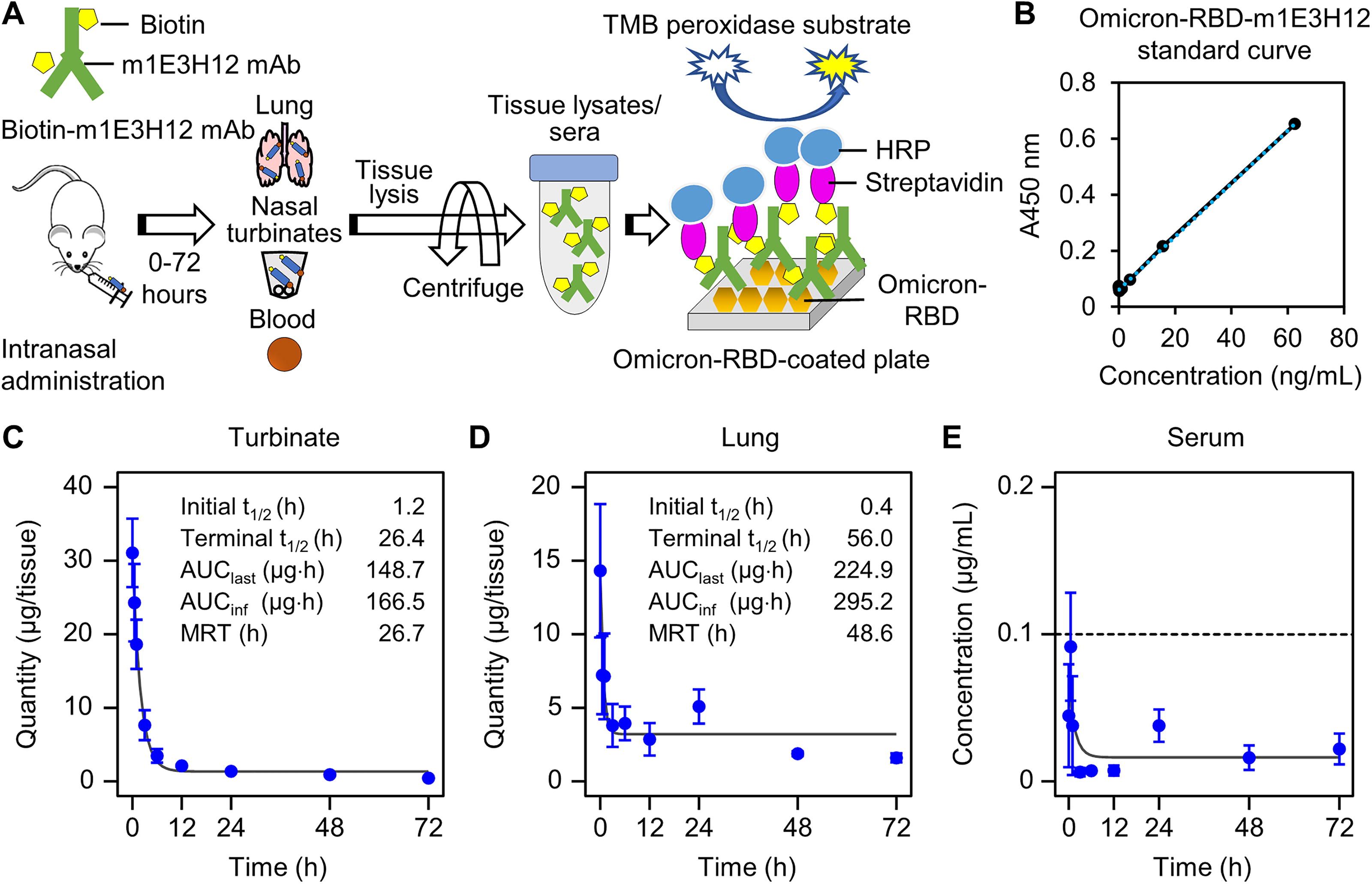
Pharmacokinetics of the m1E3H12 mAb in tissues after intranasal administration in mice.
Discussion
The route of administration of the therapeutics is a critical factor for its efficacy. The nasal drug delivery route is among the most extensively researched methods for treating respiratory infections (Parray et al., 2021). Although intravenous administration of neutralizing antibodies against SARS-CoV-2 infection is effective (Gupta et al., 2021; Weinreich et al., 2021), studies have found that intranasal treatment significantly enhances the efficacy of these antibodies during COVID-19 pathogenesis (Halwe et al., 2021; Parray et al., 2021; Duty et al., 2022; Jia et al., 2022). This is because intravenously administered antibodies cannot efficiently cross the plasma-lung barrier, necessitating large amounts of antibodies (Jia et al., 2022). Here, we compared different delivery methods to evaluate the protective effects of the recently established m1E3H12 mAb (Kim et al., 2024b) against Omicron BA.1 infection in mice.
When K18-hACE2 transgenic mice were intranasally infected with Omicron and subsequently treated with m1E3H12 mAb, intranasal administration most effectively reduced viral titers in the lungs and brains compared to intraperitoneal and intravenous treatments. In this study, we used the same antibody concentration (m1E3H12 mAb) for all routes, but the administered volumes differed: 200 μL for intravenous/intraperitoneal administration and 20 μL for the intranasal route, due to volume limitations associated with intranasal delivery, likely related to mucosal absorption capacity and safety concerns. It has been reported that administering more than 20 μL intranasally can cause suffocation and death in mice (Fox et al., 2007). Consequently, there was a discrepancy in the final dosages between the intranasal and intraperitoneal/intravenous routes. Nonetheless, even with a 10-fold lower total dose, intranasal administration of m1E3H12 mAb more effectively inhibited viral replication and enhanced survival rate compared to other routes. Therefore, we conclude that m1E3H12 mAb exerts potent therapeutic effects via the intranasal route. Its efficacy appears to be dependent on efficient delivery to the upper respiratory system and lungs. To further enhance its delivery and efficacy, we plan to optimize the formulation of m1E3H12 mAb, exploring options such as powder or solution sprays. Additionally, we acknowledge the possibility that the optimal dosage may vary depending on the administration route. Thus, conducting a dose-response experiment for intravenous/intraperitoneal routes would help determine whether higher doses via these routes could improve efficacy.
Unlike SARS-CoV-2 pathogenesis in K18-hACE2 mice infected with the parental strain or previous variants such as Alpha and Delta, Omicron infection resulted in milder clinical signs, including reduced weight loss, lethality, and less severe pulmonary damage: 75% mortality for the parental strain, 100% for Alpha and Delta, and 50% for Omicron (Natekar et al., 2022; Gonçalves et al., 2023). Consistent with these findings, we observed no significant clinical signs until 5 days post-infection (DPI), with SARS-CoV-2 Omicron causing 50% mortality by 10 DPI. Previous studies reported elevated viral loads in the lungs up to 5–7 DPI for SARS-CoV-2 Alpha and Beta infections in K18-hACE2 mice (Radvak et al., 2021). In this study, SARS-CoV-2 Omicron also induced measurable viral RNA levels in the lungs at 5 DPI, which disappeared by 8 DPI. In contrast, active viral replication in the brain was evident at both 5 and 8 DPI, accompanied by a significant increase in the chemokines RANTES (CCL5) and MIP3-Alpha (CCL20). These findings suggest that SARS-CoV-2 replication and elevated chemokine expression in the brain may contribute to the 50% mortality observed in SARS-CoV-2 Omicron-infected mice. Intranasal administration of m1E3H12 mAb significantly reduced viral replication in the brain, demonstrating its protective effect. Furthermore, expression levels of RANTES (CCL5) and MIP3-Alpha (CCL20) showed a decreasing trend following treatment, although these decreases did not reach the conventional threshold for statistical significance (p = 0.0578 and p = 0.0664, respectively). RANTES (CCL5) is known to play a role in various human disorders, including COVID-19. Targeting its receptor CCR5 with the CCR5-specific antibody leronlimab has been reported to benefit COVID-19 patients (Patterson et al., 2021; Pum et al., 2021). CCL20, which is implicated in the neuropathology of experimental autoimmune encephalomyelitis (Arima et al., 2012) and spinal cord injury (Hu et al., 2016), has also been reported to be elevated in the lungs and sera of SARS-CoV-2-infected patients (Maxwell et al., 2021; Khalil et al., 2021). Therefore, the observed decreasing trend of CCL5 and CCL20 expression following intranasal administration of m1E3H12 mAb may indicate a potential biological effect that contributes to the protection of infected mice. However, it is important to note that the K18-hACE2 transgenic mouse model was artificially engineered to overexpress human ACE2, leading to heightened susceptibility to SARS-CoV-2 and potentially more severe pathogenesis (Winkler et al., 2020). In this model, high hACE2 expression has been detected in the brain as well as the lungs, trachea, heart, stomach, intestines, kidneys, and testis. In contrast, in humans, ACE2 is primarily expressed in the lungs, stomach, intestines, kidneys, and testis (Dong et al., 2022). Moreover, we used a high viral dose and an artificial infection route, creating more extreme conditions than typical human infections. Therefore, while the K18-hACE2 model provides valuable insights, its phenotypic manifestations may not fully reflect human SARS-CoV-2 infections, particularly for Omicron, which generally causes milder disease. Caution is needed when extrapolating these findings to humans.
With some limitations, our study suggests that intranasal administration of m1E3H12 mAb could serve as an effective therapeutic intervention for SARS-CoV-2 Omicron variant infections. Previously, we developed a humanized version of the m1E3H12 mAb and demonstrated its inhibitory effects against SARS-CoV-2 Omicron variants BA.5, BQ.1.1, and XBB in vitro (Kim et al., 2024b). Further research evaluating its protective effects against Omicron subvariants and potential future variants could provide critical insights for its application in human settings.
Footnotes
Acknowledgments
The authors thank the National Culture Collection for Pathogens (Osong, Korea) for supplying SARS-CoV-2 Omicron.
Ethics Statement
The Institutional Animal Care and Use Committee of Hallym University approved the animal protocols for monoclonal antibody production and characterization and for virus challenge experiments (Permit Number: Hallym2022-46 and Hallym2023-23). All animal studies were conducted in accordance with the recommendations in the Guide for the Care and Use of Laboratory Animals of the National Veterinary Research & Quarantine Service of Korea. Research staff involved in animal care or handling took an education course for the users of the experimental animal facility in the Laboratory Animal Resources Center of Hallym University. They also took an education course for Biosafety Level 3 at the Korea Human Resource Development Institute for Health & Welfare (KOHI).
Authors’ Contributions
J.K.: Conceptualization, methodology, validation, formal analysis, writing—original draft. S.K.: Methodology, validation, formal analysis. D.K.: Conceptualization, methodology, validation, investigation, formal analysis. M.K.: Methodology, formal analysis. K.B.: Methodology, formal analysis. B.M.K.: Methodology, formal analysis. S.A.: Methodology, writing—original draft. I.G.P.: Methodology. S.K.: Methodology, formal analysis, data curation. S.P.: Methodology. J.G.S.: Methodology, formal analysis, data curation. M.N.: Methodology, formal analysis, writing—original draft. Y.L.: Conceptualization, validation, investigation, formal analysis, data curation, writing—original draft, writing—review and editing, project administration. H.-J.K.: Conceptualization, methodology, validation, investigation, formal analysis, data curation, writing—original draft, writing—review and editing, supervision, project administration, funding acquisition. All authors contributed to editing the article and approved the final article.
Author Disclosure Statement
No competing financial interests exist.
Funding Information
This research was supported by a grant from the National Research Foundation (grant number: NRF-2022M3A9I2082292, RS-2024–00401422) funded by the Ministry of Science and ICT, Republic of Korea.
Supplementary Material
Supplementary Figure S1
Supplementary Figure S2
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.