
Abstract
Objective:
Vascular endothelial growth factor (VEGF) family members are critical regulators of tissue repair and depending on their distinct pattern of receptor specificity can also promote inflammation and scarring. This study utilized a receptor-selective VEGF to examine the role of VEGF receptor (VEGFR)-2 in scar tissue (ST) formation.
Approach:
Cutaneous skin wounds were created in mice using a 4 mm biopsy punch and then treated until closure with purified VEGF-E derived from orf virus stain NZ-2. Tissue samples were harvested to measure gene expression using quantitative PCR and to observe ST formation through histological examination and changes in cell populations by immunofluorescence.
Results:
VEGFR-2-activation with VEGF-E increased expression of anti-inflammatory cytokine interleukin (IL)-10 and reduced macrophage infiltration and myofibroblast differentiation in wounded skin compared with controls. VEGF-E treatment also increased microvascular density and improved pericyte coverage of blood vessels in the healing wounds. The ST that formed following treatment with VEGF-E was reduced in size and showed improved collagen structure.
Innovation:
The role of VEGFR-2 activation in wound epithelialization and angiogenesis is well established; but its contribution to ST formation is unclear. This study tests the effect of a selective VEGFR-2 activation on ST formation following cutaneous wounding in a murine model.
Conclusion:
VEGFR-2 stimulation can enhance the quality of skin repair, at least, in part, through the induction of IL-10 expression and dampening of wound inflammation and fibrosis. Therapies that selectively activate VEGFR-2 may therefore be beneficial to treat impaired healing or to prevent excess scarring.

Introduction
T
VEGF-A is the prototype and most studied member of the VEGF family, the mammalian members of which include VEGF-B, VEGF-C, VEGF-D, and placental growth factor (PlGF). 2,3 Highly related proteins, designated VEGF-E or VEGF-F, have also been found encoded by skin viruses of the Parapoxvirus genus and isolated from Trimeresurus flavoviridis snake venom, respectively. 4,5 The VEGF family members mediate their biological functions through three VEGF receptors, VEGFR-1–3, with varying contributions from the coreceptors, neuropilin (NRP)-1, NRP-2, and heparin sulfate proteoglycans. 6,7 Each VEGF family member shows a distinct pattern of receptor specificity.
VEGF-A interacts with both VEGFR-1 and VEGFR-2 regulating multiple steps in skin repair processes. 8 –10 VEGF-A is produced by migrating keratinocytes and macrophages 11,12 and stimulates angiogenesis, the growth of new blood vessels, supplying critical nutrients and oxygen to the wound. 13 VEGF-A acts through VEGFR-2 to induce endothelial cell proliferation, migration, and survival, and promote vascular permeability. 7,11 VEGF-A-induced keratinocyte activation and matrix metalloprotease (MMP) production are also mediated by VEGFR-2. 14 –18 Consistent with these findings, animal studies have shown that reduced VEGF-A activity consistently leads to slower healing, 19,20 while augmenting VEGF-A through gene or protein delivery can accelerate healing. 14,21,22
VEGF-A has been proposed to adversely influence the quality of skin repair. 23 VEGF-A expression correlates with ST formation, 24 –26 while VEGF-A neutralization reduces scar size and improves structure of the collagen matrix. 27 Long-term exposure to VEGF-A leads to the formation of disorganized blood vessels and hyperpermeability resulting in edema, 28,29 which can detrimentally influence scar formation. 23,30 VEGF-A also promotes monocyte activation and chemotaxis through its interaction with VEGFR-1, 31 –33 which likely contributes to wound inflammation. VEGF-A also directly influences the functional activity of fibroblasts and mesenchymal stromal cells. 34,35
Although both VEGF-B and PlGF selectively bind VEGFR-1, their effects on cutaneous repair are considerably different. VEGFR-1 binding by PlGF directly induces monocyte migration and inflammatory cytokine production, 31,35 with transgenic overexpression in the skin leading to enlarged leaky vessels and increased inflammatory cell infiltration. 36 PlGF is expressed during wound healing by migrating keratinocytes and endothelial cells and PlGF-deficient mice exhibit delayed wound closure associated with reduced plasma extravasation. 37 In diabetic mice suffering from impaired healing, however, PlGF gene delivery increased monocyte recruitment and growth factor expression, indirectly resulting in enhanced wound vascularization and closure. 38 The impact of PlGF on cutaneous scarring has not been well studied, but following glaucoma filtration surgery, inhibition of PlGF reduces postoperative inflammation and collagen deposition 39 and in liver cirrhosis reduces inflammation and fibrosis. 40 VEGF-B, in contrast to PlGF, shows minimal expression in the skin. 41 Transgenic overexpression of VEGF-B also does not induce vascular permeability or inflammation, 42 potentially due to a lack of VEGFR-1 dimerization and activation. 32,43
VEGF-C and VEGF-D act via VEGFR-2 and also VEGFR-3, directly stimulating the proliferation and migration of lymphatic endothelial cells. 44,45 In skin wounds, macrophages are the major source of VEGF-C and VEGF-D, the expression of which is reduced under diabetic conditions. 46 –48 VEGF-C protein or gene delivery increased inflammation, angiogenesis, lymphangiogenesis, and closure of skin wounds, 49 –51 while specific inhibitors of VEGFR-3 resulted in lymphedema or delayed wound healing. 51,52 VEGF-C gene transfer or VEGFR-3 activation also increased the drainage function of the lymphatic system reducing ultraviolet B- and chronic contact hypersensitivity-induced skin inflammation. 53,54 Skin wounds in VEGF-D-deficient mice are highly edematous and thickened due to inadequate lymphatic drainage. 55 VEGF-C and VEGF-D also stimulate myofibroblast activation, MMP production, and collagen constriction. 56,57 The impact VEGFR-3-mediated lymphatic drainage and fibrosis may have on cutaneous scarring is unknown.
Of the nonmammalian VEGF family members, the VEGF-E but not VEGF-F variants have been shown to affect cutaneous repair. VEGF-E genes have been identified in the genomes of numerous Parapoxviruses, 4 the most studied are strains NZ-2, NZ-7, and D-1701 strains of orf virus. 58 –60 Orf virus infects wounded skin in sheep and humans, causing proliferative lesions with extensive epidermal hyperplasia and vascularization. 61,62 However, even hyperproliferative and persistent lesions reported in immunocompromised individuals resolve with minimal scarring. 63 The VEGF-E genes of the three orf virus strains share only 43–86% amino acid identity, yet are structurally similar to one another. 64,65 The purified VEGF-E proteins are specific mitogens for endothelial cells but differ in their affinity for VEGFR-2 and NRP-1 and ability to induce vascular permeability. 58 –60,65 The VEGF-ED-1701 protein induces NO release, promoting capillary-like tube formation in vitro and hyperproliferation of endothelial cells when the transgene is overexpressed in β cells of the pancreas of mice. 66,67 Transgenic overexpression of VEGF-ENZ-7 in murine skin induced angiogenesis but not lymphangiogenesis with minimal edema or inflammation, 68 while transgene delivery to skin wounds accelerated repair, with enhanced angiogenesis and less inflammatory side effects than VEGF-A delivery. 69 Administration of the VEGF-ENZ-2 protein promoted angiogenesis in ovine skin and epithelialization of murine and equine wounds. 18,65,70
The findings with the VEGF-E variants suggest that VEGFR-2 activation can promote healing of cutaneous wounds without the inflammatory side effects associated with VEGFR-1 or VEGFR-3 activation. The impact of VEGFR-2 activation on ST formation is, however, unknown. We therefore utilized the selectivity of VEGF-E to examine the role of VEGFR-2 in inflammation, fibrosis, vessel functionality, and the quality of skin repair using a murine full-thickness skin wound model. Purified VEGF-ENZ-2 protein was administered until the time of wound closure. Selective activation of VEGFR-2 with the VEGF-E variant significantly increased interleukin (IL)-10 expression, leading to decreased macrophage infiltration, myofibroblast differentiation, and ST formation, while improving capillary pericyte coverage and structure of the collagen matrix.
Clinical Problem Addressed
Impairments in healing can greatly impact quality of life, with an estimated 37 million individuals globally developing chronic wounds 71 and an estimated 100 million surgical patients in the Western world suffering from excessive scarring. 72 Chronic wounds show abnormally low levels of VEGF-A protein, which likely contributes to the impaired vascularization and delayed healing observed in wounds of different etiology. 73,74 Consistent with this, human trials have shown that augmenting VEGF-A can accelerate healing. 75 Excess VEGF-A, however, has been implicated in tissue inflammation, edema, and scar formation. 23,28,32 Given the clinical potential of VEGF-A, it is critical to define the specific activities of the VEGF family members that influence the quality of tissue repair. The findings from this study indicate that in the absence of VEGFR-1 binding, VEGFR-2 stimulation can enhance the quality of skin repair. Selective activation of VEGFR-2 may therefore be a therapeutic option for impaired healing or excess scarring.
Materials and Methods
Recombinant proteins
Recombinant FLAG-tagged VEGF-ENZ-2 and murine VEGF-A isoform164 were expressed in 293-EBNA cells, purified, and quantitated as previously described. 65
Ethics statement
Animal procedures were conducted with approval from the Animal Ethics Committee of the University of Otago (No: 67/08) with animals sourced and housed in the Hercus-Taieri Resource Unit (Dunedin, New Zealand).
Cutaneous wound model
C57BL/6 mice (female, 8 weeks of age) were anesthetized, shaved, and then two full-thickness excisional wounds were made on shaved dorsal skin using a sterile 4 mm biopsy punch (Fig. 1A). Animals were separated into three groups, control (vehicle only), VEGF-A treated, or VEGF-E treated. Two days postsurgery, each wound received 50 μL of VEGF-A (1 μg diluted in phosphate-buffered saline [PBS]), VEGF-E (1 μg diluted in PBS), or vehicle (PBS) by subcutaneous (SC) injection 0.5 cm from the lateral edge of the wound (Fig. 1B). Repeat treatments were administered on days 4, 6, and 8, at the caudal, medial, and then cranial wound edges, respectively (Fig. 1B, C). Mice were euthanized on day 3, 6, 9, or 16; wounds were surgically excised and then bisected at the center of the wound along the medial–lateral axis (Fig. 1B, D). The bisected skin was stored in RNAlater RNA Stabilization solution (Qiagen, Hilden, Germany) or fixed in 0.5% zinc salts solution, embedded in paraffin, and sectioned (4 μm).
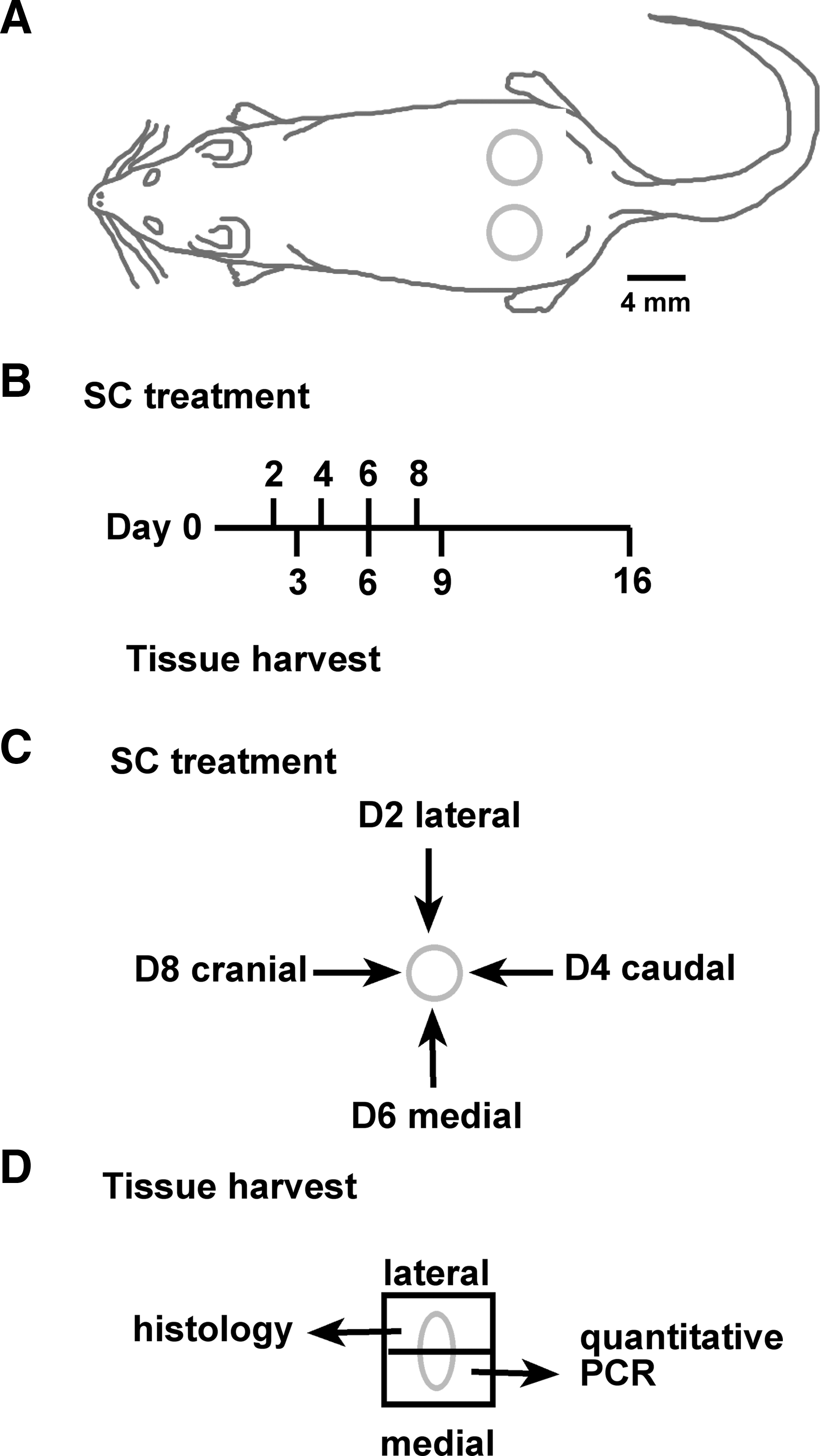
Design of experimental wounding, treatment, and tissue harvesting procedures.
Quantitative RT-PCR
Four treatment-matched wounds were combined per sample with total RNA extracted using TRIzol (Invitrogen, Carlsbad, CA) and Proteinase K (Sigma-Aldrich, St. Louis, MO) digestion and then purified using the RNeasy® Mini Kit (Qiagen). 76 Total RNA was reverse transcribed using Superscript III (Invitrogen), oligo(dT)15, and random hexamer primers. Real-time quantitative PCR was conducted with gene-specific primers (glyceraldehyde 3-phosphate dehydrogenase [GAPDH] 5′CAAAAGGGTCATCATCTCCG 3′ and 5′TAAGCAGTTGGTGGTGCAGGA 3′; CXCL2 5′CATCCAGAGCTTGAGTGTGACG 3′ and 5′GCTTCAGGGTCAAGGCAAACTT 3′; IL-1β 5′CTTCCAGGATGAGGACATGAGC 3′ and 5′AGTGCAGTTGTCTAATGGGAACG 3′; transforming growth factor [TGF]-β1 5′ GCTTGCAGAGATTAAAATCAAGTG 3′ and 5′ACCAAGGTAACGCCAGGAATTG 3′; VEGF-A 5′ GCAGGCTGCTGTAACGATGAAG 3′ and 5′GCTTTGGTGAGGTTTGATCCG 3′; platelet-derived growth factor [PDGF]-β 5′ CGGAGTCGAGTTGGAAAGCTC 3′ and 5′ATCTGGAACACCTCTGTGCG 3′; IL-10 5′CTAGAGCTGCGGACTGCCTTC 3′ and 5′AGGAGTCGGTTAGCAGTATGTTG 3′) and SYBR® Green PCR Master Mix (Invitrogen) using the ABI PRISM 7700 Sequence Detection System (Applied Biosystems, Foster City, CA). All mRNA levels were quantitated relative to GAPDH and unwounded skin taken from age- and sex-matched mice. 76
Immunohistochemistry
Paraffin-embedded skin sections (4 μm) were incubated with antibodies to F4/80 (AlexaFluor®488-BM8, No. 14-4801, dilution 1:100; eBioscience, San Diego, CA), alpha smooth muscle actin (αSMA, Cy3-1A4, No. C6198, dilution 1:400; Sigma-Aldrich), vimentin (AlexaFluor488-D21H3, No. 5741, dilution 1:100; Cell Signaling Technology, Danvers, MA), or von Willebrand factor (vWF, No. A 0082, dilution 1:200; DakoCytomation, Glostrup, Denmark) followed by incubation with AlexaFluor 488-conjugated antibody (No. A-11008, 1:500 dilution; Invitrogen). Nuclei were stained with 4′,6-diamidino-2-phenylindole (DAPI, No. D3571, 75 nM; Invitrogen) and slides mounted with SlowFade Gold (Invitrogen). Images were taken of the stained section at 40 × magnification and then converted into panoramas using Photoshop (Adobe Systems, San Jose, CA).
The number of macrophages in the wound edge and bed was quantitated in images taken of F4/80-stained sections from skin harvested 6 days postwounding. 76 Nucleated green-stained cells were counted manually to a point 500 μm distal to the cut edge of the panniculus carnosus. The corresponding region was outlined using the freehand tool in ImageJ, * the outlined area measured, and then cell number expressed relative to that area.
Myofibroblasts in the GT were quantitated in images taken of vimentin and αSMA-stained sections from skin harvested 6 days post wounding. 76 Images were converted to RGB stacks and then the GT outlined as described above. The threshold in the red channel was adjusted automatically to highlight the stained cells, and then measurements taken as a percentage of total GT area.
Blood vessels in the GT were quantitated in images taken of vWF- and αSMA-stained sections from skin harvested 9 days postwounding. 76 The GT was outlined as described above, and then, the number of blood vessels lined with green-stained endothelial cells and red-stained pericytes counted manually. The corresponding region was measured, and vessel number expressed relative to that area. 70
Histology
Paraffin-embedded skin sections (4 μm) were stained with Martius scarlet blue (MSB) and then visualized under bright light, or with picrosirius red visualized under dark field with a polarized filter. 77 Images were taken of the section at 10 × magnification and then converted into panoramas using Photoshop.
Using ImageJ, epidermal and dermal thickness measurements were obtained from images of MSB-stained sections, from skin harvested 16 days postwounding. Lines were drawn perpendicular to the epidermal–dermal demarcations and four epidermal and dermal thickness measurements were taken for each section—two within the cut edge of the panniculus carnosus, and two at a distance of 500 μm on either side of the wound. 78 Fibrotic ST extending to the depth of the panniculus carnosus was outlined in images of the MSB-stained sections using the freeform tool of ImageJ. A measurement was taken in square micrometers. The fibrotic scar size was then determined by the dermal scar index, 78 which was calculated by dividing the scar area (μm2) by the average dermal thickness (μm). For the epidermis, scar size was determined by the epidermal scar index, which was calculated by dividing the average of the internal epidermis thickness measurements (μm) by the average of the distal epidermis thickness measurements (μm).
Collagen density within the ST was examined in images of the picrosirius red-stained sections from skin harvested 16 days postwounding following conversion to 8 bit. ST was outlined as described above, the threshold adjusted automatically to highlight the stained collagen, and then, measurements taken as a percentage of the total ST area.
Collagen complexity was examined in images of the ST in picrosirius red-stained sections from skin harvested 16 days postwounding taken at 40 × magnification. Fast Fourier transform (FFT) statistical analysis was conducted on the images using ImageJ. The collagen orientation index was determined by calculating the width/length ratio of the zeroth-order maximum in the generated plot of the image. 77 Normal skin contains collagen bundles in a basket weave-like pattern that appear randomly organized, which results in a circular power spectrum and an orientation index approaching 1. In contrast, the collagen bundles in mature ST tend to be parallel to the epidermis, leading to an elongated power spectrum and a small orientation index.
Statistical analyses
Values are expressed as the mean ± standard error of the mean (SEM). The normality of each data set was confirmed using the Shapiro–Wilk normality test. One-way analysis of variance (ANOVA) was then conducted with significant points of difference between means determined using Sidak's test following correction for multiple comparisons. Values of p ≤ 0.05 were considered statistically significant.
Results
VEGF-E alters regulatory gene expression in cutaneous wounds
To evaluate the impact of selective VEGFR-2 activation on cutaneous wound healing, punch biopsy excisional wounds were treated with VEGF-A or VEGF-E by SC injection 2, 4, 6, and 8 days postinjury. Wounds were surgically excised on day 3, 6, 9, or 16 and specific transcript levels examine by quantitative RT-PCR. Administration of VEGF-A significantly increased mRNA levels for the inflammatory mediators CXCL2 and IL-1β (Fig. 2A, B) at day 3, but levels were comparable to vehicle-treated controls at the other time points examined. The reparative growth factors TGF-β1 and VEGF-A were increased significantly in wounds treated with VEGF-A at days 3, 6, and 9 compared to vehicle-treated controls (Fig. 2C, D). VEGF-E treatment also increased growth factor expression (TGF-β1: day 3–16, VEGF-A: day 3 and 9) relative to control wounds (Fig. 2C, D), but did not significantly alter the expression of inflammatory mediators (Fig. 2A, B). Compared to control, both VEGF-A and VEGF-E induced an early (day 3), and in the case of VEGF-E prolonged (day 9), wound site expression of an additional growth factor PDGF-β (Fig. 2E). VEGF-E also significantly increased mRNA expression of the anti-inflammatory cytokine IL-10, over that of VEGF-A- and control-treated wounds (day 3, Fig. 2F).

VEGF-E alters regulatory gene expression in cutaneous wounds. Expression of
VEGF-E reduces the macrophage and myofibroblast response to cutaneous wounding
Six days postwounding, following two treatments with the VEGF proteins, the infiltrate within the wound skin was examined. A substantial influx of F4/80+ve macrophages was observed at the margin of vehicle-treated control wounds (Fig. 3A, B). The number of macrophages in wound bed (WB) and wound margin following VEGF-A treatment was equivalent to controls, while there was a significant (more than twofold) decrease in macrophage number in VEGF-E-treated skin (Fig. 3C).

VEGF-E reduces the macrophage and myofibroblast response to cutaneous wounding.
An accumulation of vimentin+ve fibroblasts was also evident in the newly formed GT of vehicle-treated wounds, with overlapping staining for αSMA, which is consistent with their differentiation into myofibroblasts (Fig. 3D, E). No differences in vimentin staining were observed between the treatment groups (not shown). In VEGF-A-treated wounds, the area of αSMA staining within the GT was, however, significantly reduced relative to control wounds (twofold, Fig. 3F). Treatment with VEGF-E also resulted in a significant sixfold decrease in αSMA staining compared to vehicle-treated wounds (Fig. 3F).
VEGF-E enhances vascularization of cutaneous wounds
Nine days postwounding, following four VEGF protein treatments, vascularization within the GT was examined. The GT had now filled the WB, and contained both individual vWF+ve endothelial cells and mature blood vessels lined with vWF+ve endothelial cells and surrounded by αSMA+ve cells consistent with pericytes (Fig. 4A, B). Both VEGF-A- and VEGF-E-treated wounds showed an increase in the number of endothelial cells and mature blood vessels within the GT compared to control wounds (Fig. 4C, D). The number of mature blood vessels was, however, greater in wounds treated with VEGF-E than those treated with VEGF-A (Fig. 4D). VEGF-E-treated wounds also exhibited less extravascular αSMA+ve cells, with a significant reduction in staining compared to VEGF-A- and vehicle-treated wounds (Fig. 4E).
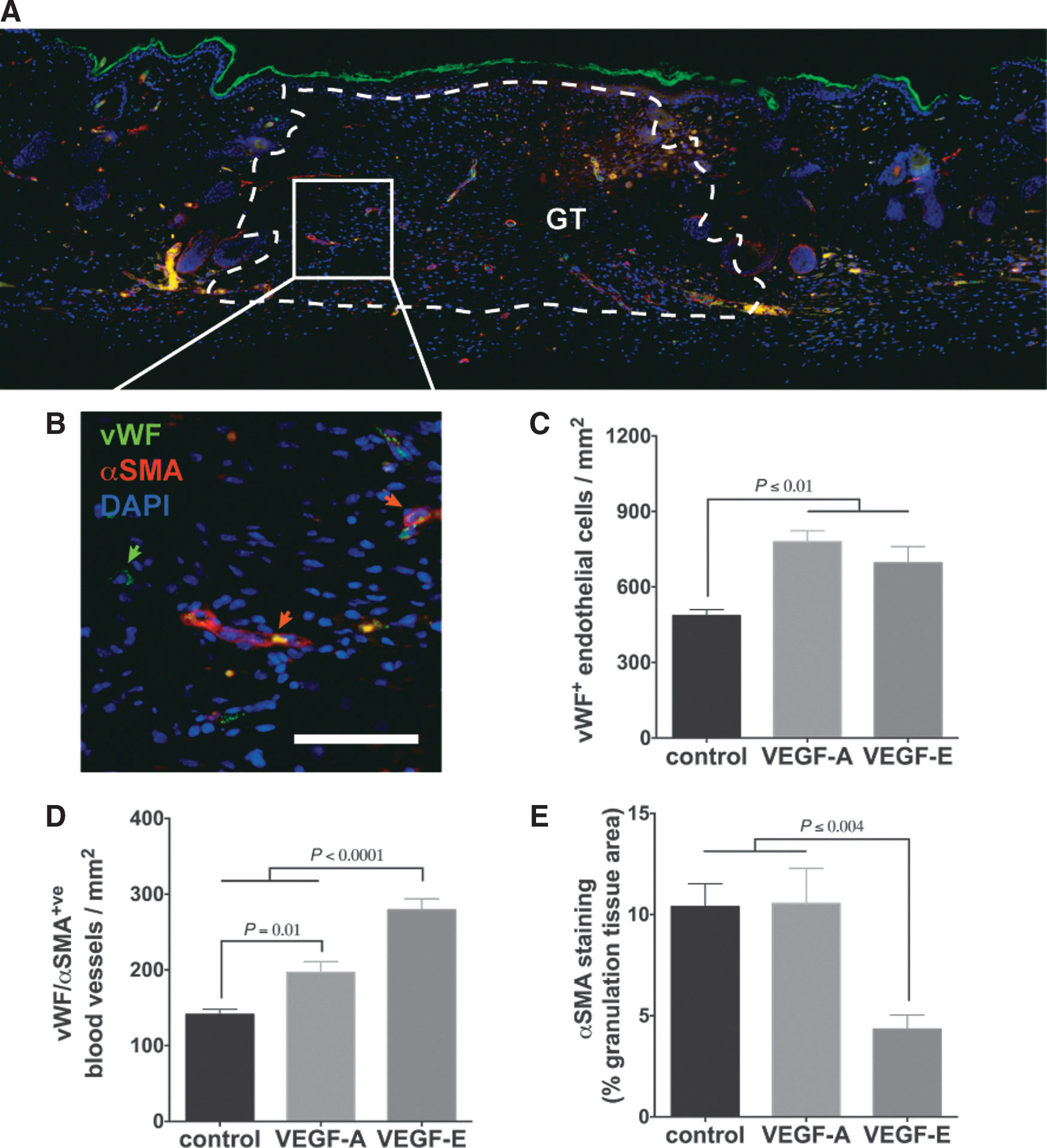
VEGF-E enhances vascularization of cutaneous wounds.
VEGF-E reduces the size and improves the quality of scars formed following cutaneous wounding
Sixteen days postwounding, 8 days after the final VEGF treatment, the size and quality of the newly formed scar were examined. A narrow scar was now evident with a thickened epidermal layer and fibrotic tissue extending from the epidermal–dermal junction to the depth of the panniculus carnosus (Fig. 5A). The epidermal scar in vehicle- and VEGF-A-treated wounds was twice as thick as the epidermis distal to the wound (Fig. 5A, B). In wounds treated with VEGF-E, however, the epidermal scar was significantly thinner than VEGF-A-treated and control wounds and closer in thickness to that of the normal epidermis (Fig. 5A, B). Wounds treated with VEGF-A also resulted in a dermal scar that was ∼50% larger than both the vehicle- and VEGF-E-treated wounds (Fig. 5A, C). The dermal scars in VEGF-E-treated wounds were, however, significantly smaller than those of control wounds (Fig. 5A, C).

VEGF-E reduces the size and improves the quality of scars formed following cutaneous wounding.
Collagen deposition and bundle formation were evident within the fibrotic scar particularly in the upper dermis (Fig. 5D, E). Both VEGF-A- and VEGF-E-treated wounds showed a significant increase in the amount of collagen staining within the dermal scar area compared with control wounds (Fig. 5F). Collagen structure within the ST was then examined using FFT analysis (Fig. 5G). The collagen bundles in the scars from vehicle- and VEGF-A-treated wounds showed a reduced FFT aspect ratio compared with unwounded skin, indicating the bundles were more parallel in orientation (Fig. 5H). Following VEGF-E treatment, the FFT aspect ratio was significantly greater than that of control scars and was approaching that of the random basket weave pattern found in unwounded skin (Fig. 5H).
Discussion
VEGF family members, acting through three distinct VEGFRs, contribute to both skin repair and scarring. VEGF-A, VEGF-C, and VEGFR-2-selective VEGF-E directly enhance wound epithelialization, angiogenesis, and the rate of closure. 13,18,22,51,69 Through the activation of VEGFR-1, PlGF and VEGF-A likely also promote wound inflammation and edema. 32,36 Activation of VEGFR-3, by VEGF-C and VEGF-D, mediates lymphatic repair, which influences wound inflammation and drainage. 51,55 This suggests that VEGFR-2 activation promotes healing of cutaneous wounds, while VEGFR-1 and VEGFR-3 activation may lead to inflammation and fibrosis. Little, however, is known as to whether VEGFR-2 stimulation impacts these processes involved in ST formation.
Macrophages are beneficial to skin repair but under certain circumstances may promote excessive inflammation or fibrosis. 79 In this study, application of the VEGF-E protein to murine skin had no effect on inflammatory gene expression but led to a reduction in macrophage infiltration of the wound. This observation was, in part, consistent with previous studies that demonstrated VEGF-E cannot induce monocyte chemotaxis in vitro or recruitment of macrophages to inflamed skin in vivo. 68,69,80 On the contrary, VEGF-A did increase expression of inflammatory genes in the wound as previously shown, 69,80 –82 but in contrast did not increase macrophage recruitment. These findings confirm a role for VEGFR-1 in inflammatory signaling but suggest that VEGFR-2 stimulation, under certain conditions, may suppress or delay macrophage recruitment, alter their differentiation or activation, or accelerate their clearance from skin. Monocyte chemotaxis can be inhibited by hypoxia or TNF treatment through the upregulation of MAPK phosphatase 1, 83 which, in endothelial cells, is directly activated by VEGFs. 84 VEGF expression and VEGFR signaling also influence the balance of classically activated proinflammatory (M1) and alternatively activated anti-inflammatory (M2) macrophage phenotypes. 85,86 In addition, VEGF-A has been shown to promote macrophage phagocytosis and apoptosis. 87,88 Myeloid-derived VEGF-A has also been implicated in inflammation resolution, through expansion of lymphatic vessels and mobilization of macrophages from inflamed skin to draining lymph nodes. 48 The next step will be to determine which if any of these mechanisms play a role in the anti-inflammatory effects of VEGFR-2.
Myofibroblast contractility promotes wound contraction but its persistence and continued activity are associated with excess scarring. 89 Following VEGF-E treatment, a sustained reduction in myofibroblasts was observed within the newly developing GT. Myofibroblast accumulation appeared delayed in VEGF-A-treated wounds, as levels were initially reduced but reached that of controls once the wounds were closed. VEGF-A directly influences myofibroblast proliferation and migration, 34,90 which suggests these functions may be induced via VEGFR-1. In the wound environment, however, activation of VEGFR-2 may, at least initially, counteract myofibroblast differentiation. A critical source of myofibroblasts is pericytes that surround the vasculature. 91 In VEGF-E-treated wounds, a stabilized vasculature with greater pericyte coverage may have limited myofibroblast differentiation. Further analysis of specific pericyte markers is needed to delineate the effects of VEGFR-2 activation on the different fibroblast and pericyte populations. Consistent with these observations, studies have demonstrated that angiogenesis driven by myeloid-derived VEGF-A limits fibrotic damage in the lung, 92 and that while VEGF-A contributes to liver fibrosis, the development of functional vessels is also critical to its resolution. 93 It will be important to determine if fibroblast function is directly altered by VEGFR-2 activation or if the effects are mediated via changes in the vasculature.
The microvasculature plays an important role in skin repair but excess capillary density and impaired functionality adversely affect ST formation. 23,94 Consistent with previous reports, 14,69,75 VEGF-A and VEGF-E administration increased microvascular density in the healed wound. Treatment with the VEGF proteins also enhanced pericyte coverage, to the greatest extent in VEGF-E-treated wounds. This was in contrast to a transgenic overexpression of VEGF-E, which resulted in poor pericyte coverage and edema in pancreatic β cells, 67 which suggests that the beneficial effects on the vasculature may be tissue, dose, or timing specific. Pericytes surround the newly formed vessels, inhibiting endothelial cell proliferation and promoting their differentiation, thereby leading to vessel stability. 95 The findings from this study suggest that functionality of the vasculature may depend on the extent of VEGFR-2 stimulation and that VEGFR-1 activity may compromise or delay this process. Consistent with this role for VEGFR-1, its conditional depletion in macrophages enhances wound angiogenesis and accelerates repair. 96 For future therapeutic benefit, it will be critical to ascertain the threshold of VEGFR-2 activation and capillary density required to maintain functionality.
VEGFs promote skin repair through accelerated wound epithelialization and GT formation, but excess VEGF-A can lead to epidermal hyperplasia 18,97 and has been implicated in scar formation. 25,98,99 In scars that formed following VEGF-E treatment, the epidermal thickness was closer to that of normal skin and the dermal scar much reduced compared to that of controls. VEGF-A treatment, however, led to a larger dermal scar than controls, which was consistent with a study that showed VEGF-A administration increased ST formation in fetal wounds, while its neutralization reduced scar size and improved collagen fibril distribution. 27 These findings implicate VEGFR-1 in mediating VEGF-A-induced dermal scarring. Both VEGFR-2 ligands increased collagen density in support of the observation that VEGFR-2 inhibition reduces fibrosis following wounding. 100 Collagen basket weave structure was also closest to that of normal skin following VEGF-E treatment, which suggests VEGFR-2 may directly or indirectly promote the transition from proliferation to remodeling. Continued investigation is, however, needed to define the long-term impact of VEGFR-2, and conversely VEGFR-1, activation on scar remodeling.
• Selective activation of VEGFR-2 with viral-derived VEGF-E increased IL-10 expression and reduced macrophage infiltration and myofibroblast differentiation in murine skin wounds.
• VEGF-E treatment increased microvascular density and improved pericyte coverage of blood vessels in healing wounds.
• Scar tissue formation was reduced following treatment with VEGF-E and showed improvements in collagen density and orientation.
Current approaches aimed at treating fibrosis and scarring are in the most part directed at inhibiting cytokines, chemokines, MMPs, and angiogenic and profibrotic growth factors. 35 VEGF-A treatment of wounds led to increased dermal scarring, and an initial increase in CXCL2 and IL-1β mRNA expression was observed. The wounds treated with VEGF-A also showed increased TGF-β and VEGF-A mRNA expression and more sustained PDGF-β than controls, but similar profiles were evident in VEGF-E-treated wounds, which led to reduced ST formation. The primary difference in gene expression between the two treatments was the relative increase of IL-10 in VEGF-E-treated wounds. These findings suggest that VEGFR-1 and VEGFR-2 differentially regulate pro- and anti-inflammatory gene expression in the wound and that VEGFR-2-mediated effects on scarring processes may be mediated via IL-10. IL-10 improves the quality of repaired tissue by limiting inflammation and fibrosis. 101 IL-10-deficient wounds show a pronounced inflammatory cell infiltrate and adult-like scarring, 102 while gene or protein delivery of IL-10 reduces inflammation and restores the normal dermal architecture. 76,103,104 When applied to human incisional wounds, exogenous IL-10 improved the appearance and reduced the redness of scars. 105 The link between VEGFR-2, IL-10, and the quality of skin repair requires further corroboration, but scarless healing in the absence of commensal microbiota is associated with high levels of IL-10 and angiogenesis, 106 and VEGFR-2-selective VEGF-E has been shown to induce IL-10 mRNA expression in dermal fibroblasts. 107
In conclusion, VEGFR-2 stimulation may enhance not only the rate but also the quality of cutaneous repair, at least in part, through the induction of IL-10 expression and dampening of wound inflammation and fibrosis. In contrast, VEGF-A-induced VEGFR-1 activation adversely influences inflammatory and scarring processes in the skin. These findings suggest that therapies selectively activating VEGFR-2 may show greater benefit as treatments for impaired healing or for preventing excess scarring.
Innovation
VEGF family members are critical regulators of tissue repair and depending on their distinct pattern of receptor specificity can promote wound repair or inflammation, fibrosis, and scarring. Functional studies with VEGFR-selective variants of the VEGFs could shed light on the impact of the individual VEGFRs during cutaneous wound repair. The principal innovation of this study was the treatment of murine wounds with viral-derived VEGF-E to assess the role of VEGFR-2 in ST formation. The findings have clinical significance in that they may lead to improved angiogenic therapies for wound complications.
Footnotes
Acknowledgments and Funding Sources
This research was supported by a contract from the Health Research Council of New Zealand (No. 13/774). We are grateful to Amanda Fisher, Janine Neill, and Alisha Shaw from the Histology Unit, Department of Pathology, University of Otago, for expert technical assistance.
Author Disclosure and Ghostwriting
The authors do not have any commercial or financial conflicts of interests to declare. The named authors wrote this article and no ghostwriters were used.
About the Authors