
Abstract
Objective:
In early gestation, fetal wounds heal without fibrosis in a process resembling regeneration. Elucidating this remarkable mechanism can result in tremendous benefits to prevent scarring. Fetal mouse cutaneous wounds before embryonic day (E)18 heal without scar. Herein, we analyze expression profiles of fetal and postnatal wounds utilizing updated gene annotations and pathway analysis to further delineate between repair and regeneration.
Approach:
Dorsal wounds from time-dated pregnant BALB/c mouse fetuses and adult mice at various time points were collected. Total RNA was isolated and microarray analysis was performed using chips with 42,000 genes. Significance analysis of microarrays was utilized to select genes with >2-fold expression differences with a false discovery rate of <2. Enrichment analysis was performed on significant genes to identify differentially expressed pathways.
Results:
Our analysis identified 471 differentially expressed genes in fetal versus adult wounds following injury. Utilizing enrichment analysis of significant genes, we identified the top 20 signaling pathways that were upregulated and downregulated at 1 and 12 h after injury. At 24 h after injury, we discovered 18 signaling pathways upregulated in adult wounds and 11 pathways upregulated in fetal wounds.
Innovation:
These novel target genes and pathways may reveal repair mechanisms of the early fetus that promote regeneration over fibrosis.
Conclusion:
Our microarray analysis recognizes hundreds of possible genes as candidates for regulators of scarless versus scarring wound repair. Enrichment analysis reveals 109 signaling pathways related to fetal scarless wound healing.

Introduction
A
Clinical Problem Addressed
Adult cutaneous wounds heal through a fibroproliferative response that results in incomplete regeneration of the original tissue. 7 There is an overproduction of an unorganized collagen meshwork with loss of dermal appendages. 8 The resultant scar tissue is weaker and has a tensile strength <80% of the original form. 9 This highly evolved process, although efficient in protecting from infection, further injury, and water loss, can be problematic. In the pediatric population, scars can restrict growth and impede movement when they occur across joints. Furthermore, scars occurring in locations such as the face can lead to devastating psychological and social consequences. In addition, humans can develop pathological scars, such as keloids or hypertrophic scars, when wound healing occurs rampantly. This results in a scar that is characterized by excessive disorganized collagen deposition and is raised, cosmetically unpleasing, and often associated with symptoms such as pain and itching. 10 The ability to prevent such scar formation and promote wounds to undergo regeneration would be highly beneficial to healthcare and society, by easing the tremendous clinical burden of scarring and fibrosis estimated to be in the tens of billions of dollars. 9
Materials and Methods
Animals
Six-week-old wild-type BALB/c mice were purchased from Charles River Laboratories (Wilmington, MA) and housed at 22°C with a 12-h day/12-h night cycle. All animals received water and normal chow ad libitum. For timed gestations, the mice were bred overnight and the day of vaginal plug was considered E0.5 day of gestation. Animals were maintained in the Stanford Animal Care Laboratory and all procedures were conducted in accordance with university-approved protocols according to National Institutes of Health (NIH) guidelines.
Fetal mouse wounding
Fetal wounding was performed as previously described. 11 Briefly, pregnant mice (gestational age E17) were utilized. After induction of anesthesia, midline laparotomy was performed using microsurgical scissors. The uterus and fetus selected for surgery were gently exposed using cotton-tip applicators. The surgical field was irrigated with warm (38°C) phosphate-buffered saline (PBS). The fetus was carefully positioned to allow access to the dorsum. A purse-string stitch using a 7-0 nylon suture was passed through the uterus overlying the site of intended dorsal wounding. A 3 mm incision was made through the uterine wall and amniotic sac in the center of the purse-string stitch. Using microsurgical scissors, a single full-thickness excisional wound, ∼1 mm in diameter, was created on the dorsum of the fetus. The wound was marked with India ink. Warm (38°C) PBS was injected into the amniotic sac with a blunt-tipped 10-guage syringe as the purse string was closed and the syringe was carefully retracted. One fetus of unknown gender per litter was wounded. The peritoneal cavity was irrigated with warm (38°C) PBS. The peritoneum and abdominal skin were everted, reapproximated, and stapled closed. Prior studies from our laboratory have shown that the histology of these wounds reveal complete regeneration within 48 h. Hematoxylin and eosin and trichrome staining reveal rapid reepithelialization and normal collagen architecture. 12 Thus, time points less than 48 h were chosen for analysis of wounds. The pregnant mice were sacrificed and fetal wounds were harvested at 1, 12, and 24 h (n = 3 for each time point) following wounding by excising the wound with a 2 mm rim of surrounding tissue using microsurgical scissors, as previously described. 6 An equivalent amount of tissue from the contralateral dorsum of the same fetus was collected in the same manner for normalization.
Adult mouse wounding
For adult wounding, 2 mm excisional wounds were generated on the back of 3-week-old BALB/c mice of mixed gender using punch biopsy after induction of anesthesia and preparation for aseptic surgery, as described above for pregnant mice. Mice were sacrificed and wounds were harvested at 1, 12, and 24 h (n = 3 for each time point) following injury for comparison to fetal wounds. Wounds were harvested by excising the wound along with a 2 mm rim of normal tissue by punch biopsy for reproducibility, as previously described. 6 An equivalent amount of tissue from the contralateral dorsum of the same fetus was collected in the same manner for normalization. The larger size wound, which is a fourfold increase in area, accounts for an approximately fourfold size discrepancy between the fetal and postnatal wound. However, despite the differences in wound size, the relative wound to body size in the fetal and postnatal mouse is not the same and may confound our data.
RNA extraction and amplification
RNA from fetal and adult wounds at various time points was extracted using the Trizol protocol (Invitrogen, Carlsbad, CA) as per manufacturer's instructions and as previously described. 13 One microgram of RNA from each experimental sample (n = 3 per group per time point) was amplified. One-microgram aliquots of universal mouse RNA were amplified in individual reaction mixtures and utilized as internal amplification controls.
Preparation of fluorescent complementary DNA probes
Fluorescent complementary DNA (cDNA) probes were prepared as previously described. 13
Pretreatment of microarray chips
The Stanford Microarray Database Center was used to print mouse microarray chips with 42,000 specific cDNAs printed onto each lysine-coated slide. These cDNAs represent single accession numbers from Genbank (Sequences and accession numbers of the cDNAs can be found on
Microarray hybridization
Microarray hybridization was performed as previously described. 13
Microarray data analysis
Scanned images were analyzed using the Genepix Pro 4.0 software (Molecular Devices), as previously described. 13 Significance analysis of microarrays (SAM) was used to select genes with significant expression differences between the E17 fetal and adult wound transcriptomes for each time point. Genes that had at least a twofold expression difference with false discovery rate <2 were selected.
Functional analysis of differentially expressed genes
To identify functional connections among significantly regulated genes, both network and pathway analyses of the probes filtered by microarray were performed using Ingenuity Pathways Analysis (Ingenuity Systems, Redwood City, CA), as previously described. 13
Results
Differential gene expression between adult and fetal wounds
Wound microarray data were normalized to age-matched unwounded control skin (taken from the contralateral dorsum of the same fetus/adult during wound collection) data sets. Normalized transcriptomes from E17 fetal wounds were directly compared to normalized transcriptomes from adult wounds. SAM identified 471 differentially expressed genes with greater than twofold difference between E17 fetal and adult wounds at 1, 12, and 24 h postinjury. At 1 h following wounding, 178 genes were upregulated in E17 fetal wounds, whereas 13 genes were downregulated when compared to adult wounds (Table 1). At 12 h following injury, E17 fetal wounds upregulated 112 genes, whereas adult wounds upregulated 141 genes (Table 2). Twenty-four hours postwounding, 16 genes were downregulated in E17 fetal wounds and 11 genes were upregulated versus adult wounds (Table 3).
Differentially expressed genes in embryonic day 17 fetal versus adult wounds at one hour postwounding
Differentially expressed genes in embryonic day 17 fetal versus adult wounds at twelve hours postwounding
tRNA, transfer RNA.
Differentially expressed genes in E17 fetal versus adult wounds at twenty-four hours postwounding
E, embryonic day.
Functional pathway analysis
Analysis of the 178 genes found to be upregulated in E17 fetal wounds at 1 h postinjury resulted in the identification of 20 functional pathways (Fig. 1B). The top five pathways were associated with the following: proline biosynthesis, arginine degradation, mevalonate pathway, and Parkinson's signaling. Utilizing the 13 downregulated genes in E17 fetal wounds, a list of 20 functional pathways were again identified (Fig. 1C). The top five pathways were as follows: interleukin (IL)-8 signaling, mechanistic target of rapamycin signaling, glucocorticoid receptor signaling, regulation of IL-2 expression in activated and anergic T lymphocytes, and T cell receptor signaling.

Microarray analysis of E17 fetal and adult wounds at 1 h postwounding.
At 12 h following injury, out of 112 genes upregulated in E17 fetal wounds, 20 pathways were identified (Fig. 2B). The top five pathways were related to apoptosis, role of osteoblasts, osteoclasts and chondrocytes in rheumatoid arthritis, Ras-related nuclear protein signaling, and protein ubiquitination pathway. Conversely, of 141 genes upregulated in adult wounds at 12 h, 20 functional pathways (Fig. 2C) reveal the top five to include the thioredoxin pathway, CXC chemokine receptor (CXCR)4 signaling, IL-1 signaling, Cdc42 signaling, and vitamin C transport.
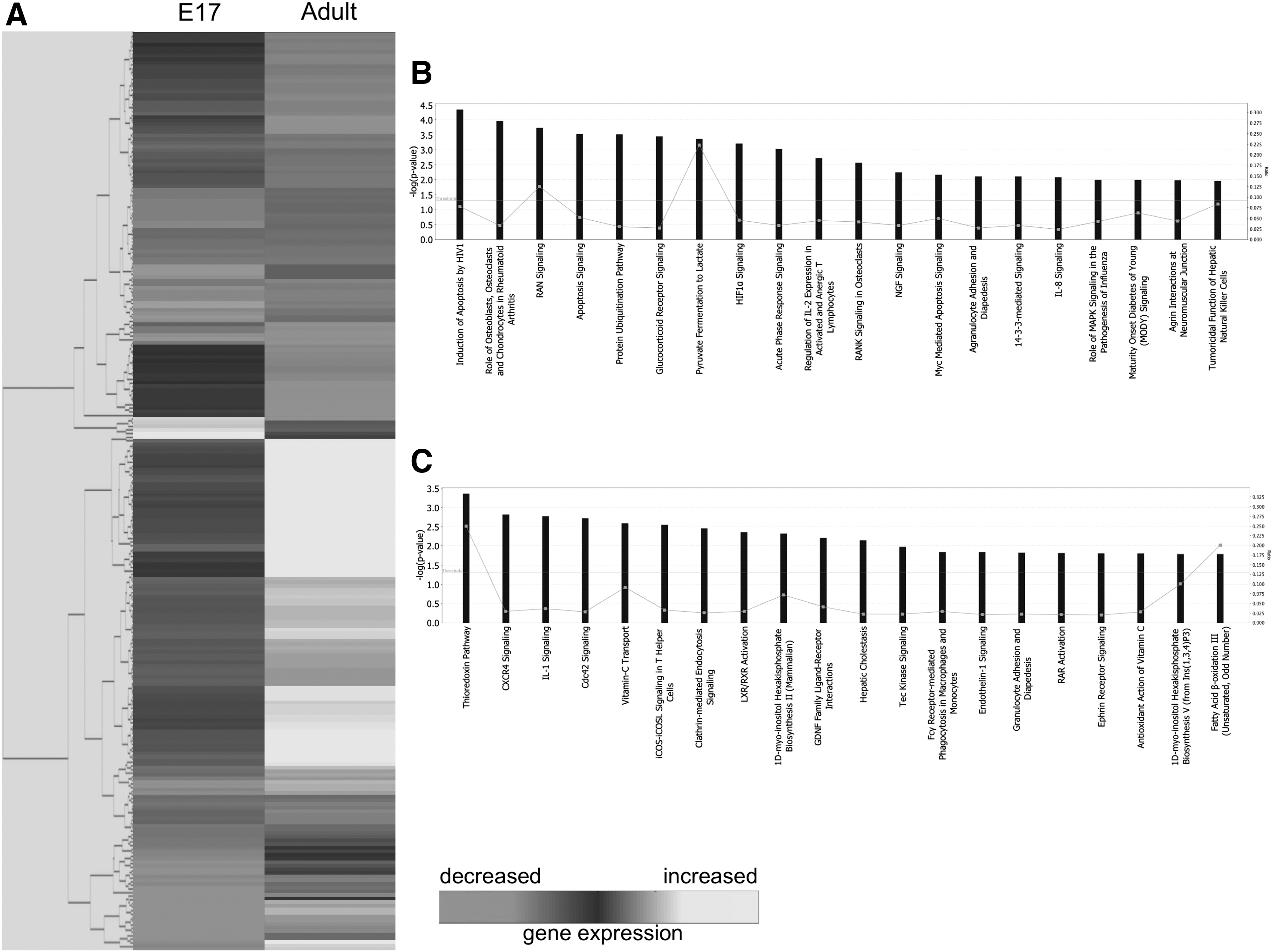
Microarray analysis of E17 fetal and adult wounds at 12 h postwounding.
Gene expression of E17 fetal and adult wounds at 24 h after injury reveal 27 differentially expressed genes. Of the 16 downregulated in E17 fetal wounds, 18 pathways were distinguished (Fig. 3B). Eumelanin biosynthesis, transfer RNA (tRNA) charging, glial cell line-derived neurotrophic factor family ligand–receptor interactions, neurotrophin/tropomyosin receptor kinase (TRK) signaling, and platelet-derived growth factor signaling comprise the top five pathways. From the 11 genes upregulated in the fetal wounds, 11 pathways were identified to be relevant to fetal wound healing (Fig. 3C). The top five functional pathways include dolichol and dolichyl phosphate biosynthesis, intrinsic prothrombin activation pathway, chronic myeloid leukemia signaling, IL-6 signaling, and atherosclerosis signaling.

Microarray analysis of E17 fetal and adult wounds at 24 h postwounding.
Discussion
Fetal cutaneous wound healing is uniquely characterized by scarless repair with full restoration of normal dermal architecture. However, in adults, cutaneous wound healing is characterized by physiologic scarring. The goal of this study is to identify candidate pathways important to the scarless wound healing process that might also contribute to decreased scarring and regenerative healing in adult wounds. To achieve this goal, we performed microarray analysis on fetal and adult wounds at three different time points following wounding: 1, 12, and 24 h, to study the temporal activation or suppression of relevant genes to regenerative healing. In addition, to better understand the functionality of observed differential gene regulation, we performed signal pathway analysis. This technique allowed us to identify gene cascades that are regulated during the different phases of wound healing.
Updated discoveries in differential gene expression
Using microarray analyses similar to those previously described by Colwell et al., we found 191, 253, and 27 genes that were differentially expressed between E17 fetal and adult wounds at 1, 12, and 24 h postinjury, respectively. E17 was the earliest gestational age where scarless healing occurs, 6,12 and that allowed reproducible and reliable survival after surgery. The results represent an overall increase from the 175, 134, and 19 differentially expressed genes, at each respective time point, which were identified in the previous study. With the updated gene database utilized in this new study, we were able to detect not only a more accurate but also expanded set of genes that may serve as potential candidates for further study in scarless regeneration. Furthermore, improved pathway analysis abilities provided an objective, unified picture of associated functional pathways rather than solely individual genes, thereby further elucidating possible mechanisms in fetal wound healing. Interestingly, our results reveal some pathways that corroborate earlier hypotheses by Colwell et al., such as the upregulation of protein ubiquitination in fetal wounds at 12 h, while providing additional context for understanding specific prior findings, such as the earlier discovery of greater expression of angiomotin and our new association of the intrinsic prothrombin activation pathway in fetal wound healing 24 h postinjury.
The phenotype of fetal scarless healing is skin regeneration that cannot be differentiated from unwounded skin. Collagen deposition is unchanged and dermal appendages are present after fetal scarless repair. Although our analyses did not solely focus on the phenotypic sequelae, we identified myriad genes and pathways that may play a role in regeneration. Below, we discuss in greater detail some of the most relevant pathways found to be differentially activated in E17 fetal versus adult wounds at various time points following injury. Although the potential roles of these pathways are discussed individually, they may interact together. Such potential interactions are yet unknown and not discussed in this article.
Proline biosynthesis I
The proline biosynthesis I pathway in humans cumulates with the synthesis of
IL-8 signaling
In adult wounds, IL-8 has been shown to be primarily responsible for chemotaxis of neutrophils, which contributes to the inflammatory process. 17 While IL-8 is known to stimulate inflammation in adult wound healing, its role in fetal wound healing remains unclear. Our results demonstrate downregulation of IL-8 signaling pathways in fetal wounds 1 h postinjury in comparison to adult wounds. This diminished IL-8 pathway activation corresponds to a diminished inflammatory response commonly found in fetal wounds as well as the lack of polymorphic leukocyte infiltrate seen in scarless wound healing. 18 Less inflammatory cell recruitment and diminished cytokine release have been suggested to cause decreased paracrine stimulation of extracellular matrix production, as well as increased fibroblast and epithelial cell migration and proliferation. 17 Thus, this lack of inflammatory cascade amplification may be crucial to the formation of an environment conducive to scarless wound healing. Last, excessive IL-8 has been found in disease states characterized by excessive fibroplasia, such as pulmonary fibrosis 19 and psoriasis, 20 further underscoring its contribution to profibrotic processes.
Apoptosis signaling
Apoptosis is critical to the normal progression of wound healing. Especially, as the wound matures, apoptosis plays an important role in the regulation of collagen synthesis and degradation through its regulation of fibroblast and endothelial cell death. Our results indicate that E17 fetal wounds demonstrated upregulation of apoptosis signaling pathways 12 h following wounding in comparison to adult counterparts. This finding is supported by published studies demonstrating a role for increased induction of apoptosis in fibroblast populations as a possible mechanism for promotion of scarless wound healing. 21 Furthermore, recent studies have suggested that decreased rates of apoptosis in mice may lead to the formation of hypertrophic scars and keloids. 22 Taken together, these data suggest that initiation of apoptosis following injury may contribute to scarless wound healing through the programmed removal of damaged and unwanted cells at the site of injury.
CXCR4 pathway
CXCR4 encodes the receptor for stromal cell-derived factor-1 (SDF-1), which acts as a potent chemoattractant for lymphocytes and monocytes 23 and is further responsible for the trafficking of circulating stem and progenitor cells to areas of tissue damage during cutaneous wound repair. 24,25 SDF-1 is often induced by proinflammatory factors, such as tumor necrosis factor-α (TNF-α) and IL-1, 26 and has also been found in elevated levels in fibrotic disorders. Our results demonstrate upregulation of CXCR4 pathway in adult wounds 12 h following injury in comparison to fetal wounds. This finding is in concordance with recent experimental data suggesting that the activation of SDF-1 signaling by proinflammatory factors recruits cells expressing CXCR4 such as fibrocytes, which contribute to the formation of hypertrophic scars. 26 The regulation of SDF-1 and CXCR4 thus provides an important ligand-receptor target for reducing scarring through inhibition of fibrocyte trafficking.
Neurotrophin/TRK signaling
Neurotrophin is synthesized and released by many skin cells, including keratinocytes, melanocytes, and fibroblasts. 27 In skin, neurotrophins help regulate innervation and act as prosurvival and growth factors. As such, they are believed to play an important role in regulating skin homeostasis in both physiologic and pathologic states. Data from our pathway analysis indicate that neurotrophin and its receptor TRK signaling are upregulated in adult wounds 24 h postinjury in comparison to E17 fetal wounds, and as such may play an important role in scar formation. This finding is supported by studies showing that neurotrophin acts to recruit fibroblasts. In addition, injury and inflammation both act to enhance neurotrophin production, mainly by keratinocytes at the site of injury, supporting the role of neurotrophin signaling during wound repair. 28 Altered levels of neurotrophin signaling may thus be expected to contribute to adult scar formation, although no studies to date have looked at this relationship.
Intrinsic prothrombin activation pathway
Thrombin, which is produced from prothrombin, enhances the production of cytokines such as IL-1, 6, and 8, and TNF-α to upregulate the production of growth factors that in turn induce cellular proliferation at the site of injury. 29 This suggests that prothrombin, thrombin, and their receptors may be responsible for modulating the various phases of scar formation. Our microarray results indicate that the intrinsic prothrombin activation pathway is upregulated in fetal wounds 24 h following wounding in comparison to the adult. Increased levels of thrombin and prothrombin have been found in old scars, suggesting that these proteins play an extended role in wound healing beyond the more immediate coagulation following injury. The unexpected result that this pathway is more active during scarless repair suggests an unknown function during extracellular matrix formation and remodeling. Interestingly, studies have found that the administration of thrombin peptides to incisional wounds in rats accelerates normal wound healing and enhances neovascularization. 30 However, more studies are required to delineate the exact mechanisms by which differential activation of this pathway contributes to scarring versus regeneration.
Innovation
Using functional pathway analysis, we demonstrated differential pathway regulation in E17 fetal wounds that undergo scarless regeneration following injury. Due to the large amount of data generated by pathway analysis, we have limited our discussion to pathways known to be particularly relevant to wound healing. We believe that identifying these pathways most likely to be proregenerative or profibrotic provides a valuable foundation for further experimental study investigating mechanisms underlying the regenerative ability of early embryonic skin, with possible applications to other organ systems.
Footnotes
Acknowledgments and Funding Sources
This work was supported, in part, by a grant from NIH grant R01 GM087609 (to H.P.L.), a Gift from Ingrid Lai and Bill Shu in honor of Anthony Shu (to H.P.L.), the Hagey Laboratory for Pediatric Regenerative Medicine and Children's Surgical Research Program (to M.T.L. and H.P.L.), and NIH grant R01 GM116892 (to M.T.L. and H.P.L.). Additional funding was provided by the American Society of Maxillofacial Surgeons (ASMS)/Maxillofacial Surgeons Foundation (MSF) Research Grant Award (to M.S.H., M.T.L., and H.P.L.), the Sarnoff Cardiovascular Research Foundation (to W.X.H.), the California Institute for Regenerative Medicine (CIRM) Clinical Fellow training grant TG2-01159 (to M.S.H.), and the Stanford University School of Medicine Transplant and Tissue Engineering Fellowship Award (to M.S.H.).
Author Disclosure and Ghostwriting
No competing financial interests exist. The content of this article was expressly written by the authors listed. No ghostwriters were used to write this article.
About the Authors