
Abstract
Significance:
Cutaneous wounds are a major problem in both human and equine medicine. The economic cost of treating skin wounds and related complications in humans and horses is high, and in both species, particular types of chronic wounds do not respond well to current therapies, leading to suffering and morbidity.
Recent Advances:
Conventional methods for the treatment of cutaneous wounds are generic and have not changed significantly in decades. However, as more is learned about the mechanisms involved in normal skin wound healing, and how failure of these processes leads to chronic nonhealing wounds, novel therapies targeting the specific pathologies of hard-to-heal wounds are being developed and evaluated.
Critical Issues:
Physiologically relevant animal models are needed to (1) study the mechanisms involved in normal and impaired skin wound healing and (2) test newly developed therapies.
Future Directions:
Similarities in normal wound healing in humans and horses, and the natural development of distinct types of hard-to-heal chronic wounds in both species, make the horse a physiologically relevant model for the study of mechanisms involved in wound repair. Horses are also well-suited models to test novel therapies. In addition, studies in horses have the potential to benefit veterinary, as well as human medicine.

SCOPE AND SIGNIFICANCE
Over 11 million people in the United States are affected by acute wounds yearly, 1 and ∼2% of the population in developed countries suffers from chronic wounds. 2 –4 These numbers are growing, corresponding with increasing rates of diabetes, obesity, and an aging population, all of which predispose to chronic wounds. 5
The impact of wounds in horses is not well documented, but skin wounds are commonly treated by equine practitioners. 6,7 Moreover, a United States Department of Agriculture (USDA) report states that in horses younger than 20 years, 16–27% of euthanasias are carried out in response to injury, wounds, or trauma. 8
TRANSLATIONAL RELEVANCE
At its 2012 annual meeting, The Wound Healing Society drafted a consensus statement that “strongly encourages all wound researchers to involve human wound tissue validation studies to make their animal and cell biology studies more translationally and clinically significant.” 9
The horse is a physiologically relevant model animal, well suited to address questions about human cutaneous wound healing. Horses share anatomical skin features with humans and naturally suffer from wounds reminiscent of human conditions, making them an ideal animal to study particular skin pathologies.
CLINICAL RELEVANCE
The skin serves as a protective barrier to pathogens and chemical compounds, regulates body temperature, prevents dehydration, and allows for sensing the environment. 10 As uncompromised skin is necessary for good health and well-being, treatment of skin wounds is a high priority. The complexity of the cutaneous wound healing process contributes to difficulties in executing successful treatment protocols when spontaneous wound healing is not adequate. Using the horse as a model to study wound healing processes and evaluate novel therapies will improve wound treatment protocols for both humans and horses.
BACKGROUND
The healing of full-thickness skin wounds is far more intricate than implied by the conventional description of the four phases of wound healing. It is clear that each of the “phases” (hemostasis, inflammation, proliferation, and remodeling) is made up of interdependent events executed by resident skin cells, migrating immune cells, and extracellular matrix (ECM) components, in addition to growth factors, cytokines, and chemokines, released by cells and the surrounding environment. 11
The events required for successful wound healing consist of a cascade that is initiated by wounding and continues until long after wound healing is achieved. 12 Conventional treatments for cutaneous wounds of humans and horses broadly aim to improve the quality of the wound bed by keeping it moist, free of pathogens, and adding or removing pressure/tension, depending on the type of wound. 13,14 These methods have remained largely unchanged over the past decades.
In parallel with our increasing knowledge of wound healing processes in humans, there has been an effort to incorporate increasingly sophisticated treatment strategies based on specific cellular responses to injury. Various recombinant growth factors, such as vascular endothelial growth factor (VEGF) and fibroblast growth factor (FGF), have been used to treat wounds in experimental settings. However, malignancy risks and other problems associated with adding recombinant proteins to cutaneous wounds have been recognized, and these therapies have not yet been fine-tuned for clinical use. 15
Hyperbaric oxygen therapy, which consists of breathing pure oxygen, while in a total body chamber where pressure is controlled, is also used as a method to positively influence cells involved in wound healing by altering clotting, cell proliferation, angiogenesis, and tissue repair. 16 Although there are few negative side effects to this type of therapy, it is currently not widely used due to the extensive equipment and technical expertise required, and long-term positive results are not well documented. 16,17
Likewise, equine practitioners have made efforts to refine treatments for cutaneous wounds, based on the growing knowledge of the cellular mechanisms that drive wound repair. The choice of treatment for cutaneous wounds in equine patients depends on the anatomic location of the wound.
The healing of skin wounds on the lower neck and trunk of horses is enhanced by contraction of a thin sheet of striated muscle lying just beneath the superficial fascia, which draws the edges of the wound together. In contrast, wounds on the upper neck, face, and legs close primarily by epithelialization, consisting of the migration of keratinocytes over granulation tissue. Similar to studies for the treatment of human skin wounds, active research is focused on determining the effects of specific proteins, such as VEGF-E and interleukin-10 (IL-10), or oxygen therapy, on the healing of wounds in horses, 18 –21 but as in human medicine, these treatments are not yet available.
In the United States alone, billions of dollars are spent annually to treat the direct and indirect effects of wounds. An evaluation of Medicare data from 2014 revealed that almost 8 million Medicare beneficiaries were affected by chronic wounds, at an estimated cost of $28 to $96 billion. 4 In the United States, over $12 billion is spent annually on indirect costs associated with skin wounds, such as scar removal and lost wages caused by chronic nonhealing wounds. 5
The economic cost of wounds in horses is not well documented, but skin wounds are among the most common ailments treated by equine practitioners. 6,7 Moreover, a USDA report states that in horses younger than 20 years, ∼16–27% of euthanasias are carried out as a way to humanely deal with injury, wounds, or trauma. 8 The costs associated with the management of cutaneous wounds, as well as the lack of adequate treatment options for both humans and horses, create an urgency for basic scientific research designed to understand the mechanisms that drive normal and delayed wound healing, and the development and testing of targeted adjunctive and novel therapies in both human and veterinary medicine.
The goals of this review are to (1) describe the common limitations of current wound therapies for humans and horses, which, in part, result from an incomplete understanding of wound healing processes, (2) compare skin physiology and normal wound healing in humans and horses, (3) propose the horse as a model for two types of chronic skin pathologies suffered by humans, and (4) offer evidence that the horse is an interesting model to test novel therapeutics for cutaneous wound management. Based on the information provided in this review, we put the horse forward as a relevant model for wound healing studies, in particular those relating to wound contracture or naturally developing fibroproliferative disorders, which will benefit both human and veterinary health.
DISCUSSION
Skin physiology and normal wound healing in humans and horses
Skin physiology
The skin of humans and horses comprises two principal layers that protect internal organs from mechanical damage and invasion by pathogens. The outer epidermis, derived from the embryonic ectoderm, is a thin, cellular layer that provides a waterproof barrier to the environment. The dermis lies beneath the epidermis and is made up of two regions, the papillary dermis and the reticular dermis. This layer contains fibroblast cells that produce and remodel the dense, collagenous connective tissue that makes up the bulk of the dermal layer. The dermis is also rich with blood vessels, lymphatic vessels, and glands, such as sweat and sebaceous glands. Hair follicles originate in this layer as well. Epidermal extensions, referred to as rete ridges or pegs, extend into the dermal layer.
Resident and infiltrating immune cells can be found in both layers of the skin. The hypodermis, or superficial fascia, is located below the dermis and primarily consists of fat and connective tissue, but also contains blood vessels. This layer is not considered part of the skin, but rather serves as an insulating layer and helps attach the skin to underlying structures.
The human epidermis comprises four to five layers of stratified epithelial cells, 22 while the horse epidermis is described as being made up of five stratified squamous cell layers. 23,24 Human and horse skin both have a thick dermal layer, which in the human contains sparse hair follicles, while the hair follicles in the horse are densely distributed (Fig. 1A). 22,25 Human skin thickness is relatively uniform from site to site on the body, 26 while the thickness of horse skin is variable according to body site. 14
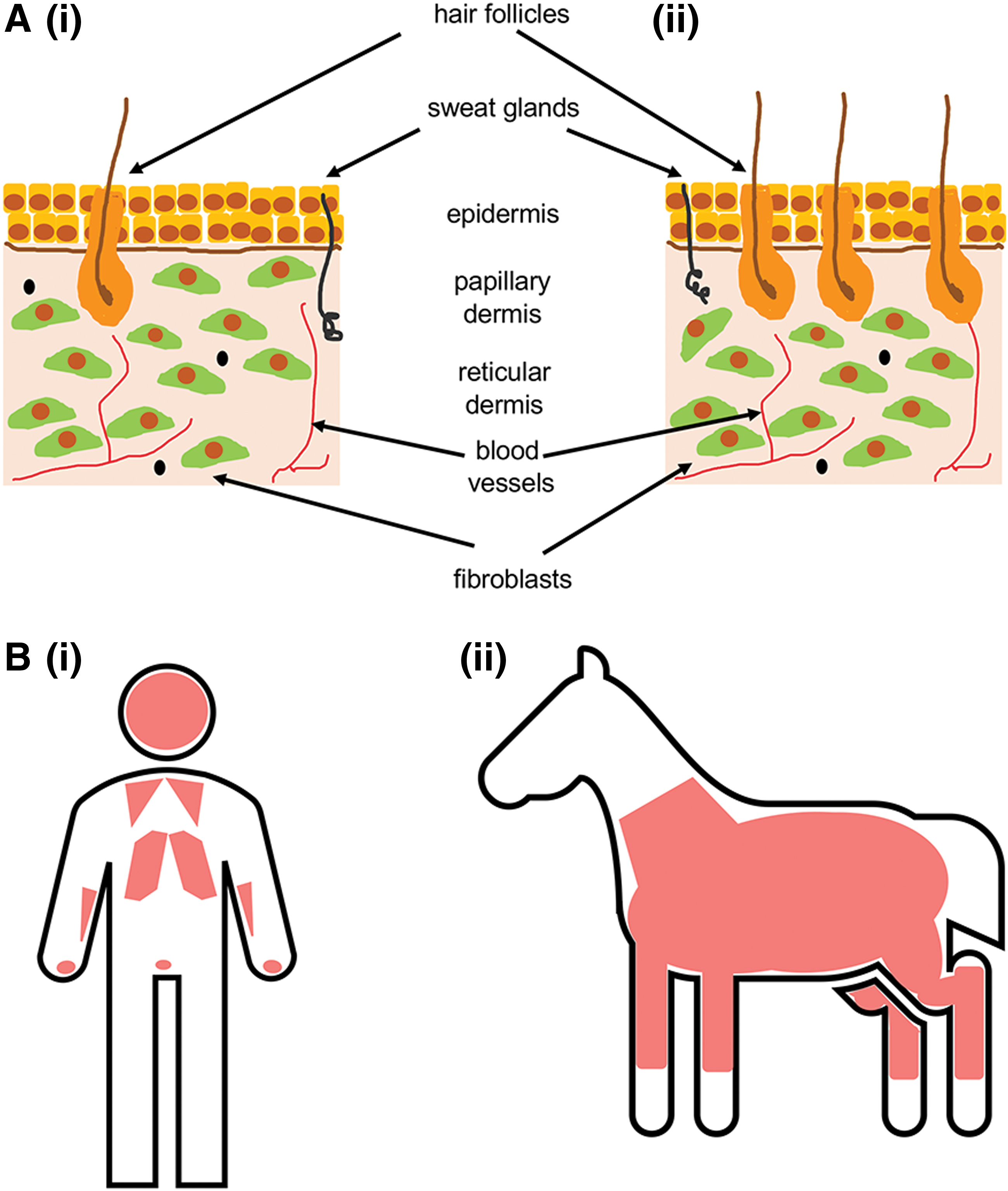
Human and horse skin physiology.
Humans are considered “tight-skinned,” meaning that the skin is firmly attached to the underlying subcutaneous connective tissue. 22 Horses are often also described as “tight-skinned,” 27 but since the qualities of horse skin vary across the body, some parts of the horse, such as the trunk, are considered to have “loose-skin,” which is more typical of that of laboratory rodents. Humans and horses both have a disrupted panniculus carnosus, a layer of striated muscle beneath the hypodermis. In humans, remnants of this muscle are found in the face, hands, chest, and scrotum, 28 whereas it stretches from the knees to the base of the neck in horses (Fig. 1B). 29
Normal wound healing
Similar to most mammals, successful skin wound healing in humans and horses involves a network of synchronized biological processes that restore the integrity of the skin after injury. The process of wound healing after full-thickness injury is commonly divided into four phases (Fig. 2). The duration and intensity of these phases vary depending on species, anatomical location, and wound size.
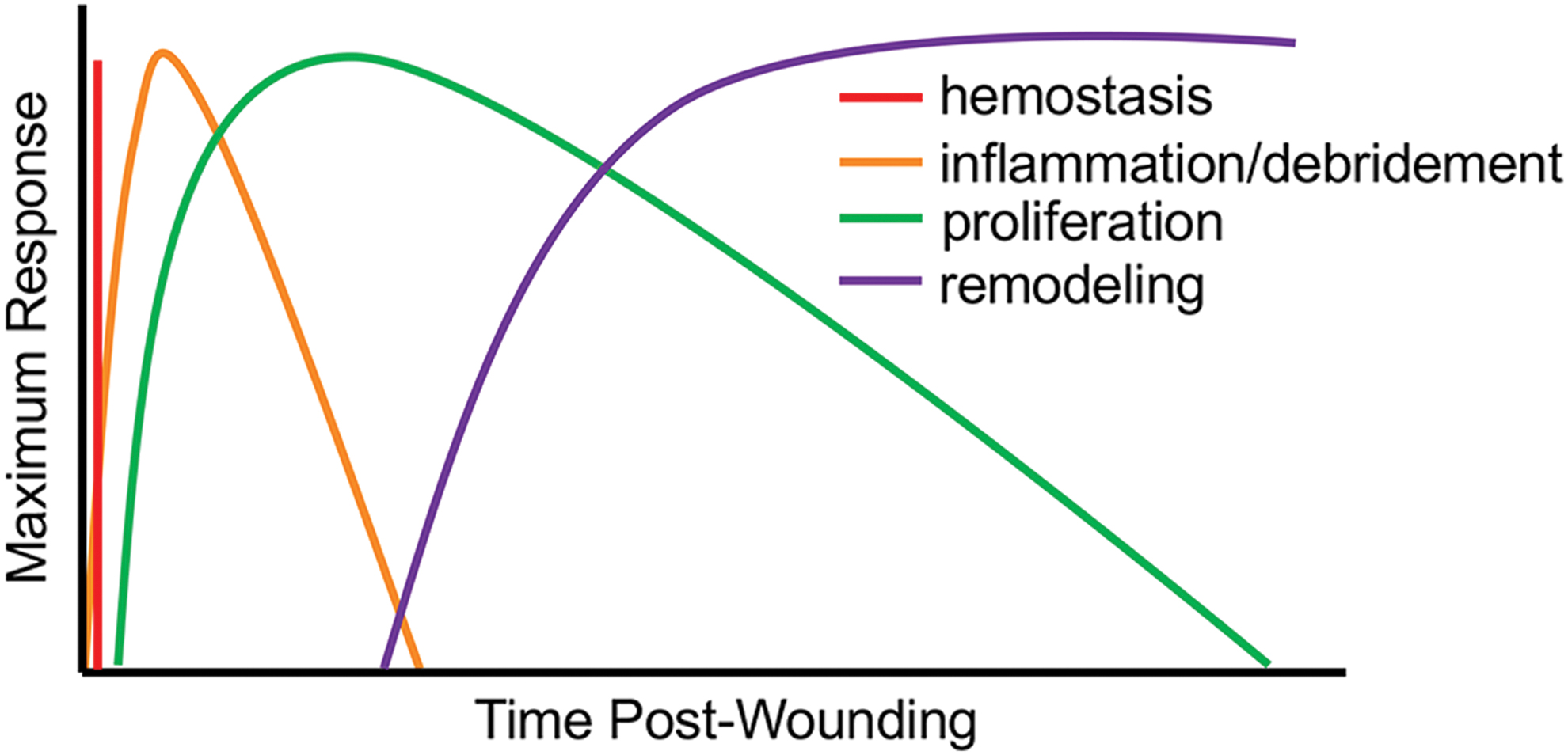
Phases of normal wound healing in mammals. Normal wound healing in mammals comprised an overlapping series of events. Immediately after injury, bleeding is stopped by hemostasis. An intense inflammatory response follows, which serves to clear pathogens and induce proliferation of cells needed to fill the wound space. The remodeling phase continues long after wound healing. In this phase, stromal cells coordinate the turnover of extracellular molecules in the wound bed, leading to increased tissue strength. Color images are available online.
During hemostasis, the immediate response to injury, vasoconstriction, and coagulation serves to stop blood loss and to concentrate the growth factors that contribute to wound healing. The subsequent inflammatory or debridement phase is characterized by infiltration of immune cells that control contamination and secrete cytokines and growth factors, which induce the proliferative phase. 30 During the proliferative phase, epithelial cells migrate from the wound margins to close the wound, and a population of dermal fibroblasts (DF) are activated by tumor necrosis factor alpha (TNFα), platelet-derived growth factor (PDGF), transforming growth factor beta 1 (TGFβ-1), and mechanical stress in the wound bed. 31 Activated DF secrete collagen and other ECM proteins and may differentiate into myofibroblasts, which contribute to wound contraction. Finally, during the remodeling phase, ECM proteins are alternately laid down and degraded, creating an increasingly organized tissue environment 30 (Fig. 2).
In many cases, however, complete tissue regeneration is not achieved, leading to chronic nonhealing wounds or abnormal scar formation. As the details of wound healing processes are gradually understood, it has become apparent that the four phases of healing are not distinct, rather the events required for successful wound healing are a continuous cascade that is initiated by wounding and continues until long after the wound has closed. 12
Due to similarities in skin architecture, aspects of normal cutaneous wound healing in humans and horses are very much alike (Table 1). Humans have skin that is firmly attached to the underlying connective tissue and as a result, rely on epithelialization for the primary surface healing of skin wounds, followed by dermal healing by stromal granulation and myofibroblast migration and contraction. This is very similar to how skin healing progresses on the distal limbs of horses. 32 In contrast, the mechanism of primary wound closure on the horse trunk is more similar to that seen in rodents, which involves contraction of the panniculus carnosus. 33
Normal wound healing in humans and horses
Another concordance between humans and horses that may contribute to similarities in skin wound healing is the ratio of neutrophils to lymphocytes (NLR) in peripheral blood. Human and horse blood each have a ratio of about 60% neutrophils to 40% lymphocytes, which is much higher than the NLR of mouse blood. 26 In human medicine, the NLR in blood, as well as in tissues, is emerging as a measure of subclinical inflammation. 34 –38 Since inflammation is a critical aspect of cutaneous wound healing in mammalian species (Fig. 2), similar basal NLR may contribute to similarities in this phase of wound healing.
In addition to parallels, there are basic physiological differences between humans and horses that may contribute to distinct mechanisms of skin wound healing between these species (Table 1). Humans can be classified as stalking predators, whereas horses are flight animals, with hearts capable of going from a resting rate of 16 beats/min to 240 beats/min almost instantly. This extreme flight response results in contraction of the spleen, which releases a reserve of red blood cells into the circulation, increasing the oxygen-carrying capacity of blood to levels far higher than possible in humans. 39 As successful skin wound repair depends on a steady supply of well-oxygenated blood, it stands to reason that the different patterns of blood oxygen availability in humans and horses may result in alternative mechanisms for healing between the species.
A subtler difference between the species is the platelet composition of the blood; human blood has a much higher concentration of platelets than horse blood, 40 which may affect the rate of coagulation and alter the initial inflammatory response to wounding. There are no published reports describing how these basic physiological differences between humans and horses lead to differences in skin wound healing between the species, but an awareness of them may help researchers determine which human skin conditions can be accurately recapitulated in the horse model.
The horse as a model for cutaneous wound healing research
Researchers have been studying the mechanisms of cutaneous wound healing in humans and horses for decades. For an overview of the models used for human wound healing research, the reader is referred to other reviews. 41 –46 In this review, we will focus on the models, both in vitro and in vivo, used to study the (patho)physiology of wound healing in horses.
Since body wounds of horses tend to heal by wound contraction, whereas skin wounds on their distal limbs heal primarily by epithelialization, similar to wounds in humans, the horse provides a unique model in which wound healing by different mechanisms can be studied within the same animal, eliminating interindividual variation. Moreover, the size of horses also provides the following experimental benefits: (1) multiple experimentally induced wounds can be generated at one site on an animal, allowing for controls and treatments to be performed on the same individual, and (2) multiple samples of wounds can be collected over time to evaluate healing processes in a way that cannot be performed in humans or small animal model species.
Naturally occurring chronic skin pathologies similar in humans and horses
Several types of naturally occurring skin pathologies of horses are uniquely similar to conditions experienced by humans, making the horse an ideal animal model to study them. Two such naturally occurring wound conditions, namely fibroproliferative wounds and compromised skin resulting from allergic responses, are particularly relevant because the development of these wounds is not well understood and treatment options in both species are limited.
Fibroproliferative disorders
The formation of normal granulation tissue is an essential process in successful cutaneous wound healing in both humans and horses. 32,47,48 Granulation tissue is created by fibroblasts and is primarily composed of type-III collagen. It fills the wound space, forming a barrier between the body and external contaminants, and provides epithelial cells with a surface over which to migrate. Granulation tissue also serves as support for cells that contribute to wound repair. Endothelial cells in the granulation tissue form new capillaries, needed to bring oxygen and immune cells to the wound bed. Fibroblasts secrete ECM molecules, and the enzymes required to break them down during the rounds of ECM remodeling that occur over the course of tissue repair. 49
Fibroproliferative disorders, characterized by the deposition of excessive ECM or granulation tissue, are associated with chronic low-grade inflammation, cellular proliferation, and under some conditions, lack of wound closure. Fibroproliferative tissue develops uniquely in humans, often as hypertrophic scars and keloids, and on the distal limbs of horses, where it is called proud flesh or exuberant granulation tissue (EGT). Keloids typically develop after reepithelialization has occurred, whereas EGT is characterized by delayed reepithelialization. Table 2 provides a comparative overview of key features of the fibroproliferative tissue that makes up human keloids and equine EGT.
Human keloid compared to horse exuberant granulation tissue
EGT, exhuberant granulation tissue.
Keloids are defined as scars that grow continuously and invasively beyond the margins of the original wound, which can range from pea sized to as large as a football. 50 The quality of life of patients suffering from keloid disease can be reduced due to both physical complications, including severe itching, pain, loss of mobility, and infection, as well as psychological impairments, such as embarrassment, lack of self-confidence, and depression. 51
There is no single, effective treatment regime, and although many therapies are available, most are associated with extremely high rates of recurrence. 52 Recent reviews of keloid therapies discuss a wide range of noninvasive medical therapies, including laser and light-based treatments, pressure garments, and silicone gel sheeting, as well as the injection of corticosteroids and compounds that inhibit cell proliferation and stimulate apoptosis. Surgical removal of keloids and cryotherapy is also practiced. 53,54 Exploring novel therapies for keloids is an active area of research. Growth factors, botulism toxin, calcium blockers, and the mTOR inhibitors that are currently used as chemotherapy for cancer patients, are all currently under investigation for their use in treating keloids. 54 –56
Keloid development involves alterations in complex pathways, but the mechanisms of initiation and regulation of these pathways in the context of keloid scarring are unknown. 57 Although chronic inflammation has been shown to contribute to keloid formation, details of keloid pathogenesis in the early stages are poorly understood. In response to trauma, 15–20% of people of Asian, African, and Hispanic origins develop keloids. There is evidence of genetic susceptibility to keloids, but the underlying pathogenesis is not well defined. 52
As described above, horse skin wounds heal by distinct mechanisms according to the body site at which they occur. Wounds on the distal limbs that heal by epithelialization usually close slowly and can develop EGT. EGT is visually irregular with a pebbled, moist surface that protrudes beyond the margins of the wound. This phenotype is a result of residual fibrin deposits, disorganized fibroblasts and vasculature, and a lack of epithelial cell coverage. 14 As this tissue is exposed to the environment, it is susceptible to abrasion, secondary infection, and reinjury.
Removal of EGT by surgical excision is currently considered the best treatment option, even though the procedure often needs to be performed multiple times before successful wound closure occurs. 58 Interestingly, ponies, which are the same species as horses, but are distinguished based on size (ponies are <1.48 m in height at the withers, while horses are taller), rarely develop EGT in association with limb wounds. The intriguing differences in wound healing according to anatomic location in horses, and in horses versus ponies, have been the focus of many in vivo studies that led to what is known about EGT, and are described in more detail in the Equine in vivo skin wound models section. Aside from differences in incidence of EGT between horses and ponies, there is currently no evidence to clearly support breed-specific or genetic predisposition to EGT.
A comparative histological study of keloids and EGT determined that both tissues are characterized by disorganized thickened collagen fibers, minimal numbers of tissue macrophages and mast cells, moderate-to-severe dermal fibrosis, and similar numbers of fibroblasts. Elastic fibers were not observed in either tissue type. Granulation tissue was absent in keloid tissue, but present in EGT. Dermal vascularity was moderate to severe in keloids, and minimal in EGT. Keloidal collagen and myofibroblasts were observed in keloids, but not in EGT (Table 2). 32
Gene and protein expression have been well studied in keloids and EGT, as well as in individual cell types isolated from these tissues. Analysis of mRNA and protein from keloids has shown that the expression of TGF-β1, matrix metalloproteinase-1 (MMP1), FGF12 and FGF13 is increased in this tissue compared to normal skin. 59,60 These growth factors directly and indirectly contribute to a prolonged low-level inflammatory phase of wound healing and delay the resolution of the fibroproliferative phase. 57
Early work measuring growth factor concentrations in homogenates made from horse wounds located on the limb and on the thorax demonstrated that TGF-β1 is more highly expressed in limb versus thoracic wounds, while transforming growth factor beta 3 (TGF-β3) and FGF expression patterns are similar irrespective of wound site. 61 A general analysis of the inflammatory mediators in the TGF-β family in wounds in horses and ponies showed that, while wounds on ponies express higher levels of active TGF-β postwounding regardless of wound location (trunk or limb), wounds on horse limbs that are prone to develop EGT exhibit a sustained low-level expression of this family of growth factors. 62 Although not specifically evaluated in this study, the authors did speculate that in addition to overall expression levels, the ratios of the TGF-β1, TGF-β2, and TGF-β3 isoforms to each other are likely to contribute to the formation of EGT. Building on this work, a study was performed to analyze the function of polymorphonuclear leukocytes (PMN) as well as the local cytokine profiles in limb and neck wounds of horses and ponies. 63 The results of this study indicated that pony PMN produce higher levels of reactive oxygen species compared to horse PMN, and that TNFα, IL-1, and chemoattractants are produced at higher concentrations in pony wounds than in horse wounds.
Collectively, these suggest that an intense, yet temporally limited inflammatory response supports efficient wound closure, while a sustained low-level inflammatory response promotes excessive fibrosis in both humans and horses (Fig. 3).
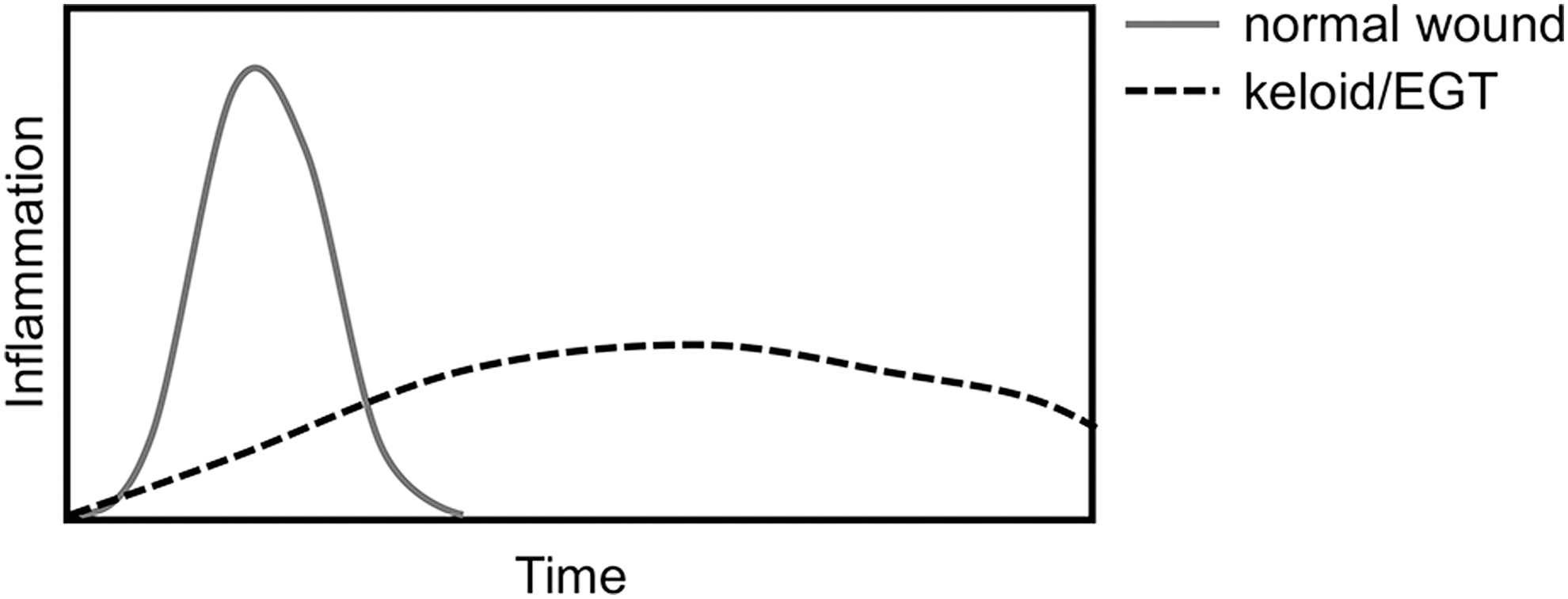
Illustration of the inflammatory response in normal wound healing and during the development of keloid or EGT. Normal wound healing is characterized by an intense and short-lived inflammatory response immediately after wounding. During the development of human keloid or of EGT in horses, the inflammatory response is delayed, suboptimal, and does not fully resolve.
Suppression subtractive hybridization was used to identify genes differentially expressed in the margins of normally healing wounds compared to intact skin in horses. 64 This work resulted in a list of 226 genes expressed 7 days postwounding. The authors then used these data to compare gene expression patterns in samples taken from normally healing wounds on horse trunks, as opposed to samples collected from wounds on horse limbs, predisposed to EGT formation. Annexin A2 (ANXA2) and MMP1 mRNA were found to be upregulated in trunk, but not limb wounds, corresponding to the rapid tissue turnover required for normal wound healing. 65
Another study by the same group examined the expression, both on mRNA and protein levels, of laminin receptor 1 (RPSA) in healing wounds on the bodies and legs of horses. 66 This work showed upregulation of RPSA in thoracic wounds at 4 to 6 weeks postwounding, when epithelialization was occurring, while expression in limb wounds was downregulated at postwounding times. Data on the expression patterns of RPSA, a receptor that binds multiple ligands, including a variety of ECM molecules, may help tease out events that favor normal wound healing over the development of excessive fibrotic tissue and chronic wounds.
Follow-up work showed that OB-cadherin, a protein positively associated with contractile fibroblasts, was higher in thoracic than limb wounds. 67 In addition, osteopontin (OPN), which is essential for proper ECM organization in normal wound healing, was also differentially expressed in horse wounds depending on wound location, which the authors suggest may reflect the delayed and low-level inflammatory response characteristic of EGT. 68
Microvascular occlusion, the excessive proliferation of fibroblasts, and a skewed apoptotic balance are all characteristics of keloids and hypertrophic scars in humans, and of EGT in horses, which may contribute to the excessive deposition of ECM (Table 2). 69 –72 Tissue hypoxia has been implicated as a contributing factor to fibroproliferative diseases in humans, although the underlying mechanisms are not fully understood. Studies in horses indicate that microvascular occlusion leads to a relative state of hypoxia in limb wounds, which may promote the weak, but prolonged inflammatory response characteristic of chronic, fibrotic wounds. 73,74
Based on the similarities between the two diseases, and the benefits provided by studying cutaneous pathologies in a large animal species (discussed in the Equine in vivo skin wound models section), EGT in horses has been proposed as a model for the study of keloids in humans. 27
Atopic dermatitis
Another chronic inflammatory skin disease experienced by both humans and horses is atopic dermatitis (AD). Intense itch and recurrent eczematous lesions, triggered by exposure to harmless environmental allergens, characterize this increasingly common condition. Clinical signs of AD in humans often develop during childhood and include pruritic lesions, hives, lichenified skin, and staphylococcal colonization. 75 –77
AD is frequently accompanied by asthma and allergic rhinitis in humans, 78 –80 and food allergies are another associated condition. 81,82 As the tendency to have an allergic condition is associated with susceptibility to additional allergic conditions, symptoms often appear in sequence with age, supporting the concept of an “atopic march.” 75 AD in humans is also associated with nonatopic comorbid conditions, including other autoimmune-mediated diseases as well as depression and anxiety. 83 –85
Patients with AD often present with ultrastructural abnormalities of the stratum corneum, characterized by dry skin and compromised barrier function. These abnormalities have been linked to mutations in the filaggrin (FLG) gene, which lead to absence of this protein in the outer layer of the skin. 86,87
AD in humans has historically been considered IgE mediated, supported by an exaggerated T helper 2 (Th2) response, 88,89 but this view is expanding into one in which epidermal barrier disruption, the responses of various T cell subsets, and dysbiosis of the commensal skin microbiota all contribute to the pathology. 75 Although it is proposed that molecules secreted upon damage of the epidermis and products of the immune cells that respond to them are increased in AD lesions, the cytokine profiles in skin affected with AD have not been accurately defined. It has been shown, however, that epithelial cells from AD patients express high levels of thymic stromal lymphopoietin, a mediator that indirectly stimulates T cells to produce Th2 cytokines in other pathologic situations. 90
Increased mRNA levels of the Th2 IL4, IL10, and IL13, and the Th22-associated cytokine IL-22, have all been detected in acute AD lesions when compared to healthy skin. 91 CD4+ T cells from AD lesions produce IL-4, IL-13, and IL-22, whereas CD8+ T cells from AD lesions are a source of IL-13, IL-22, IL-17, and interferon gamma (IFNγ), 92 and type 2 innate lymphoid cells (ILC-2) in AD lesions express IL-4. 89
AD in horses presents both as a skin and a respiratory disease, but evidence of a progressive atopic march is only anecdotal in horses. 93 Clinical signs of AD in horses include hives and pruritus, and similar to human AD, staphylococcal colonization is commonly found. 93 Food allergies can lead to AD in horses, although other conditions, such as hypersensitivity to proteins in the saliva of biting midges in the genus Culicoides, are more commonly associated with equine AD. 94 Ultrastructural abnormalities in lipid lamella and disorganization of the stratum corneum, which resembles skin lesions in human AD, have been reported in horses. 95 The cytokine profiles in the skin and/or serum of horses that are sensitive to AD have not been described thoroughly, but it has been shown that an increased expression of IL-4 is associated with the disease. 96,97
The horse is currently not actively used as an animal model for the study of human AD, but efforts to increase our knowledge of equine AD and to better define similarities and differences will provide better insights into the suitability of the horse as a model for AD in humans. Some of the key features, both similar and different, that have been studied in both species are shown in Table 3.
Human and horse atopic dermatitis
FLG, filaggrin; Th2, T helper 2.
Equine in vitro skin wound models
In vitro cell culture models have been used extensively to study the characteristics of cells involved in equine cutaneous wound healing, such as resident skin cells and migrating immune cells, as well as their responses to potential treatments.
Two-dimensional cell culture
Because of their simplicity, two-dimensional (2D) cell models are heavily used to define basal characteristics of skin cell populations such as cell morphology, population doubling time, gene and protein expression patterns, and cell motility. They are also useful to detect changes in target cells in response to an insult, treatment, or change in culture conditions.
For example, the inherent growth characteristics of DF isolated from the trunk and distal limbs of horses and ponies have been evaluated in a 2D culture system. 98 Results from this study showed that DF isolated from the horse limb grow more slowly than DF isolated from the horse trunk, as well as from pony limb and trunk, and led the authors to conclude that differences in DF growth may contribute to the development of EGT. The authors speculated that decreased fibroblast growth may lead to reduced numbers of myofibroblasts, poor wound contraction, and dysregulated ECM deposition and remodeling during the healing process. Consequently, influencing DF growth could be a potential way to control the development of EGT.
Another study compared basic characteristics of DF isolated from healthy horse skin and horse EGT in a 2D monoculture model. 99 This work showed that DF cultured from EGT display a more cuboidal morphology in culture compared to DF cultured from healthy skin, and that DF isolated from EGT have a longer population doubling time, migrate more slowly, and have gene expression profiles that are distinct from those observed in DF isolated from healthy skin. Moreover, EGT-derived DF were found to be more susceptible to busulfan-induced senescence than DF derived from healthy horse skin. These observations could provide clues as to why EGT fails to resolve in vivo.
In addition to DF, primary equine keratinocyte cultures 100 and equine digital vein endothelial and smooth muscle cell cultures 101 have been established in 2D culture systems. As these cell types are all involved in skin wound healing in vivo, they can be used in future experiments to learn more about the cell–cell and cell-ECM interactions that occur during normal and impaired wound closure.
Migrating immune cells can be isolated from horse blood, characterized in culture, and then used in various 2D and three-dimensional (3D) coculture models serving to observe cell–cell interactions relevant to wound healing processes. For example, CD4+ T cells were purified from horse and pony peripheral blood mononuclear cell preparations and cultured, allowing for the identification of a subpopulation of FOXP3+ cells analogous to regulatory T cells in other species. 102,103 As these cells are known to be dysregulated in human AD and fibroproliferative tissue, they may prove to be a useful tool for in vitro wound healing studies with equine cells.
Cultured PMN isolated from horses and ponies showed that (1) reactive oxygen species production of pony PMN is higher compated with horse PMN and (2) in vitro chemotaxis of PMN differs between those isolated from ponies and horses, providing the basis that these cells can contribute to differences observed between horse and pony skin wound healing. 63 Equine monocyte-derived macrophages 104 and mesenchymal stromal cells (MSCs) 105,106 are additional examples of migratory cells that have been characterized using 2D cell culture systems and contribute to skin wound healing.
Three-dimensional cell culture
A variety of 3D cell culture systems have been developed for the study of equine cutaneous wound healing (Fig. 4). These models are often considered more anatomically relevant than 2D cell culture, as they incorporate both a cellular support system or matrix and/or multiple cell types, which allow for the establishment of cell–matrix and cell–cell contacts that mimic the in vivo wound microenvironment. 107

Three-dimensional equine in vitro cell culture models. Collagen gels are used to determine the contractile capacity of cells, which can be quantified. Organ cultures, which maintain in vivo architecture, are useful to observe responses to treatment and monitor cell–cell interactions, but their lifespan in vitro is limited. Uniform dermal equivalents are used in automated wound healing studies. Organoids, which have a long lifespan in culture and exhibit a high degree of cellular organization, are used to observe responses to treatment, monitor cell–cell interactions, and study disease progression. Color images are available online.
The contractile capacity of equine DF isolated from the buttocks and distal limb of horses and ponies was determined by taking advantage of relatively simple 3D cell culture models, growing equine DF in both floating and anchored collagen gels. 108 From these studies, it was concluded that differences in in vivo contraction between wounds on the limbs versus buttocks, and between horses versus ponies, are not caused by the inherent contraction capacity of DF at wound sites, suggesting that the activity of these cells in vivo is determined by the wound environment.
Organ cultures are 3D models that are perhaps the most directly reminiscent of the in vivo environment. For these models, organ samples are collected and maintained in medium, with or without physical support, for several weeks. 109 Organ cultures are primarily used to compare the responses of tissue to treatments, 110,111 test the toxicity of compounds, 112 and monitor cell migration and epithelization after wounding. 113 While organ cultures (1) contain a greater diversity of cell types than can be incorporated in an engineered 3D model and (2) maintain the complex architecture of the skin, they have a limited life span in culture.
Equine organ cultures have been used as a source of hair follicle-derived stem cells, 114 as well as primary keratinocytes, 115 and we are currently optimizing an equine skin explant model to study bacterial biofilm growth in skin wounds. Equine skin organ culture models that recapitulate skin morphology and function are also currently being developed and validated for the study of wound healing as well as for drug testing. These models are created from full-thickness skin collected from horses, allowing for the ex vivo manipulation and analysis of samples from the trunk or limbs that may exhibit inherently different responses to wounding or wound treatments. Epithelialization is a specific aspect of wound healing that these models are well suited to address. Cultured skin biopsies are able to form peripheral epithelial tongues, mimicking the process of reepithelialization, which includes keratinocyte migration, proliferation, and differentiation. Reepithelialization can be accurately measured in organ cultures over time in a way that is not possible in other model systems. 116
Dermal equivalent models are another type of 3D cell culture model, often comprising DF, embedded in a solid ECM, and seeded with an “epidermis” of confluent keratinocytes. In addition, a hypodermis of adipose tissue can be incorporated. 117 Over several weeks in culture, the keratinocytes will form several layers, including the stratum corneum. These models are typically less complex than tissue explants, but since it is possible to generate many uniform dermal equivalents that may be maintained for a relatively long time in culture, they are considered ideal for automated wound healing studies and toxicity testing. 118,119
Recently, an equine dermal equivalent model consisting of “normal” horse DF and keratinocytes, supported by a collagen matrix and resembling equine skin morphologically and structurally, was developed. 120 Although not currently available, construction of similar models using cells isolated from EGT or from animals suffering from AD, with or without the addition of immune cells, could prove to be a productive and physiologically relevant way to study pathological wound healing processes in vitro.
Finally, organoids are defined as ex vivo multicellular fragments containing the major cell types of a particular organ that approximates in vivo organization. One strategy for developing organoids is to culture multipotent or pluripotent stem cells in a 3D matrix under conditions that promote self-organization of the cells. As these models achieve a level of cellular organization not possible with dermal equivalents, and they can be maintained in culture longer than tissue explants, they are useful models for the study of pathologies, cell–cell interactions, and drug discovery. 121 –123 Murine and human organoids have been developed from retina, kidney, liver, intestine, stomach, lung, and brain, 121 and very recently, from skin. 124 Equine intestinal organoids are currently the only organoid type described in the horse, 125 although we anticipate that equine organoids from additional tissues, including skin, will be established in coming years.
Equine in vivo skin wound models
In vivo studies in horses have provided researchers with a well-controlled environment to study conventional therapies such as bandaging and ointments, 126 –128 and they have been used to evaluate the efficacy of full-thickness grafts, 129 to understand complications of sores associated with casting, 130,131 and to elucidate the mechanisms responsible for AD. 94,132 However, the majority of experimentally induced and controlled horse wound studies have been carried out with the goal of increasing our understanding of the spontaneous development of cutaneous fibroproliferative disorders.
The horse model for experimentally induced EGT was developed and verified in 2001, 61 and has since been adopted by many veterinary researchers. 21,72,133,134 In this model, multiple full-thickness (6.25 cm2) wounds are created on the lower limb using a scalpel. Wounds are positioned 1.5 cm apart and are typically arranged in a vertical column. Postoperative bandaging reliably induces the formation of EGT (Fig. 5A). Full-thickness wounds of the same size, inflicted on the thorax or neck, and which do not develop EGT, can serve as controls (Fig. 5B). This model has allowed for the study of the effects of wound oxygenation and temperature on EGT development, 73,74 and the evaluation of gene and protein expression patterns in EGT compared to wounds that heal quickly. 66 –68,135,136
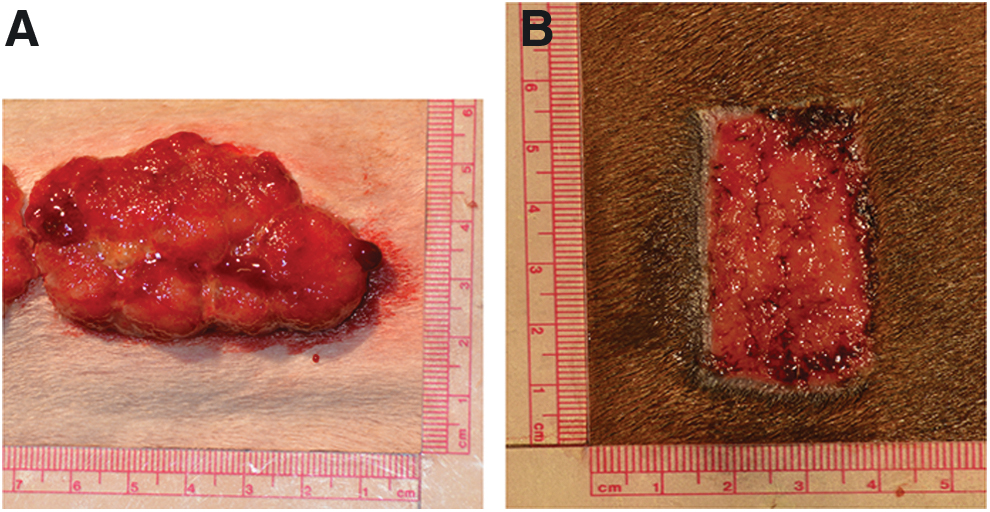
Experimentally inflicted horse skin wounds, 14 days after creation.
The horse as a model to explore novel therapeutics for wound management
As a large animal, with physiological concordance to humans, the horse may serve not only as a model for the biological processes involved in wound healing but also as a translational species to study novel therapies. Benefits of using the horse to test new wound management therapies include (1) the ability to experimentally inflict well-controlled skin injuries instead of having to rely on human patients enrolled in clinical trials with different stages of chronic wounds and/or additional underlying diseases, and (2) testing alternate therapies for skin wounds in experimentally inflicted wounds of horses allows for earlier invention than is possible in humans, where unconventional treatments are usually only administered as “a last resort.” Finally, progress in the development of novel therapies in an equine model will not only benefit humans but also equine patients. The therapies described in this section are currently being developed for both humans and horses.
Stem cell therapies
Stem cells, or their products, are a type of cell-based therapy that may be used to augment the natural events required for successful wound healing, while avoiding the side effects associated with administering recombinant factors and the limits associated with a technology-intensive procedure. 137 Stem cells are defined as undifferentiated cells that can (1) self-renew, maintaining the stem cell pool, and (2) differentiate into mature cell types. Stem cells are classified based on their potency. Totipotent stem cells, which are present during the zygote stage, can differentiate into all cell types of the body. Pluripotent stem cells, such as embryonic stem cells (ESC) and induced pluripotent stem cells (iPSCs), can give rise to almost all of the cells that comprise an organism, with the exception of placental tissues. Multipotent stem cells are found after birth, and these adult tissue-specific stem cells exhibit the ability to differentiate into any of the cell types that arise from their germ layer of origin (Fig. 6). Additional cell types exhibiting varying degrees of potency, such as transit-amplifying cells, develop in the space between true stem cells and terminally differentiated cells. These cells may also prove to be useful therapies for cutaneous wounds, as they are highly relevant to skin maintenance and repair.
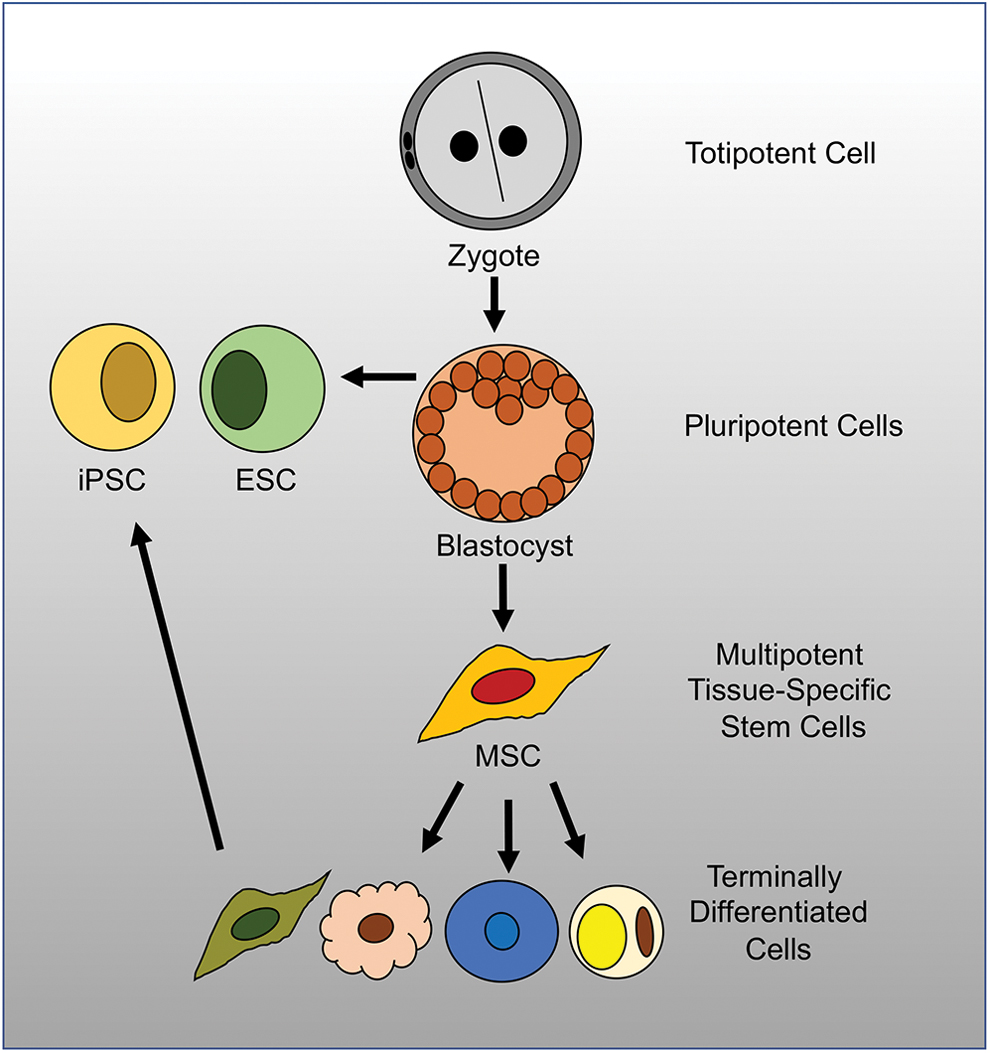
Stem cell hierarchy. The totipotent cells of the zygote can differentiate into all cell types of the body. The ESCs of the blastocyst are pluripotent stem cells that can give rise to most of the cells making up an organism. MSCs are considered to be multipotent tissue-specific stem cells, able to differentiate into the cell types making up their specific tissue of origin. Terminally differentiated cells can be reprogrammed into iPSCs, with a potential analogous to that of ESC. ESC, embryonic stem cells; iPSC, induced pluripotent stem cells; MSC, mesenchymal stromal cells. Color images are available online.
Embryonic stem cells
Human ESCs with a limitless capacity to self-renew and the ability to differentiate into almost any cell type could be therapeutically valuable, but ethical issues associated with collecting cells from embryos have largely hindered their evaluation for this purpose. Ethical concerns are partly alleviated when studying ESCs isolated from veterinary species, but these cells are currently not actively being used in equine medicine. Putative equine ESCs have been described and validated based on marker expression and differentiation potential, but did not form teratomas when injected into immunocompromised mice, which is the gold standard for human and murine ESC verification. 138 After this first description, additional studies with equine ESCs have been carried out in relation to their potential for treatment of tendon injuries, 139 –142 but these cells, to our knowledge, have never been explored as a therapy for cutaneous wounds.
Induced pluripotent stem cells
iPSCs are adult cells, often skin fibroblasts, which have been reprogrammed from a differentiated state into a less-differentiated state, typically by the introduction of transcription factors specific to pluripotent cells. 143 Human iPSCs were first derived from adult human DF in 2007. 144 Since then, they have been differentiated successfully into cardiomyocytes, cells of the skeletal system, hepatocyte-like cells, and cells of the skin, with the goal of generating cells and tissues that can be used for transplantation. 145 –148 In human medicine, the skin has been recognized as an ideal organ for iPSC-based applications. Skin equivalents or other 3D models can be generated from iPSCs derived from patients with inherited genetic skin conditions, allowing for the study of these diseases in vitro. 148,149 In addition, disease-causing mutations can be corrected in iPSCs, allowing for the generation of gene-corrected replacement skin for patients suffering from genodermatoses. 150
Equine iPSCs have been generated from fetal fibroblasts and keratinocytes. iPSCs derived from horse fibroblasts expressed endogenous markers associated with pluripotency, displayed a stable karyotype over long-term culture, and formed teratomas containing all three embryonic germ layers when grafted into mice. 151,152
Equine fetal fibroblast-derived iPSCs have also been differentiated into a keratinocyte lineage with a higher proliferative capacity than primary keratinocytes. 153 The motivation for this work was to produce functional keratinocytes that can be used for wound management or the treatment of skin diseases. Keratinocyte-derived iPSCs have been reported to express endogenous pluripotent stem cell markers and to form derivatives of the three germ layers when injected into mice. With the appropriate in vitro conditions, they were found to resemble cholinergic motor neurons that were functionally active, based on the ability to generate action potentials. Importantly, these cells could be expanded without the support of feeder cells, a desirable quality if these cells are to be used for biomedical applications. 154
Mesenchymal stromal cells
MSCs are a type of adult, multipotent progenitor cells that can be readily expanded in culture. 155 These cells actively secrete a rich composite of bioactive factors that may prove to be a safe, easy-to-deliver therapy to improve tissue repair. 156 Human MSCs have been actively studied as a therapy for the healing of burn wounds, incisional wounds made by a scalpel during surgery, and excisional wounds generated by the removal of a portion of skin. 157 –160
On the NIH Clinical Trials website, there are currently several trials listed to test the efficacy of MSCs to promote the healing of diabetic foot wounds as well as burn wounds. Although human MSC therapy for wound, and other injuries and diseases, has been studied for many years, it still is not accepted in clinical practice. This is in contrast to equine medicine, where experimental and clinical studies of MSC therapy have led to the widespread clinical use of MSCs, mainly to improve orthopedic injuries. 161,162 As a standard practice, veterinarians isolate MSCs from the patient's bone marrow, expand the cells in culture, and then cryopreserve the cultured cells in the event the patient suffers a tendon or ligament injury later in life. If and when this happens, these autologous MSCs are revived and injected at the site of injury.
In vitro systems have been used to study the potential of equine MSCs derived from peripheral blood, a less invasive source than bone marrow, for wound management. It has been shown that these cells secrete factors that stimulate angiogenesis, secrete proteins that promote DF migration, and produce antimicrobial compounds that inhibit the growth of bacteria commonly found in skin wounds. 163 –165 It was also determined that when exposed to MSC-derived secreted factors, the expression of genes associated with a contractile cell phenotype is altered in EGT-derived DF, but not DF isolated from normal skin, and that these factors do not stimulate the migration of EGT-derived DF as they do for DF derived from normal skin. 99 Finally, methods of cell encapsulation that may facilitate the delivery of equine MSC-secreted factors to skin wounds have been explored. 105 Collectively, these in vitro studies suggest that factors secreted by equine MSCs could promote wound healing in vivo and may be used as a treatment option to prevent or reduce EGT on the lower limbs of horses.
A recent in vivo study from the group of Borjesson showed that injection of allogeneic MSCs into experimentally induced wounds on the distal limb altered gene expression at the wound site and enhanced early healing. 134 The promising results of this study provide a strong basis for additional in vivo studies designed to further evaluate the effects of MSC therapy on skin wounds of the horse.
Platelet concentrates
A fine-tuned balance of growth factors and cytokines is critical to successful wound healing. Because these signaling proteins are responsible for stimulating and directing the dynamic cell–cell and cell-ECM interactions that occur during healing, the composition and concentration of growth factors and cytokines vary both across the wound bed and temporally during the healing process. Since growth factors such as PDGF, VEGF, and EGF, and cytokines such as ILs, IFNs, lymphokines, and TNFs, are necessary for cutaneous wound healing, their therapeutic potential has been studied. Applying individual growth factors or cytokines to wounds, however, has not proven successful. This is not surprising since the wound bed contains a changing array of proteases, enzymes that may immediately destroy specific exogenous proteins, which can inhibit the action of these growth, and other, factors. Conversely, the application of individual growth factors or cytokines to wounds might flood the wound bed with overwhelming signals, which could disable resident cells and prevent them from participating in the healing process. 14
The administration of platelet concentrates, also called platelet-rich plasma (PRP), has been explored as an alternative to treating wounds with individual factors, in both human and equine medicine. Platelet concentrates are usually generated by centrifuging the patient's own blood, to collect the autologous platelet-rich fraction as well as blood cells that secrete an array of growth factors and cytokines. Human platelet concentrates of various compositions have been classified and characterized based on leukocyte and fibrin content in an attempt to identify features that add therapeutic value. 166 A recent review on PRP adjunct therapy in humans described that PRP (1) improved the healing of acute wounds, (2) had mild (or no) positive effects on wounds resulting from laser therapy, and (3) has not been studied in the context of burn wounds extensively enough to draw any conclusion for the treatment of this type of wounds. 167
In horses, several studies suggest that PRP has beneficial effects on the healing of surgical as well as burn wounds, 168,169 while it has been reported in other studies that topical application of PRP does not accelerate the repair of small granulating wounds on the distal limb of horses. 170 Clearly, more work is needed to determine what types of platelet concentrates are the most appropriate for the treatment of specific types of cutaneous wounds.
Maggot debridement therapy
For centuries, fly larvae have been purposefully used to debride necrotic tissue from human cutaneous wounds, but the efficacy and full benefits of larval therapy have not been thoroughly assessed or optimized. 171 The most commonly used flies belong to the family Calliphoridae, with Lucilia sericata, the greenbottle blowfly, being the most widely accepted species. Newly emerged, germ-free larvae can be purchased from FDA-approved vendors with a prescription for both human and veterinary use. 14
Historically, it has been accepted that maggots debride wounds, which prepares the wound bed for healing. More recent evidence suggests that larvae may contribute to wound healing in additional ways. In vitro studies have shown that maggot secretions can reduce biofilm 172,173 and digest components of necrotic tissue, 174 implying that they may serve to disinfect wounds. A retrospective analysis designed to describe and assess the results of maggot debridement therapy in horses concluded that, “maggot debridement therapy can be recommended in equids for debridement and enhanced healing and its potent antibacterial action.” 175 Similarly, a survey of veterinarians who used maggot debridement therapy on hoof and leg wounds of horses concluded that maggot therapy can serve as a useful treatment. 176
This therapy, especially until fully optimized and proven effective, may be rejected by people as it is considered to be esthetically unappealing by many. The horse is a reasonable model to refine maggot debridement therapy, to elucidate the mechanisms, whereby it improves wound healing, and to provide evidence that it can be a safe and effective therapy for both humans and horses.
Negative pressure therapy
In human medicine, negative-pressure wound therapy (NPWT) is used as an adjunct to traditional methods for treating a variety of cutaneous wound types. There are several types of NPWT systems, but all are similar in that they involve dressing the wound to create a sealed area that is then subjected to a vacuum. Drainage tubing is also incorporated to allow for the removal of fluids moved to the surface by the vacuum. 177 NPWT promotes second-intention healing, by reducing the wound size, removing excess fluid, and improving the wound environment in ways that are not fully understood, but are thought to include modulation of cytokine expression in the wound bed toward a profile that favors angiogenesis, ECM remodeling, and deposition of granulation tissue. 178,179
A recent and comprehensive review of NPWT concluded that it can be used to manage chronic, complex, and infected wounds, but that results from randomized control trials are variable. 177 Therefore, additional research focused on understanding the mechanisms of action of NPWT is needed to fully take advantage of this promising adjunctive therapy. Several case studies in horses have also concluded that NPWT facilitated second-intention healing and shortened the time to healing. 180 –182 However, and in line with human studies, the mechanisms through which NPWT acts in horses have not been defined. Clinical trials in horses could shed light on how NPWT alters the wound microenvironment, leading to more effective treatments for both humans and horses.
Engineered skin tissue substitutes
Skin substitutes, grown ex vivo, are commercially available for use in human patients. They are used when patients do not have adequate skin available for autografting and to reduce the risks of rejection and disease transmission that are associated with allografted skin. Engineered skin substitutes may be used to replace the dermal or epidermal layers and comprised various combinations of skin cells and ECM molecules.
A recent review of human skin substitutes determined that these products have revolutionized the management of wounds, but that more robust trials are needed to identify optimal skin substitutes for specific types of wounds, as well as to determine what features can be added to improve the intrinsic quality of currently available substitutes. 183 Veterinary researchers have developed equine cell culture systems to support the proliferation and differentiation of keratinocytes and to produce skin equivalents that consist of fibroblasts and an epidermal layer of differentiated keratinocytes, 115,120 but engineered skin substitutes are not currently used in equine clinical medicine.
Major limitations of skin substitutes for both species are that (1) they do not include organs such as glands, blood vessels, and nerves and (2) allogeneic skin substitutes do not incorporate into the wound bed, but rather serve as biologic dressings that may be rejected by the recipient's immune system. An ideal skin substitute would be generated from autologous stem cells, which could divide and differentiate into the various cell types found in skin, without risk of rejection.
SUMMARY
Healthy, uncompromised skin is required for both humans and horses to experience a high quality of life. Due to a lack of understanding of the mechanisms that contribute to normal and impaired cutaneous wound healing, current therapies are inadequate for both species. This poses a large burden to the human and veterinary health care systems not only economically but also physically, as a result of the pain and suffering experienced by human and equine patients with nonhealing wounds. The horse is a physiologically relevant large animal species that may serve as a model to define mechanisms involved in wound healing, and to test therapies for both humans and horses. We believe that an increase in basic scientific research and clinical studies focusing on skin wound healing in horses will produce data that will benefit both human and veterinary health.
TAKE-HOME MESSAGES
Cutaneous wounds are a problem in human and equine medicine.
Conventional methods for treating skin wounds are generic and have not changed in decades.
A better understanding of the mechanisms involved in both normal wound healing and the development of chronic nonhealing wounds has gradually led to the development of more specific and targeted therapies.
Relevant animal models are needed to study the mechanisms involved in wound healing and to test newly developed therapies.
The horse is a physiologically relevant large animal that may serve as an ideal model to study targeted processes involved in wound repair, as well as test novel therapies.
Studies performed in horses will benefit veterinary as well as human medicine.
Footnotes
Acknowledgments and Funding Sources
G.V.D. is supported by the Harry M. Zweig Memorial Fund for Equine Research at Cornell University and by the Agriculture and Food Research Initiative Competitive Grant no. 2018-67015-28309 from the UDSA National Institute of Food and Agriculture. C.L.T. is supported by Natural Sciences and Engineering Research Council of Canada (NSERC grant No. 250231).
Author Disclosure and Ghostwriting
The authors declare they have no competing interests. No ghostwriters were used to prepare this article.
About the Authors