
Abstract
Objective:
The healing of skin wounds is typified by a pattern of robust angiogenesis followed by vascular regression. Pigment epithelium-derived factor (PEDF), a recognized endogenous antiangiogenic protein, regulates vascular regression in resolving wounds through an unknown receptor. Among the multiple receptors for PEDF that have been identified, low-density lipoprotein receptor-related protein 6 (Lrp6) has been described as a regulator of angiogenesis in multiple systems. The purpose of the current study was to determine if the Lrp6 receptor plays a role in vessel regression in wounds.
Approach:
Excisional skin wounds were prepared on C57BL/6 mice. RT-PCR and immunoblots were performed to measure Lrp6 expression over a time course of wound healing. Immunohistochemistry was performed to localize Lrp6 in both recombinant PEDF (rPEDF)-treated and control wounds. To examine whether Lrp6 is critical to the regulation of capillary regression in vivo, wounds were treated with Lrp6 siRNA to minimize its presence in wounds. Immunohistochemistry for CD31 was performed to quantify blood vessel density.
Results:
PCR and immunoblots revealed significant increases in Lrp6 expression during the vascular regression phase of wound healing. Lrp6 was found to colocalize with CD31+ endothelial cells in wounds. The addition of rPEDF to wounds caused an increase in Lrp6-CD31+ endothelial cell colocalization. Inhibition of Lrp6 by siRNA impeded the vascular regression phase of healing.
Innovation:
This study is the first to demonstrate an association between Lrp6 and vessel regression in wound healing.
Conclusion:
Lrp6 is expressed in wounds in a temporal and spatial manner that suggests it may be a receptor for PEDF during vascular regression. PEDF increases Lrp6 expression in the wound vasculature, and inhibition of Lrp6 blocked vascular regression in wounds. The results suggest that Lrp6 is important to vascular regression in wounds, possibly through direct interaction with PEDF.
Luisa A. DiPietro, DDS, PhD

Introduction
During the wound healing process, an ordered sequence of capillary growth and regression takes place. Disruption of any aspect of this angiogenic process may lead to the development of healing disorders, and dysregulated angiogenesis has been described to play a role in impaired healing, such as that seen in diabetes and with aging. 1 –4 Many factors, including VEGF, promote angiogenesis in wounds, a process that is necessary for the restoration of oxygenation and nutrient support to injured tissue. 5,6 However, in wounds, vessel sprouting and branching first create an expansive array of immature and tortuous blood vessels. 7,8 Eventually, as healing proceeds, this immature network resolves by systematic pruning of vessels. 9 Multiple mediators, including CXC3 ligands, TSP1, and Sprouty 2, have been shown to play a role during the resolution phase of healing, and to regulate to vascular regression and stability. 10,11 More recent studies in our laboratory demonstrate that pigment epithelium-derived factor (PEDF) is likely to be the most significant mediator of vessel regression in wounds, and that PEDF provides the necessary signals required to remove immature capillaries. 12 Several receptors for PEDF have been identified, yet the distribution and function of these receptors among the cell types found in dermal wounds is unclear. 13 In this study, we focus on lipoprotein receptor-related protein 6 (Lrp6) as a candidate PEDF receptor in wounds, primarily due to its well-described ability to modulate Wnt signaling in processes, such as fibrosis, tumorigenesis, and angiogenesis, in other systems. 14 –16 Furthermore, mutations in Lrp6 have been shown to result in impairments in migration, proliferation, apoptosis, and Wnt signaling in human umbilical vein endothelial cells. 17,18
Clinical Problem Addressed
Angiogenesis is a hallmark of the proliferative phase of wound healing. Abnormal angiogenesis such as impaired angiogenesis in diabetic wounds 19 and excessive angiogenesis in hypertrophic scar or keloids 20 –23 contributes to the pathogenesis of chronic wound healing and scar formation, respectively. Understanding the roles and interactions of PEDF and Lrp6 in vessel regression during wound healing could provide a new therapeutic means to correct pathological angiogenesis.
Materials and Methods
Animal model
Eight-week old-female C57BL/6 mice (Harlan Laboratories, Inc., Indianapolis, IN) were subjected to standardized skin wounds 12 as previously described. Briefly, mice were anesthetized with ketamine (100 mg/kg) and xylazine (5 mg/kg), and two 3-mm-diameter full-thickness excisional wounds were made on the shaved dorsal skin using a biopsy punch (Acuderm, Inc., Ft. Lauderdale, FL). Wounds were harvested at multiple time points after injury; samples were stored in RNA later for PCR, snap frozen for immunoblot analysis, or embedded in optimal cutting temperature (OCT) compound for immunohistochemistry. All animal procedures were approved by the Institutional Animal Care and Use Committee of the University of Illinois at Chicago.
PEDF treatment
Topical recombinant PEDF (rPEDF) treatment was applied daily to each wound until day 10 postinjury as described in our previous study. 12 For the first 3 days, 2 μg of rPEDF in 10 μL 25% pluronic gel (Sigma-Aldrich, St. Louis, MO) was directly applied to the surface of each wound. From day 4 to 10, 2 μg rPEDF in 10 μL phosphate-buffered saline (PBS) was injected intradermally into each wound. Control wounds were treated with PBS. Wounds were harvested at day 10 postinjury and placed in OCT compound for immunohistochemistry.
Protein extraction and Western blot analysis
Frozen 3-mm-diameter wound samples were homogenized in 500 μL of radioimmunoprecipitation assay buffer (Thermo Fisher Scientific, Waltham, MA), including a protease inhibitor cocktail (1/100 dilution; Sigma-Aldrich). Centrifugation of samples was performed for 15 min at 13,000 rpm at 4°C. Supernatants were collected, and protein concentrations were quantified using a BCA Protein Assay Kit (Pierce, Rockford, IL). The protein extracts were combined with SDS-PAGE buffer and 2-mercaptoethanol (5% of total volume), and boiled for 3 min. One hundred twenty micrograms of each protein sample was loaded into a 10% Tris-glycine acrylamide gel (Bio-Rad, Hercules, CA) and separated using a Mini-Protean Electrophoresis System (Bio-Rad). Proteins were transferred to a nitrocellulose membrane using a standard protocol. 10 Membranes were blocked by treatment with 5% skim milk powder in Tris-buffered saline for 1 h, then incubated with primary antibodies of either rabbit anti-mouse Lrp6 (1/1000; Abcam, Cambridge, UK) and rabbit anti-β-actin (1:1000 dilution; Abcam) for 1 h at room temperature. After washing with Tris-buffered saline/Tween 20, membranes were incubated with goat anti-rabbit horseradish peroxidase (1/2,000; Bio-Rad). Enhanced chemiluminescence was used for the detection of positive bands. Images were acquired and relative protein quantification was determined using a ChemiDoc system (Bio-Rad).
In vitro Lrp6 siRNA treatment
Primary mouse skin dermal endothelial cells (Cell Biologics, Chicago, IL), primary mouse skin dermal fibroblasts, and an immortalized mouse skin keratinocyte cell line, PAM212, (kindly provided by Dr. Jonathan Jones, Washington State University), were cultured in endothelial cell medium (Cell Biologics), Dulbecco's modified eagle medium with 10% fetal bovine serum (FBS), and minimal essential medium with 10% FBS, respectively. Cells were cultured in 12-well plates until cell density reached 60–80% confluency, and then transfected with Lrp6 siRNA or scrambled siRNA control (Thermo Fisher Scientific) using lipofectamine RNAiMAX transfection reagent (Thermo Fisher Scientific) following the manufacturer's instructions. Forty-eight hours later, transfected cells were harvested and RNA was prepared for real-time PCR as described below.
In vivo Lrp6 siRNA treatment of mouse skin wounds
Lrp6 siRNA–Invivofectamine 2.0 complexes were prepared according to the manufacturer's instructions (Thermo Fisher Scientific). Briefly, 500 μL of 3 mg/mL Lrp6 siRNA or control siRNA (as described above) was combined with 500 μL complexation buffer, and mixed with 1 mL Invivofectamine 2.0 reagent by vortexing. The mixture was incubated at 50°C for 30 min. The Lrp6 siRNA–Invivofectamine complex was then dialyzed in PBS using an 8–10 kDa molecular weight cutoff Float-A-Lyzer G2 dialysis device (Spectrum Laboratories, Inc., CA) for 2 h at room temperature. The final concentration of Lrp6 siRNA was 750 μg/mL. The complexed siRNA was stored at 4°C for no longer than 1 week before use. Twenty microliters (15 μg) Lrp6 or control (scrambled) siRNA–Invivofectamine 2.0 complexes were applied topically onto the open wound immediately after injury, on day 1, and on day 2. From day 3 postinjury onward, 20 μL (15 μg) Lrp6 siRNA–Invivofectamine (Thermo Fisher Scientific) or control was administered into each wound through intradermal injection.
Real-time PCR
Wounds containing 0.5–1.0 mm of the wound edge were collected at multiple time points. Wound samples were homogenized in TRIzol (Invitrogen, Carlsbad, CA) using a Power Gen 125 homogenizer (Fisher Scientific), and total RNA was then extracted according to the manufacturer's instructions. Total RNA from cultured cells was also extracted using TRIzol but without homogenization. One microgram of each RNA sample was treated with DNAse I (Invitrogen), and subjected to reverse transcription using a RETROscript Kit (Invitrogen). Relative mRNA expression of Lrp6 was examined using a StepOne Plus real-time PCR system (Applied Biosystems, Carlsbad, CA) that employs SYBR Green PCR mix (Roche, Basel, Switzerland). Gapdh was used as a housekeeping gene for calibration. Primer sequences are: Gapdh: forward: 5′-TCACCACCATGGAGAAGGC-3′, reverse: 5′-GCTAAGCAGTTGGTGGTGCA-3′; Lrp6: forward: 5′-AGCCCTATGATCTCAGCATTG-3′, reverse: 5′-TCGCCTTTTAGAACCACTCC-3′.
Indirect immunofluorescence
Eight-micrometers-thick frozen sections were prepared from wound tissues embedded in OCT compound, air dried, and fixed in cold acetone for 10 min. Sections were blocked with 10% goat serum in PBS for 30 min. Sections were incubated with rat anti-mouse CD31 (1/1600 dilution; BD Biosciences, San Jose, CA) and rabbit anti-mouse Lrp6 (1:200 dilution) (Abcam) for 45 min. After washing, the sections were incubated with Alexa 594 goat anti-rat IgG (Invitrogen) and Alexa 488 goat anti-rabbit IgG (Invitrogen) for 45 min. All staining procedures were performed at room temperature. Stained sections were observed using a fluorescence microscope (Axioskop 40; ZEISS, Oberkochen, Germany), and fields were captured to cover nearly the entire wound bed. For each section, two to five fields per section were analyzed and averaged to produce a unique value for each animal. No overlapping images were used for the quantification; only unique areas were counted. Images were recorded with a digital camera (AxioCam MRc; ZEISS). The area within the wound bed and the percentage of endothelial cells that were positive for Lrp6 were measured using ImageJ. The vessel density was quantified by determining the percent of CD31-positive area in the wound bed.
Results
Lrp6 mRNA and protein expression increases in wounds during the time of vascular regression
The expression pattern of Lrp6 was determined over the time course of wound healing and compared with the known and previously published pattern of angiogenesis and vascular regression in the 3 mm excisional wound model used in this study. 12 Lrp6 mRNA expression levels were significantly increased at days 7 to 21, time points that represent the periods when vascular regression occurs and PEDF levels are high (Fig. 1A). 12 Immunoblot analysis demonstrated that Lrp6 protein expression increased in a pattern generally similar to Lrp6 mRNA. Significant differences were observed between day 7 and 14 (p < 0.01), day 10 and 14 (p < 0.01), and day 14 and 21 (p < 0.05) (Fig. 1B, C).
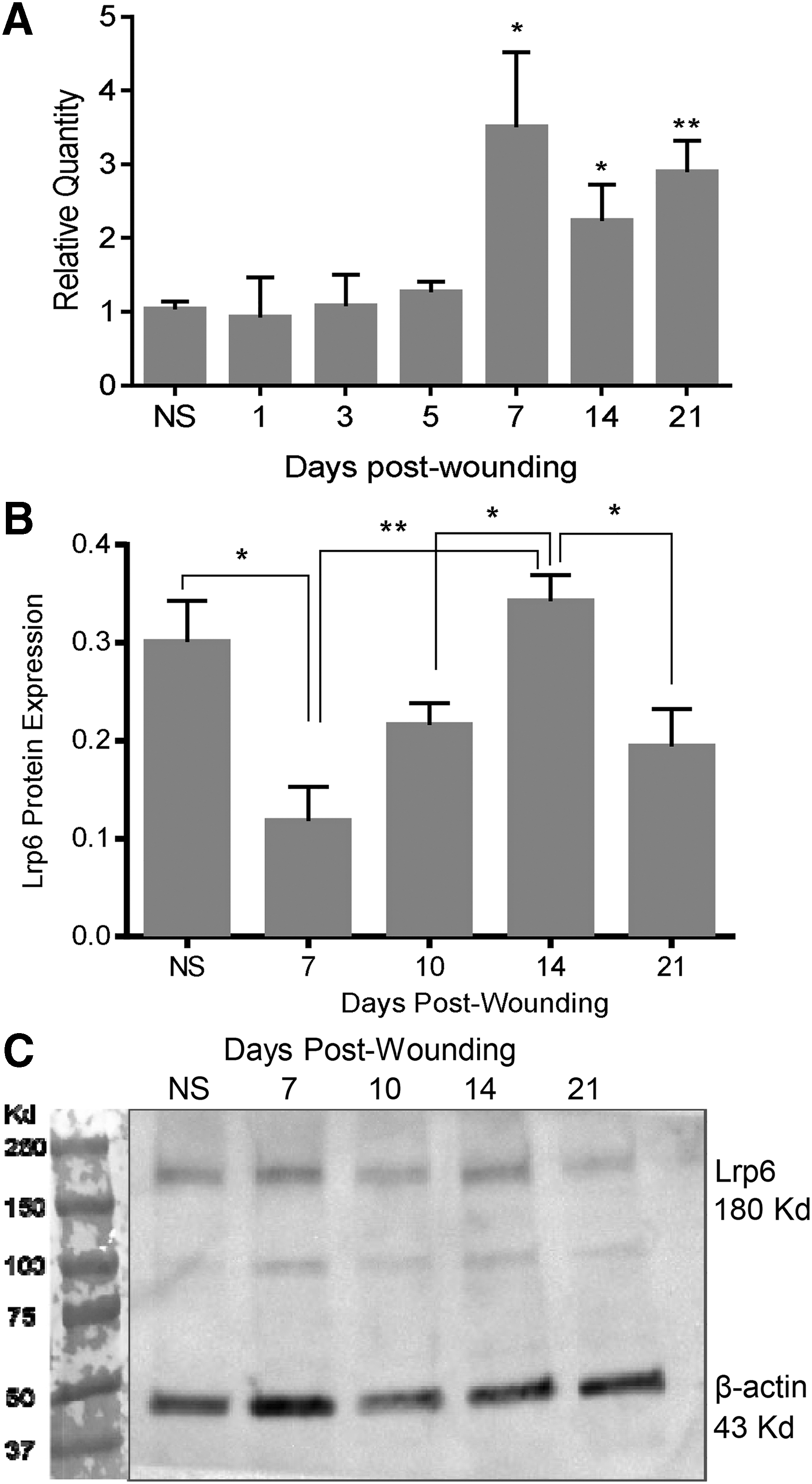
Lrp6 expression in skin wounds.
Colocalization of Lrp6 and blood vessel increases during the remodeling phase
To determine whether Lrp6 is expressed on endothelial cells in wounds, blood vessels were examined in histological sections by image analysis following CD31 and Lrp6 immunohistochemical staining. Colocalization of CD31 and Lrp6 increased significantly over the time course of healing and reached peak levels at day 21, during the remodeling phase of wound healing (Fig. 2A, normal skin vs. day 21, p < 0.01, day 7 vs. day 21, p < 0.05, day 14 vs. day 21, p < 0.01). Wounds that were treated with recombinant PEDF demonstrated a statistically significant increase in colocalization between Lrp6 and CD31 at day 10 (p = 0.05) (Fig. 2A, B).
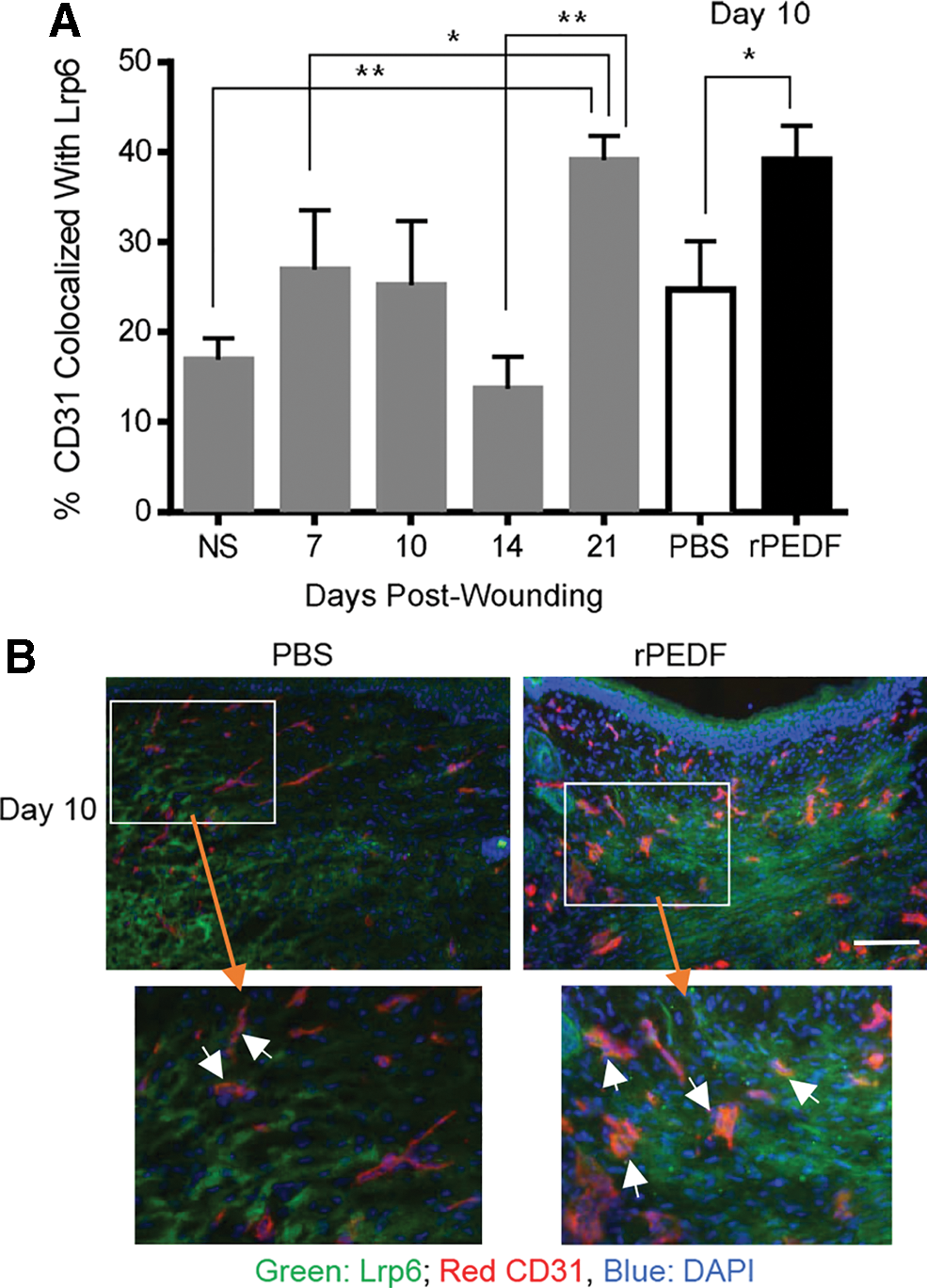
Colocalization of Lrp6 and CD31 in skin, control wounds, and in wounds treated with rPEDF. Immunohistochemical detection of Lrp6 and the EC marker CD31 was performed on histological sections.
siRNA treatment of endothelial cells, fibroblasts, and keratinocytes effectively inhibits Lrp6 expression in vitro
To determine whether siRNA approach could be used to inhibit Lrp6 production in wounds, the effectiveness of Lrp6 siRNA was first tested in vitro. Transfection of Lrp6 siRNA in vitro caused a trend toward decreased Lrp6 mRNA expression in endothelial cells (p = 0.17), and a significant decrease in Lrp6 mRNA in fibroblasts (p < 0.0001) and keratinocytes (p < 0.0001) as compared with treatment with control media or a scrambled siRNA (Fig. 3A).

Blockade of Lrp6 production by siRNA treatment is effective in vitro and in vivo.
siRNA treatment of wounds inhibits Lrp6 expression in vivo and results in increased capillary content in the resolving wound
To test the role of Lrp6 in vascular regression in wounds, Lrp6 was inhibited by treatment of wounds with Lrp6 siRNA. To confirm the effectiveness of the Lrp6 siRNA, semiquantitative real-time PCR was performed. Treatment of wounds with Lrp6 siRNA led to a significant decrease in the level of Lrp6 mRNA in wounds (Fig. 3B). Functionally, siRNA knockdown of Lrp6 in vivo resulted in a significant increase in day 10 wound capillary density as compared with the control scrambled siRNA treatment (p = 0.01) (Fig. 3C, D). The results demonstrate that a decrease in Lrp6 levels leads to higher capillary content during the vascular regression phase of wound healing, suggesting that Lrp6 plays a role in vascular regression.
Discussion
While many prior studies have examined the factors that lead to capillary growth in wounds, the process of capillary pruning during resolution has received less study. By examining a panel of candidate antiangiogenic factors in wounds, we previously demonstrated that the endogenous antiangiogenic factor PEDF is the predominant mediator of capillary regression in healing wounds. 12 These prior studies also demonstrated that PEDF colocalizes with endothelial cells in the healing wound, and that the level of colocalization increases as remodeling and capillary refinement occur. These findings prompted us to examine the role of Lrp6, a known PEDF receptor, in the wound healing process.
Several well-described receptors for PEDF exist, including PEDFR, Lrp6, and the laminin receptor. 16,24 –26 PEDFR, also known as patatin-like phospholipase domain-containing two protein, 25 is a lipase-linked cell membrane protein that causes the plasma membrane to release free fatty acids and lysophosphatidic acids. 13 PEDFR increases gamma-secretase activity, an action that can lead to the cleavage of the VEGF receptor 1. 26,27 Since wound angiogenesis relies heavily upon VEGFR2, 27 a role for PEDFR in wounds seemed unlikely. In contrast to PEDFR, Lrp6 has been described as a regulator of angiogenesis in several systems other than healing wounds. 17,18 In models such as the retina, and in developmental systems in both frog and mouse, antiangiogenic proteins such as PEDF have been shown to bind Lrp6 and subsequently block Wnt signaling. 13,14 This interaction ultimately halts the process of angiogenesis and regulates blood vessel density. 28 Wnt ligand-induced Lrp6-Frizzled receptor dimerization is a key aspect of Wnt signaling, and importantly, interaction between PEDF and Lrp6 blocks this event, thus disrupting angiogenesis. 15 Given the known role of Lrp6 as a modulator of angiogenesis, as well as previous studies that show that the Wnt pathway plays a role in the wound healing process, Lrp6 seems a likely candidate PEDF receptor in healing wounds. 15
The current study supports the idea that Lrp6 is important in the pruning of the vasculature in resolving wounds, and suggests that Lrp6 may serve as a PEDF receptor in this context. Similar to PEDF, Lrp6 was found to colocalize with endothelial cells in wounds. The colocalization pattern of Lrp6 and blood vessels in wounds was in fact quite parallel to what was seen previously for PEDF and blood vessels. 12 The highest level of colocalization was seen at day 21, a time when blood vessel regression and vessel maturation are prominent features. Treatment of wounds with rPEDF intensified the levels of Lrp6 in wounds, suggesting a feedback mechanism by which PEDF stimulates Lrp6 expression. Finally, inhibition of Lrp6 expression was shown to impair vascular regression in wounds, signifying that Lrp6 is functional during the process of capillary regression in dermal wounds.
The studies here focus on a role for Lrp6 as a potential endothelial cell receptor for PEDF during wound healing. The results here show a clear distribution of Lrp6 on endothelial cells during vessel regression in the phase of wound healing remodeling. A limitation of our studies is that we do not have direct in vivo evidence for an interaction between Lrp6 and PEDF. A second limitation of our study involves the distribution of Lrp6 in the wounds. Previous studies, including our own, have demonstrated that PEDF can affect other cell types within the wound, such as keratinocytes and fibroblasts. 29 –31 Because Lrp6 is produced by multiple cell types, the inhibition of Lrp6 in the wound might include cells other than endothelial cells. Nevertheless, the presence of Lrp6 on endothelial cells, along with the change in vascular regression when Lrp6 production was inhibited, both strongly support the idea that one function of Lrp6 in wounds involves its activation during vascular pruning.
Conclusions
The results provided here demonstrate that Lrp6 is expressed in wounds in a temporal and spatial manner that aligns with its role as a possible receptor for PEDF during vascular regression. The results further show that rPEDF increases Lrp6 expression in wound vasculature, a response that would enhance endothelial responsiveness to PEDF. Inhibition of Lrp6 in vivo led to impaired capillary regression in wounds, thereby demonstrating that Lrp6 plays a role in the vascular regression process. These studies show that, similar to their ligands, the expression of the receptors for proangiogenic and antiangiogenic molecules may modulate during wound healing. These results suggest that the development of effective proangiogenic or antiangiogenic strategies must consider both ligand and receptor status in tissues. This concept may have substantial impact on the development of PEDF as a therapeutic for angiogenesis-related diseases such as scar formation, fibrosis, and malignancy.
Innovation
PEDF has been described as an important endogenous regulator of angiogenesis in several tissues, including the eye and the ovary. 32 –34 Given its potent antiangiogenic effects, PEDF is currently under investigation as a therapeutic agent for diseases characterized by active angiogenesis such as malignancy and ocular disease. 13,35,36 Recent data suggest that the formation of scars and keloids is also preceded by a period of active angiogenesis, with some studies demonstrating that a reduction in angiogenesis reduces scar formation. 4,5 Thus the potential for the use of PEDF as a therapeutic is broad. The current finding suggests that any effective antiangiogenic strategy will need to consider whether adequate expression of functional receptors also occurs in the targeted tissue.
Key Findings
Lrp6 is expressed in wounds in a temporal and spatial manner that aligns with its ligand PEDF during vascular regression.
PEDF increases Lrp6 expression in wound vasculature.
Inhibition of Lrp6 in vivo impeded capillary regression in wounds. The results suggest that effective proangiogenic or antiangiogenic strategies must consider both ligand and receptor status in tissues.
Footnotes
Acknowledgments and Funding Sources
This work was supported by National Institutes of Health Grants T32-DE018381 (to LAD, ERM), R01-GM-50875 (to LAD), F30-DE024683 (to ERM). The authors would like to thank Wendy Cerny, David Fine, and Yan Zhao for their help and support with our studies.
Author Disclosure and Ghostwriting
No competing financial interests exist. The content of this article was written by the authors listed. No ghostwriters were involved.
About the Authors