
Abstract
Significance:
Chronic wounds, such as diabetic foot ulcers, venous stasis ulcers, and pressure ulcers affect millions of Americans each year, and disproportionately afflict our increasingly older population. Older individuals are predisposed to wound infection, repeated trauma, and the development of chronic wounds. However, a complete understanding of how the attributes of aging skin affect the wound healing process has remained elusive.
Recent Advances:
A variety of studies have demonstrated that the dermal matrix becomes thinner, increasingly crosslinked, and fragmented with advanced age. These structural changes, as well as an increase in cell senescence, result in altered collagen fiber remodeling and increased stiffness. Studies combining mechanical testing with advanced imaging techniques are providing new insights into the relationships between these age-related changes. Emerging research into the mechanobiology of aging and the wound healing process indicate that the altered mechanical environment of aged skin may have a significant effect on age-related delays in healing.
Critical Issues:
The interpretation and synthesis of clinical studies is confounded by the effects of common comorbidities that also contribute to the development of chronic wounds. A lack of quantitative biomarkers of wound healing and age-related changes makes understanding structure–function relationships during the wound healing process challenging.
Future Directions:
Additional work is needed to establish quantitative and mechanistic relationships among age-related changes in the skin microstructure, mechanical function, and the cellular responses to wound healing.

Scope and Significance
The structural and mechanical changes that occur with increasing age can have a significant effect on the susceptibility of skin to injury and the ensuing wound healing process. 1 This review highlights imaging modalities and mechanical testing methods used to quantify extracellular changes in the skin that occur with age, and how they relate to altered mechanical properties. We focus on how these structural and mechanical changes to the skin relate to each other and impact cellular responses during wound healing.
Translational Relevance
With advanced age, changes to the skin microstructure and mechanical microenvironment result in decreased cell sensitivity to mechanical stimulus and impaired healing outcomes. 2 This review seeks to relate knowledge of the biological and mechanical changes in aged skin to observed complications and delays in the healing process. By providing context into the mechanisms of delayed healing in aged skin, this information can help guide the development of new therapies and treatment strategies aimed at improving healing outcomes and preventing ulceration.
Clinical Relevance
The biological effects of intrinsic aging alter the mechanical environment of the skin and predispose it to delays and/or impairment during wound healing. Reduced collagen deposition and increased crosslinking in aged skin affect cell signaling in older skin, which results in prolonged inflammation, delayed migration/proliferation, and insufficient remodeling. Due to the prevalence of other comorbidities, key questions remain in understanding how intrinsic aging can lead to poor wound healing outcomes. It is critical to develop new quantitative tools to elucidate the relationships between the mechanical properties of aged skin and healing outcomes.
Overview
Through clinical and basic science research, many structural, compositional, and mechanical changes to the skin have been described as a function of age over the last century. 3,4 However, the relationship among these changes and their effect on the wound healing process are not well established. Skin aging is a combination of biochemical, mechanical, and environmental changes that accumulate over time. 5,6 These age-related changes result in clear visual changes and functional deficits that make the skin more susceptible to injury and can result in delays or impairments in the healing process. 7 As a result, older individuals are predisposed to wound infection, trauma, and the development of chronic wounds.
Although the sun, and other radiation sources, can produce significant damage and result in skin photoaging, this review will focus primarily on structural and mechanical changes resulting from intrinsic aging. The goal of this review is to integrate these reports of age-related changes to the skin and highlight emerging research on structure–function relationships in the skin using advanced imaging techniques and mechanical testing. We first provide a description of the structural changes that occur in the skin with age, with relevant background on the various imaging modalities used to characterize skin microstructure. We then discuss the mechanical properties of the skin and how they change with age, along with current testing methods used to derive these material properties. Finally, we describe how these age-related effects may influence the wound healing process.
Age-Related Structural Changes to the Skin
Epidermis
Structural changes throughout the epidermis from the dermal/epidermal junction (DEJ) to the stratum corneum (SC) occur with aging. At the DEJ, keratinocytes in the basement membrane of the epidermis project small, cytoplasmic villi into the upper layers of the dermis. 8,9 The DEJ also consists of dermal papillae that extend up from the dermis, and rete ridges that extend down from the epidermis, and range in size from 26 to 62 μm. 10 With increasing age, the villi of the epidermal membrane retract, causing the papillae and ridges to flatten with age. 8 Reflectance confocal microscopy (RCM) has been utilized to produce 3D images of the DEJ in vivo, as well as characterize the dermal papillae and rete ridges. 10 Through RCM, smaller rete ridge height is observed in aged skin, but no changes in the density of ridges is noted between young and aged skin. 10
The rete ridges contain a population of highly proliferative keratinocytes in their tips not seen in other areas of the stratum basale (SB), which provide structural support at the DEJ. 11 The keratinocytes are reported to have a diminished proliferative potential in aged skin, but the migration of the keratinocytes from the SB up through the SC remains unchanged. 9 The composition of the basement membrane also changes with age, with collagen IV, collagen VII, collagen XVII, integrin β4, and laminin-332, all decreasing over time. 12
The epidermis becomes thinner with advanced age, ranging from 35 ± 10 μm in young skin to 25 ± 8.7 μm in aged skin; although the adult SC is on average 30% thicker than young skin. 13 –15 (Fig. 1). The organization of the SC also changes with age. In young skin, corneocytes form a relatively consistent network of honeycomb-shaped polygons that establish the lines and furrows on the surface. 15 The furrows, or spaces between the keratinocytes, deepen with age and contribute to the aged look of skin. The primary furrows of the skin (>60 μm) increase in depth with age, whereas the secondary furrows (<60 μm), which are related to Langer's lines, decrease in number. 16
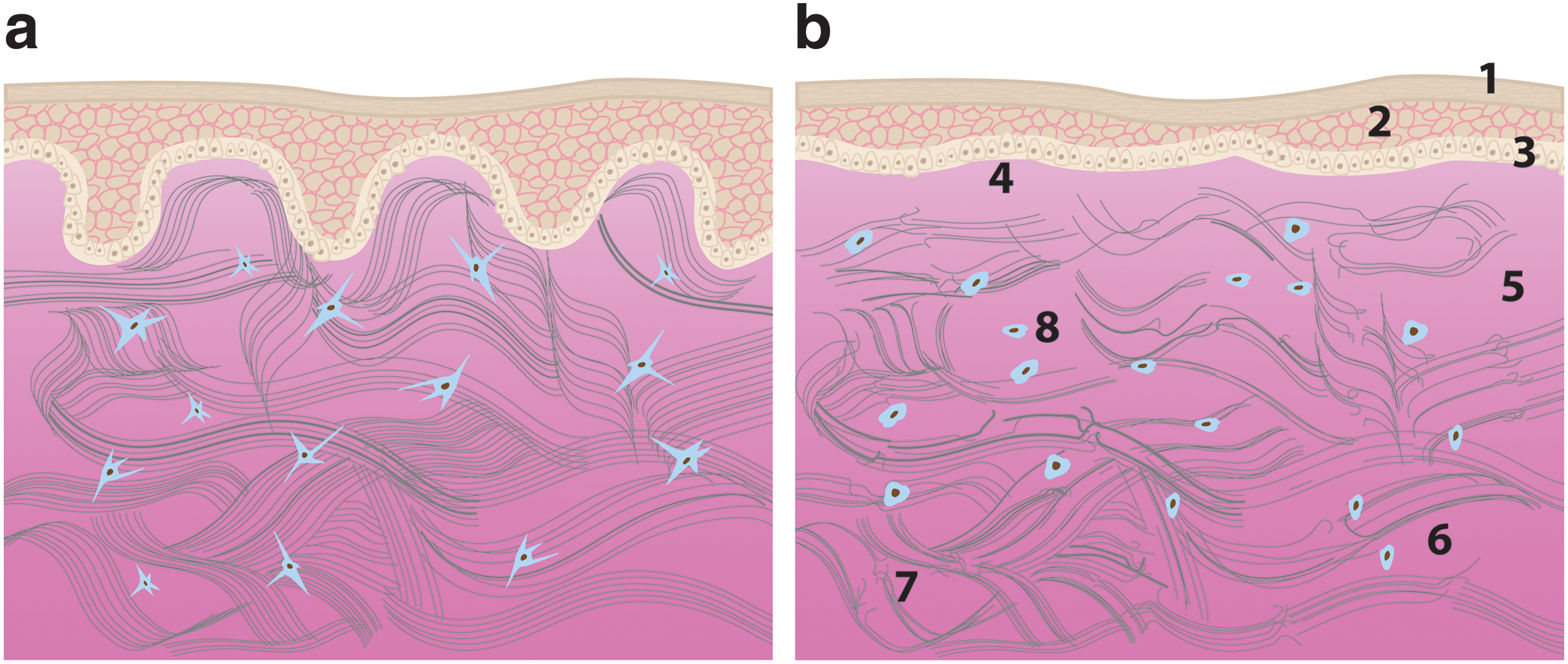
Schematic of skin sections in the young
As the skin ages, UV light imaging reveals that the surface topography becomes more irregular as surface corneocytes grow larger and the average size of a polygon in the network increases with age. 17 The lines that form these polygons are external structures that encircle the underlying dermal papillae. 17 As the papillae flatten with advanced age, the surface polygons expand such that the network no longer corresponds to the underlying papillae structure. The polygon density is not strongly correlated to the roughness of the skin, which also increases with age. 17
Dermis
The composition and organization of the dermis also undergo substantial changes with age, which results in altered physiological and mechanical function. The dermis consists largely of an extracellular matrix (ECM) composed of 70–90% type I collagen in adults, with the remaining components including elastin fibers, proteoglycans, and hyaluronic acid (HA). 18 The amount of collagen, normalized by skin surface area, decreases with age. 19,20 Through image analysis of the volume fraction of collagen from stained skin sections, it has been reported that collagen occupies 69% of the space in the papillary dermis, but this fraction decreases to 46% with advanced age. 21 Similarly, the collagen density decreases from 81% in young skin to 58% in aged skin in the reticular dermis. 21
In addition to changes in collagen density, the reticular dermal thickness is reported to double in thickness from birth to age 50, but then decrease at an even greater rate after age 50. 21 The proportion of collagen types also changes with age. Type III collagen is present near the DEJ, and in mouse skin (MF1-h hairless albino) the amount of type III collagen per total collagen content shows a steady increase with age. 20,22 Full abdominal skin samples from 30 human subjects reveal similar changes in collagen type with age, with 18–21.5% type III collagen in young to middle-aged donors and 19–23% in older individuals with some subjects showing as much as 31% type III collagen. 22
These changes in ECM composition may in part be the result of changes in cell–matrix interactions. 23 The number of focal adhesion sites, which help transmit mechanical forces and signaling between the ECM and cells, decrease with age. This decrease in adhesion sites limits the ability of the fibroblasts to interact with the ECM, and aging fibroblasts are known to have a decreased ability to migrate on their surrounding collagen network 23 (Fig. 1). The lack of focal adhesion sites and age-related changes to the cytoskeletal organization of fibroblasts is associated with decreases in collagen production. 24 This lack of mechanotransduction can stimulate the production of matrix metalloproteinases (MMPs) by the fibroblasts, which in turn degrade collagen fibers.
While MMP levels are nearly undetectable in young skin, MMP-1 shows a significant increase in expression with age. 25 Along with an increase of MMPs with age, levels of tissue inhibitors of matrix metalloproteinases (TIMPs) are not elevated with age, allowing further accumulation of MMPs and an increase in collagen matrix degradation. 25 Collectively, the decrease in collagen production from fibroblasts, the increase in MMP expression, and the unchanged TIMP levels result in a perpetuating degradation cycle leading to changes in matrix composition and organization in aged skin.
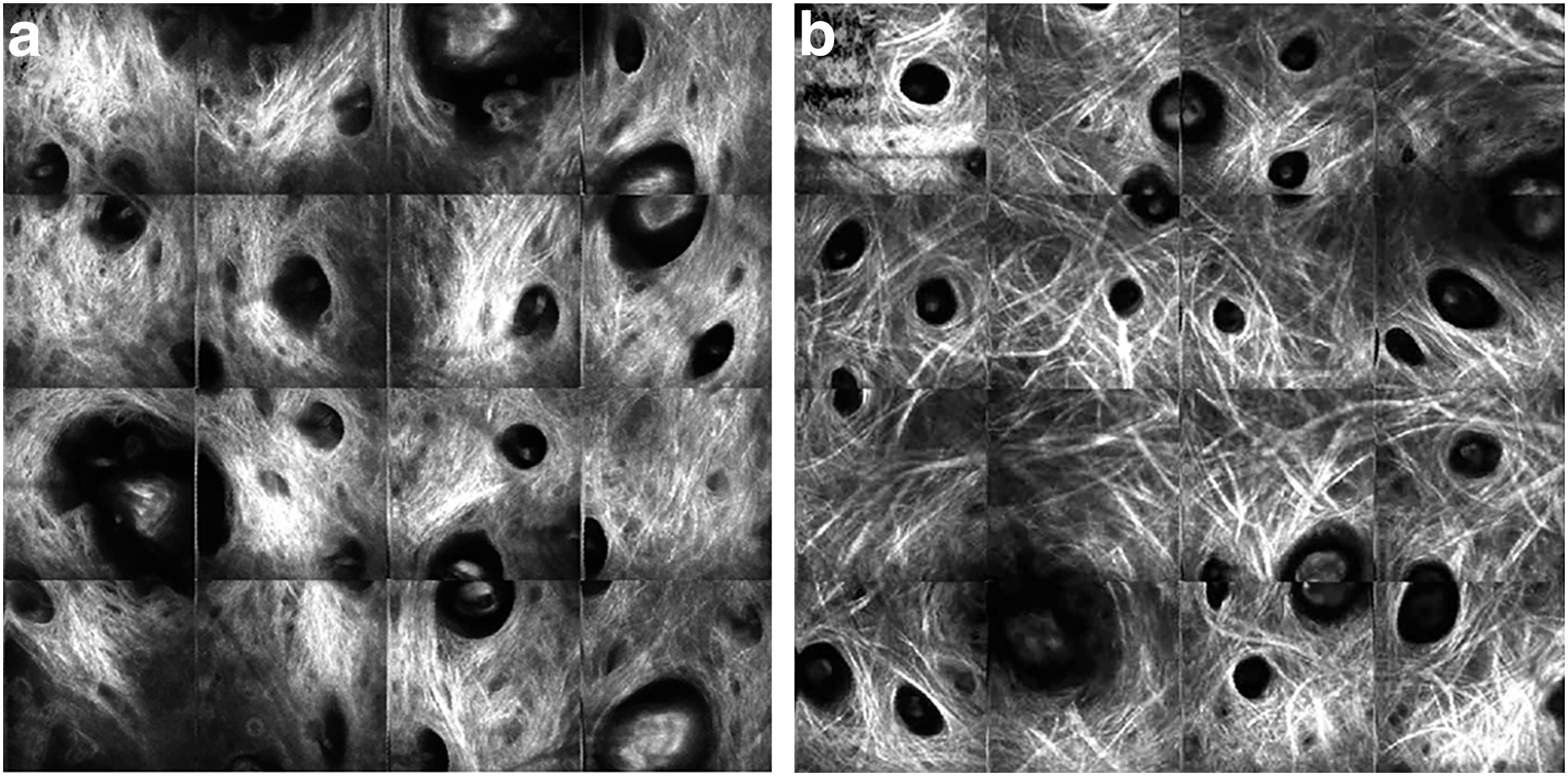
Altered dermal collagen organization with age has been quantified using a variety of imaging techniques. At the nanoscale, scanning electron microscopy revealed a more fragmented, clustered ultrastructure in aged skin (Fig. 2). 25 At the microscale, the thickness of collagen bundles from human abdominal skin were measured from Masson's trichrome-stained sections of skin, and the average fiber diameter from the papillary dermis was found to decrease with age, ranging from 1.1 μm down to 0.65 μm. 21 In the reticular dermis, the thickness of the collagen bundles increased with age until 45 years old, and then decreased with advanced age. Nondestructive imaging methods characterizing organization at a different length scale also show signs of altered collagen organization with age. In vivo RCM was used to compare young and old collagen networks, and older patients were found to possess increasingly coarse fiber bundles, and in some cases the fibers were found to curl. 15

SEM images of the dermis reveal long fibrils of intertwined collagen The network in young skin
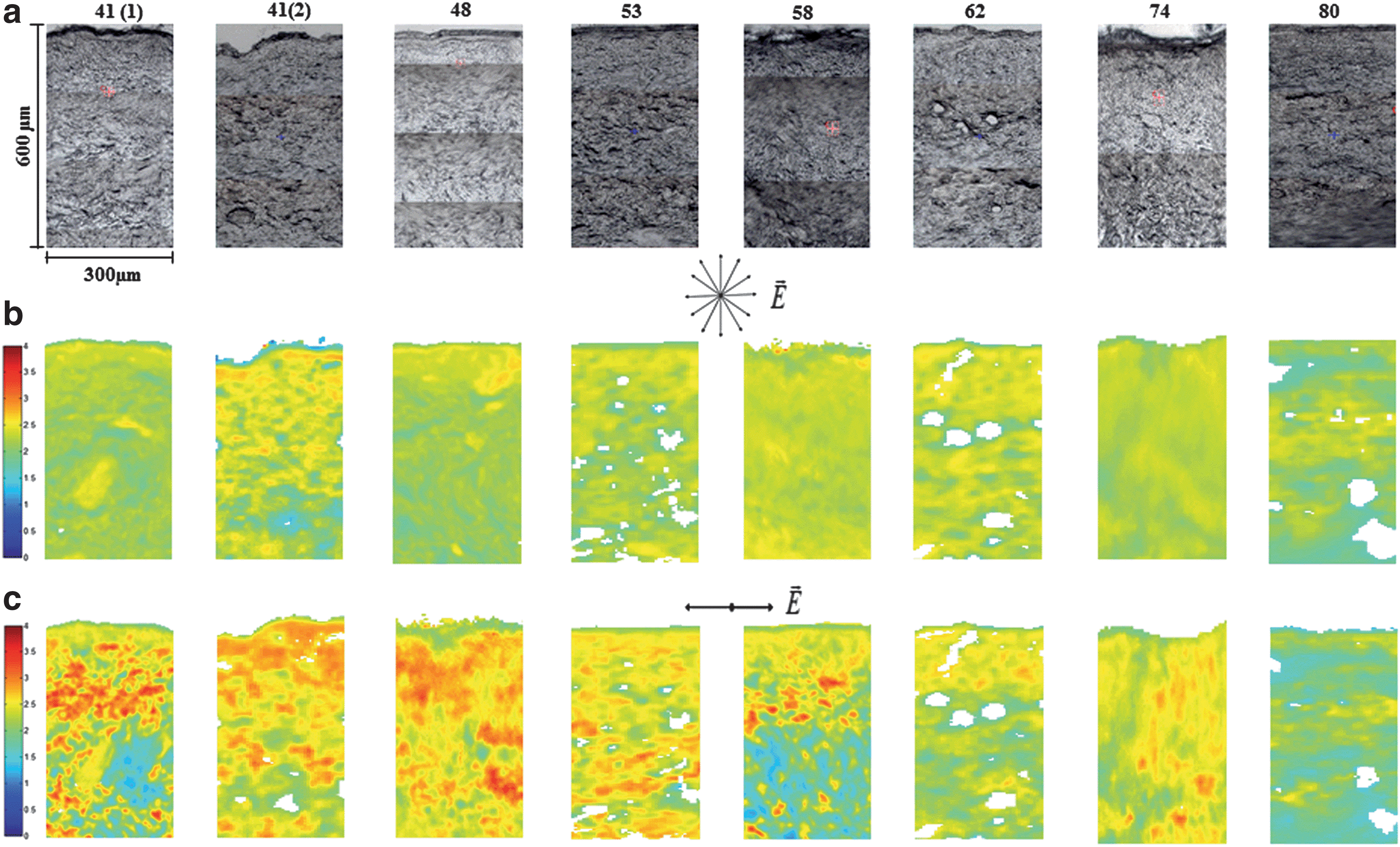
By exploiting the noncentrosymmetric nature of collagen molecules, second-harmonic generation (SHG) has also emerged as a powerful tool to image dermal collagen organization in vivo. SHG images with a 2.4 mm field of view of human cheek skin demonstrated a difference in collagen structure, with thicker, coarser collagen bundles with age (Fig. 3). 26 SHG images from Kunming mouse skin were also analyzed using a Fourier transform approach and provided a measure of collagen orientation, which indicated that fibers become more aligned with age. 27 In a separate study, Fourier transform infrared spectroscopy (FTIR) maps were acquired from human skin sections from women spanning 41–80 years of age (Fig. 4). Through the use of polarized light, collagen fibers oriented perpendicular to the skin surface could be isolated (Fig. 4c) and these polarized FTIR maps revealed that fiber orientation becomes more parallel to the skin surface during aging. 28
Multiple SHG images (each at 2.4 × 2.4 mm) of the collagen network from the cheek skin of a 20 year old
Skin from donors of different ages
Collagen crosslinking may have a significant effect on the observed changes to the organization of the collagen network with advanced age. Crosslinked collagen is incredibly stable, and can last ∼30 years before replacement. 29 Crosslinking begins as early as the developmental stages when collagen is stabilized by immature, enzymatic connections. 30 Lysyl oxidase (LOX)-mediated enzymatic crosslinks do not degrade easily; Histidinohydroxylysinonorleucine, a LOX-derived enzymatic crosslink, has been reported to increase more than six fold in the skin of both cows and humans over the first 6 years and six decades of life, respectively. 31 However, the relative proportion of LOX-mediated crosslinks are reported to decline with advanced age due to an even greater accumulation of nonenzymatic crosslinks. 18,30,32 Nonenzymatic crosslinks are the result of glycation reactions with collagen side chains and form advanced glycation end products. Because these crosslinks cannot be broken down as easily as the collagen fibrils themselves, it has been suggested that crosslinked collagen fragments accumulate over time and contribute to the aged appearance of skin. 29 In fact, fragmented collagen has been shown to increase over 400% in aged individuals (>80 years old) compared with young individuals (21–30 years old). 25 Nonenzymatic collagen crosslinks are autofluorescent, and emerging imaging modalities such as multiphoton microscopy (MPM) and fluorescence lifetime imaging (FLIM) can allow for the nondestructive quantification or classification of crosslink accumulation with time (Fig. 5). 33,34

Image of young
Hypodermis
Located below the dermis and above the underlying muscle, the hypodermis plays a key role in thermoregulation, provides a reservoir of progenitor cells, and is involved in key paracrine and endocrine signaling. 35 –37 The hypodermis is comprised primarily of adipose tissue, which undergoes substantial conformational changes with age. 35,38 There is a significant redistribution of subcutaneous fat in the body with increasing age, but overall subcutaneous white adipose tissue (sWAT) volume decreases with age, while visceral WAT volume increases. 37,39,40 This progressive loss of subcutaneous fat plays a role in reduced skin tension with age and the appearance of common age-associated features such as skin sagging and wrinkling. 39,41 –43 More importantly, subcutaneous fat helps to shield the skin from high mechanical stresses concentrated over bony tissues, such as the feet. The loss of subcutaneous fat with age, therefore, makes the skin of elderly patients more prone to high mechanical pressures, local tissue ischemia, and ulcer formation. 44,45
Although typically CT and MRI have been utilized to understand the dynamics of subcutaneous adipose tissue composition during the aging process, 39,46,47 coherent antistokes Raman scattering (CARS) is a promising MPM technique that can be combined with collagen SHG imaging for sWAT characterization at the microscale. 48 CARS imaging takes advantage of the molecular vibrations from C-H bond stretching and demonstrates a robust ability to resolve lipids and adipocytes without exogenous dyes in both in vitro and in vivo models. 48 –50
The Mechanical Function of Skin and its Relation to Microstructure
As described in the previous sections, intrinsic aging results in many changes to the biological structure and function of the skin, which in turn influences the mechanical properties of the tissue. Mechanical measurements of skin have been conducted using multiple testing methods, sampling sites, specimen species, and sample ages. Tensile stretching, indentation, suction, and torsional testing have all been used to characterize age-related changes in the mechanical properties of the skin. Additionally, imaging technologies (e.g., ultrasound elastography, optical coherence tomography) have recently been used to noninvasively evaluate the mechanical properties of soft tissue, 51 –53 but to our knowledge, studies employing these imaging methods have not examined the effect of aging. Wide variability in the derived mechanical properties of skin can be found in the literature (Table 1) due to the different experimental conditions and assumptions regarding linearity, time dependency, and anisotropy when modeling the mechanical response of cutaneous tissue. 54
Young's modulus of skin and how it varies with loading condition, sample site, and age
All species are human unless noted otherwise
V, ventral; D, dorsal; NA, not available; SC, stratum corneum.
Mechanical testing typically involves evaluating the relationship between the force applied to tissue and its displacement. Typically, the applied force is normalized by tissue cross-section and reported as stress, while the displacement is normalized by tissue length and reported as strain. The slope of the stress–strain curve produced during mechanical loading provides a measure of Young's Modulus, which is a material property measuring tissue stiffness independent of the geometry (e.g., thickness) of the skin. Most mechanical studies of aging skin focus on the Young's Modulus, which is generally accepted to increase with advanced age (Table 1). However, due to the inherently different assumptions and loading conditions associated with tensile, indentation, suction, and torsional material testing, Young's Modulus can vary by orders of magnitude (Table 1) and will be discussed separately for each testing mode. It also is important to note that age-related changes to the structural (force-displacement) response can differ from the stress–strain response due to changes in skin geometry with advanced age. Previous work has also demonstrated that the skin's tensile strength 55 and the mechanical anisotropy also change with age. 56
Tensile testing
A nonlinear relationship between stress and strain (or force and displacement) is observed when skin and other collagenous soft tissues are mechanically loaded. This nonlinear response is largely related to the initial collagen organization and the fiber kinematics in response to tensile loading (Fig. 6). When a tensile load is initially applied to skin, the underlying microstructure in the dermis begins to realign and straighten in the direction of loading (Fig. 6a, b), resulting in significant tissue deformation. If the tensile load in that direction continues to increase, the dermal collagen fibers begin to straighten (Fig. 6c) and bear load, producing an increase in tissue stiffness. The individual fibers stretch until they reach a critical point and break. When a critical number of collagen fibers break with increasing load, the ultimate tensile strength of the tissue is reached and the tissue begins to visibly tear (Fig. 6d). Young's Modulus is typically measured as the slope of the linear portion of the stress–strain curve before failure during uniaxial tensile loading (Fig. 6). Notably, for other nondestructive mechanical testing modalities (e.g., suction, torsion, indentation), the collagen network may not be sufficiently engaged to yield Young's Moduli from a linear stress–strain response.
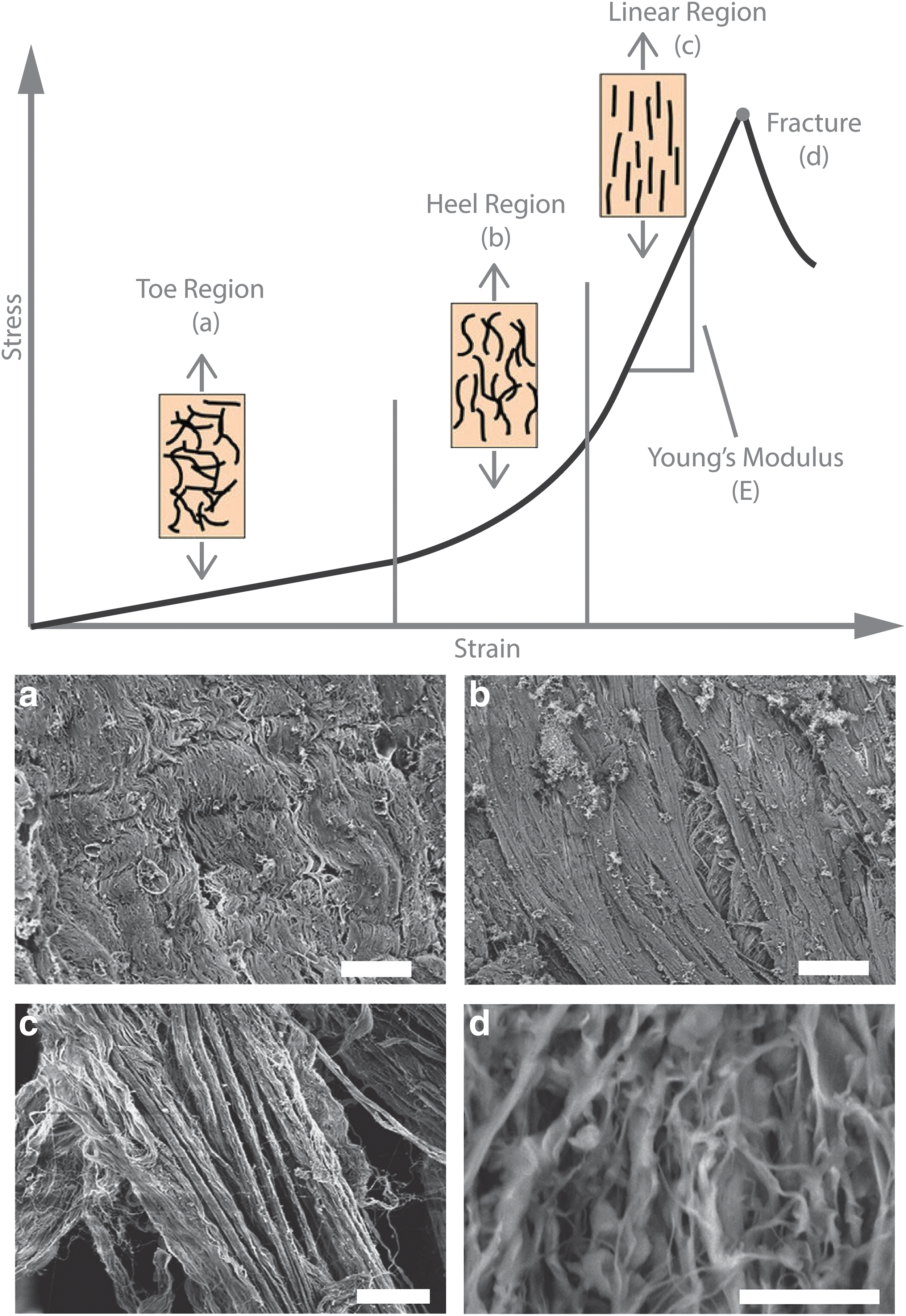
Schematic of how collagen fiber organization relates to the stress–strain curve during tensile loading. The toe region
Due to its simplicity and the straightforward derivation of stiffness, some of the earliest mechanical tests on cutaneous tissue were performed by applying a uniaxial tensile load to excised samples. 55 Jansen and Rottier used human cadaveric abdominal skin samples under uniaxial load and recorded E values of 1.27–1.96 MPa, and demonstrated an increased Young's modulus in samples over the age of 40. 55
While tensile testing generally results in greater E values than other testing modalities, the modulus of individual dermal collagen fibers is orders of magnitude greater than the bulk skin properties due to skin volume fraction occupied by components that do not bear load or are less stiff (e.g., elastin, HA). 54 Therefore, the volume fraction of collagen fibers within tissue can have a significant effect on the bulk material properties of skin. The reported increases in E with age are not consistent with the reports of decreased collagen density in the previous section. However, skin hydration is known to decrease with age, and hydration can have substantial effects on the measured skin geometry and mechanical properties.
Biaxial testing and constitutive modeling is often employed to obtain a more nuanced understanding of the material properties of soft collagenous tissues and their relationship to fiber alignment. A number of studies have demonstrated the mechanical response in one direction of loading is often dependent on the applied load in the orthogonal direction. 58,59 However, biaxial testing to evaluate the effect of aging on skin has been relatively rare. A study including uniaxial and biaxial experiments performed by Daly and Odland revealed a progressive loss in elastic and viscoelastic recovery in samples of 72–86 years of age compared with skin from 15 to 25 year olds. 60 In that study, it was hypothesized that a loss of elastic recovery may relate to a degenerative change in the elastin network.
More recent studies measuring collagen alignment and organization in the dermis of murine skin through collagen SHG imaging during mechanical loading report a lengthening of the heel region (Fig. 6b) with increased age. 61 This change in the mechanical response was attributed to age-related crosslinking within the dermis, and the degradation of proteoglycans that help retain water and thereby the viscoelasticity of the dermis. 61,62 The increase in the number of collagen crosslinks present in the dermis of older tissue may also increase the overall tissue stiffness. 33,62,63 In other tissues, such as the myocardium, nonenzymatic collagen crosslink density has correlated with the mechanical stiffness of the ECM. 33 In that study, nonenzymatic crosslink density was estimated through two-photon excited fluorescence (TPEF). Often used in combination with SHG imaging, TPEF can be used to provide high-resolution 3D maps of collagen crosslink autofluroescence and can highlight other key ECM proteins such as elastin. 33
Indentation testing
Indentation tests can target the mechanical properties of bulk skin tissue or the properties of single collagen fibrils in the dermal microstructure, depending on the experimental design and execution. In applications probing the bulk tissue response, indentation experiments require additional modeling to account for the contributions of underlying subcutaneous fat and muscle. 64,65 In vivo macroscale indentations of human forearm skin yield average Young's modulus values from 4.5 to 8 kPa, 64 which is orders of magnitude lower than tensile testing. Indentation tests of aged (57–79 years) skin reveal a slower elastic recovery than young (20–26 years) skin in the anterior forearm and thigh, which is attributed to the inability of aged skin to retain water after deformation and the increased presence of irreversible crosslinks. 66
A common method of performing microindentation techniques utilizes atomic force microscopy (AFM). This method is limited to ex vivo samples that have been sectioned and placed on slides, which must be considered when comparing this technique with other testing modalities that engage and reorient the collagen network. Human breast skin biopsies were tested using AFM and Young's modulus values of 0.1–10 kPa were measured, with an increasing modulus up to age 55 and a decreasing modulus in subsequent years. 67 Other AFM studies have focused on specific components of the skin such as type I collagen fibrils from the reticular dermis, and were able to find significant differences with age based on the hydration state of the samples. 18 The Young's modulus for dehydrated samples ranged from 4.19 to 8.11 GPa, with a significant decrease in aging samples.
Torsion testing
While tensile testing and some indentation approaches require excising skin samples, torsion tests provide an alternative to measure the bulk mechanical properties of skin in vivo with minimal discomfort. Torsion tests apply a constant torque to the skin and measure the resulting angular deformation to calculate mechanical properties such as the Young's modulus and viscoelastic properties. Agache et al. performed torsion tests on the dorsal forearm and reported the Young's modulus as 0.42 MPa in subjects under 30 years of age and 0.85 MPa in older subjects. 68 Similar studies report Young's modulus values of 1.12 MPa from human forearm skin that increase up to 20% after the age of 65. 69 A conflicting study reported that the Young's modulus of aged skin (0.02 MPa) was lower than that of young skin (0.1 MPa); however, the skin sample was not immobilized using a guard ring as in other studies. 68,69
While Young's modulus is derived from the linear relationship between stress and strain, other parameters have been defined to characterize time-dependent characteristics of the mechanical response. By recording skin deformation as a function of time, viscoelastic parameters can be derived from the tissue response following a change in load (Fig. 7). The amount of elastic deformation that can be measured as the initial, instantaneous deformation after a load is applied is called the immediate extensibility (Ue). With additional time, viscoelastic creep under constant load occurs producing an ultimate deformation of the skin (Uf). The immediate recovery of the skin (Ur) can be measured as the load is removed. Additional deformation with time can be measured to obtain the ultimate recovery of the skin (Ua).

Example of the deformation curve of skin produced under torsion or suction testing. Commonly reported parameters from this type of loading include: the immediate extensibility (Ue), the immediate recovery (Ur), the ultimate recovery (Ua), and the ultimate deformation (Uf). Color images are available online.
Ratios of these parameters have been used extensively to understand how the combination of viscous and elastic characteristics of the skin change with age (Table 2). A torsion test on the dorsal forearm of 138 subjects revealed decreasing Ue and Ur values after the age of 30, and later tests involving 145 subjects showed a decrease in Ue for skin greater than 35 years of age. 68,70 Decreases in these parameters are associated with hydrostatic recovery and may be related to a reduction in glycosaminoglycan (GAG) availability to bind water with age. 61,71 –73
Viscoelastic properties of human skin and how they vary with loading condition, sample site, and age
Suction testing
Similar to torsion tests, suction tests can also be performed in vivo, and have the ability to record multiple aspects of the skin's elastic and time-dependent mechanical properties. Rather than an applied torque, a vacuum is applied to the skin, and the Young's modulus (Table 1) or viscoelastic properties (Table 2) can be derived from the measured deformation and recovery of the tissue. Quantitatively, suction testing produces values of E that are close to that measured during standard tensile testing of excised skin. 74 In human trials, early suction studies on the dorsal forearm found a Young's modulus ranging from 1.8 to 5.7 MPa with significant increases in stiffness detected after the age of 70. 4 Measurements of skin from the back and forearms of patients from ages 2 to 67 years found a minimum in E between the ages of 15–25, and a gradual increase after 30 years of age. 75 Using an echorheometer, a device combining mechanical suction and ultrasound imaging, average Young's modulus values were reported from 0.08 to 0.26 MPa on the volar forearm, and the measured stiffness increased between 20 and 80-year-old subjects. Correlations between stiffness and age were also observed to be sex dependent, with men having a 70% greater increase in Young's modulus with respect to age than women. 76
Viscoelastic properties have also been measured using a Cutometer, a commercial device which measures the deformation of skin under a vacuum. The properties associated with the immediate and ultimate recovery of skin (Ur, Ua, respectively), as well as their proportion to the ultimate deformation (Ur/Uf, and Ua/Uf) following loading, are reported to decrease with age. 77 Efforts have also been made to isolate the properties of individual layers of the skin through suction tests and modeling. A combination study using suction, torsion, and computational modeling was used to derive Young's modulus for the SC (1.993 MPa) and the dermis (0.066 MPa). 78 The study reported that the modulus of the SC increases with age, while the elastic recovery ratio Ur/Uf of the SC decreased. 78
At the DEJ, a study by Kiistala separated the epidermis from the dermis by use of a suction device to produce small blisters on the skin of the ventral forearm and abdomen. 79 Blister formation, indicating a separation of layers at the DEJ, occurred significantly faster with increased age, which suggests an increased susceptibility to injury likely related to the retraction of the villi of the basement membrane and the shortening of the ridge and papillae microprojections in aged skin. 9,79 In addition to having an increased susceptibility to mechanical trauma, the changes to the microstructure and mechanical properties of aged skin summarized in this study may contribute to delays in the healing process and formation of nonhealing wounds. 80
Age-Related Differences in Wound Healing
After suffering a cutaneous injury, the wound healing response relies on the interaction of biochemical and mechanical signaling cascades to coordinate the stages of hemostasis, inflammation, proliferation, and remodeling that must occur for a wound to heal. However, it has widely been observed that aged skin wounds heal differently than young wounds, and are associated with delayed rates of closure and mechanically weaker tissue following repair. 7,81 These conditions place elderly patients at greater risk for developing chronic wounds, and consequently an increased chance of morbidity and mortality. 7 To improve wound care for the elderly, there is a need to better understand how the inherent differences in ECM organization and mechanics of aged skin affect the cell responses involved in wound healing.
Hemostasis and inflammation
With advanced aging, the initial hemostatic cascade is altered by increases in factors affecting platelet activation and coagulation. Fibrinogen increases by 10 mg/dL per decade of age. 82 Along with an increase in a variety of coagulation factors, women show a slight increase in antithrombin levels as they age, while men show a gradual decrease. 82 The platelet aggregation response to collagen is also greater in individuals over the age of 60, resulting in decreased bleeding times in the elderly. 82 Collagen crosslink accumulation and increased collagen network fragmentation combined with an increase in coagulation factors with age likely contribute to this altered hemostatic response. 83
The ensuing inflammatory phase of wound healing has been reported to be delayed in aged murine models, where assessments of leukocyte concentration after incisional wounding peaked at 3 days in young mice, but was delayed until 7 days in aged mice. 84 Causes commonly attributed to this age-related delay include altered GAG content, variations in mechanical stiffness and mechanotransduction, and a decline in macrophage function. As skin ages, total GAG content begins to decline and molecules like HA become more sparsely distributed throughout the basal epidermis and dermis (Fig. 8). 85
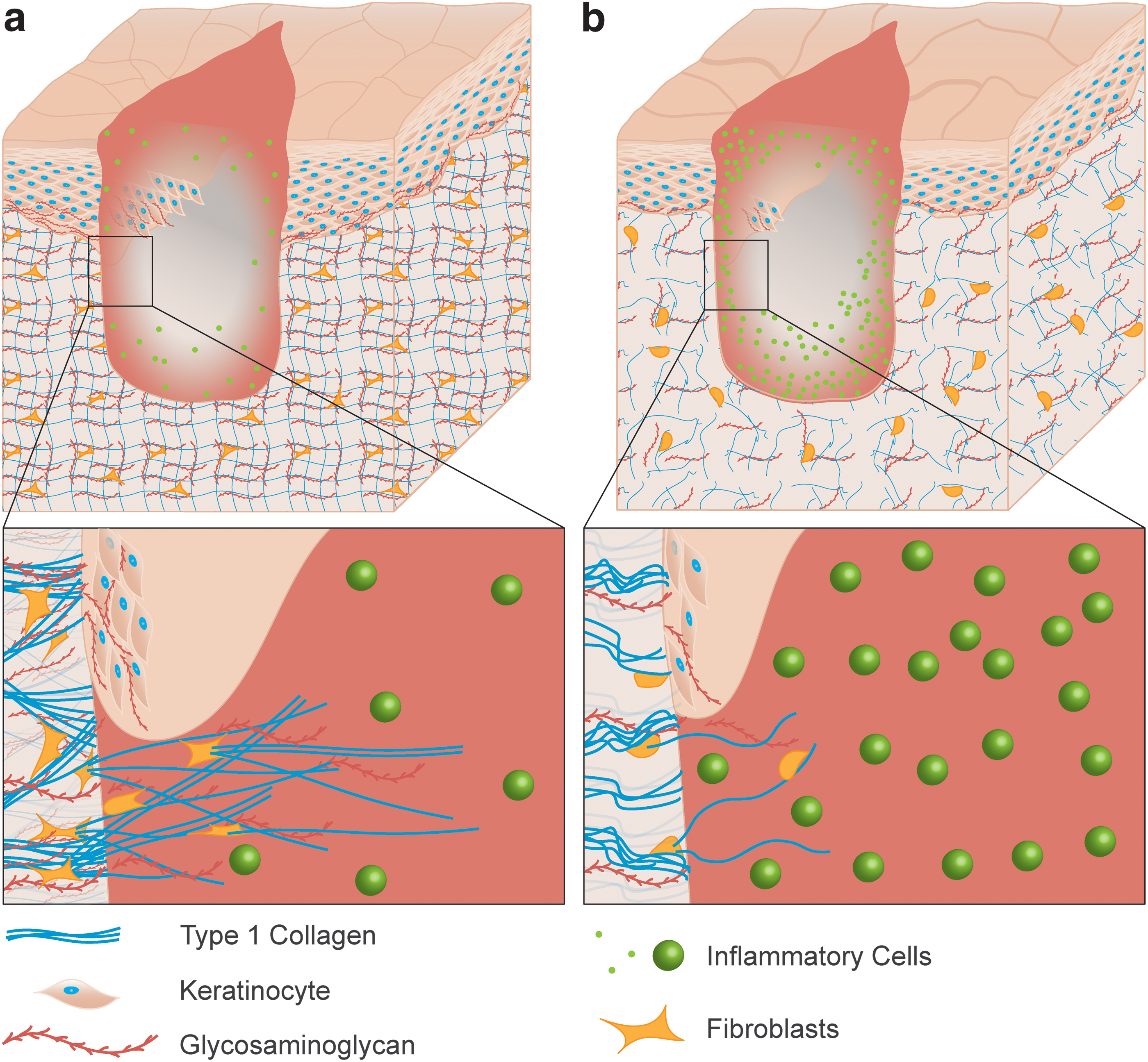
Schematic of a dermal wound in young
After sustaining an injury, HA functions as a promoter for inflammation, and a subsequent lack of HA in aged skin will delay the onset of this response. 86 Additionally, HA is responsible for water retention throughout the skin, and the decline of HA and similar GAGs with intrinsic aging likely contribute to the decline of viscoelastic recovery and increase in mechanical stiffness. 60 The increase in skin stiffness with age would be expected to shield a wound site from mechanical stress, resulting in reduced mechanical stimulation or mechanosensitivity within the wound. This stress shielding has been shown to reduce focal adhesion kinase (FAK) signaling and inflammatory cytokine pathways, 87,88 and may play a role in an extended inflammatory phase with increasing age. In addition, the mechanical changes to the skin may inhibit macrophage mechanosensation as substrate stiffness and deformation have been shown to modulate the macrophage inflammatory response. 89 Macrophages from aged individuals are noted to be less phagocytic 90 and produce fewer growth factors overall 91 than those from younger individuals. However, additional work is needed to understand the extent to which an altered mechanical environment affect the cells and structural changes associated with inflammation.
Proliferation and migration
Aged epidermal keratinocytes and dermal fibroblasts, both display reduced proliferative and migratory potential during wound healing. 7,92 –94 While this supports observations of slower reepithelialization and wound contraction in the elderly, the underlying causes for these delays are the result of a host of different biological factors that influence the structure and mechanical function of aged skin. Cellular senescence occurs naturally to maintain healthy skin, but the amount of senescent cells in the epidermis and dermis increases significantly with advanced age. 95,96 When the elderly suffer a cutaneous wound, the effects of increased senescence can directly impair skin regeneration and hinder migratory potential through inadequate maintenance of ECM structure. 43,97 While there is evidence that a small population of senescent cells improves granulation tissue formation and encourages wound closure through myofibroblast differentiation, 98 elevated levels of senescent cells are characteristic of chronic wounds 99,100 that experience prolonged inflammation and continued release of MMPs. 101 The onset of proliferation is delayed by this sustained inflammation, and in vitro assays of dermal fibroblasts suggest that the structural changes associated with increased MMP activity reduce the mobility and migratory potential of cells at the wound edge. 94 Furthermore, adipose-derived stem cells (ASCs) located in the dermis and hypodermis show the ability to differentiate into other cell types and contribute growth factors and cytokines to stimulate proliferation and migration during wound healing. 102 –104 Given the reduction of sWAT volumes and increase in cell senescence with age, changes in adipocyte and ASC function may also play important roles in age-related delays in healing.
The composition of the ECM, and GAGs in particular, play a significant role in the proliferative phase of healing. HA specifically participates in signal transduction to modulate proliferation and migration in the epidermis and dermis. 105 In healthy skin, HA can be found alongside proliferative keratinocytes in the basal epidermis where it maintains a hydrated matrix able to facilitate migration and reepithelialization in the event of injury (Fig. 8). 106,107 This is supported by studies where the reepithelialization rate was increased in samples of adult skin wounds treated with amniotic fluid containing high concentrations of HA. 108 As aged skin tissue has lower quantities of GAGs like HA compared with young skin, the resulting decreased hydrophilicity of the ECM likely inhibits the transport of signaling molecules and nutrients, leading to reduced keratinocyte proliferation and migration. 109
At the DEJ, the thinning and flattening of the rete ridges with advanced age combined with a decrease in GAGs and water retention contribute to the delayed viscoelastic recovery of the skin and an increased susceptibility to shearing injuries, such as blisters, in the elderly. 110 In addition to the epidermis, HA is found in the early granulation tissue in the pericellular regions around migrating dermal fibroblasts. When these fibroblasts were isolated from aged patients, the amount of HA in the pericellular region was lower than in young samples corresponding to a decrease in receptor interactions that lead to myofibroblast differentiation. 111
Myofibroblasts in the granulation tissue synthesize ECM molecules like collagen I and III as well as generate a contractile force that helps to close skin wounds. 112 While healthy wounds rely on tension from myofibroblasts initially formed in the granulation tissue to stimulate fibroblasts' differentiation into myofibroblasts, granulation tissue from older patients suffers from decreased ECM deposition and increased MMP activity that likely leads to delayed or inadequate tension generation. This is supported by animal studies in which the granulation tissue of aged mice was less mature than in young wounds, and was characterized by increased levels of inflammatory cells. 84
The tension created in the wound is also responsible for encouraging ECM alignment during the formation of adhesion sites for fibroblasts and myofibroblasts, 113,114 which in turn promotes directional migration into the wound. 62 As the collagen network from aged individuals is correlated with altered fiber organization and alignment in the wound, 62 closure is further inhibited as directional migration becomes more difficult in aged tissue. Furthermore, nonenzymatic collagen crosslinks that accumulate with advanced age have been directly implicated in hindering fibroblast contraction and tension generation during healing. 115
Remodeling
Aged wounds have long been noted to gain tensile strength more slowly than young wounds, 116 –118 and ultimately develop scar tissue of weaker mechanical strength in both animal models 119,120 and humans. 116,121 Due to the stiffness of aged skin, stress shielding within the wound site may result in reduced collagen deposition by fibroblasts during remodeling (Fig. 8). FAK activation of dermal fibroblasts is potentiated by mechanical loading in the ECM through extracellular related kinase (ERK) during healing and plays a significant role in fibrosis. 122 In stiffer aged skin, the decrease in stress-sensitive FAK signaling may contribute to decreased collagen deposition by reduced secretion of monocyte chemoattractant protein-1 (MCP-1) and a resulting downregulation of the inflammatory chemokine pathways that regulate fibrosis. 87
These changes in ECM organization are supported by observations made by Ashcroft et al. who reported that macroscopic and microscopic examination of scars in aged human and animal models were less fiber dense than scars in young tissue. 7 Additional studies in murine models found similar results from incisional wounds after closure because collagen fiber bundle density within the wound was lower with advanced age. 84 The young mice showed a greater concentration of relatively aligned type I collagen bundles dispersed throughout the wound up to 56 days postinjury, whereas the old mice were consistently observed to have smaller, less dense networks of fibers that seemed to lack a favored alignment direction similar to unwounded tissue. 84
Variations in collagen alignment likely effect mechanotransduction cues because tensile loads are distributed differently between aligned and unaligned networks. 1 Reduced collagen deposition and altered organization in the healed wounds of aged individuals may also be the result of high levels inflammation and dysfunction in ECM turnover resulting from intrinsic aging or age-related diseases. 90 High levels of inflammatory cytokines and altered MMP and TIMP levels after healing may therefore predispose aged skin to failure and the recurrence of chronic wounds. 123 Collectively, these observations indicate a clear reduction in collagen deposition and organization within aged wounds, as well as a decrease in the mechanical strength of wound scars with advanced age.
Conclusion
Summary
Intrinsic aging causes many underlying changes to the behavior, sensitivity, and signaling pathways of cells in the skin (Fig. 9). As we age, dermal fibroblasts begin to show signs of reduced sensitivity to mechanical stimulation, resulting in a subsequent decrease in collagen synthesis and increase in active MMPs. Changes in matrix remodeling result in a reduction in total collagen, and a coarser dermal collagen network 15,21 with more crosslinks and lower GAG content than younger tissue. 1,32,63 These changes to ECM structure result in altered mechanical function, in which aged skin exhibits a higher Young's modulus, decreased viscoelastic recovery, and lower tensile strength than young skin making it susceptible to traumatic and pressure-related injury. 80
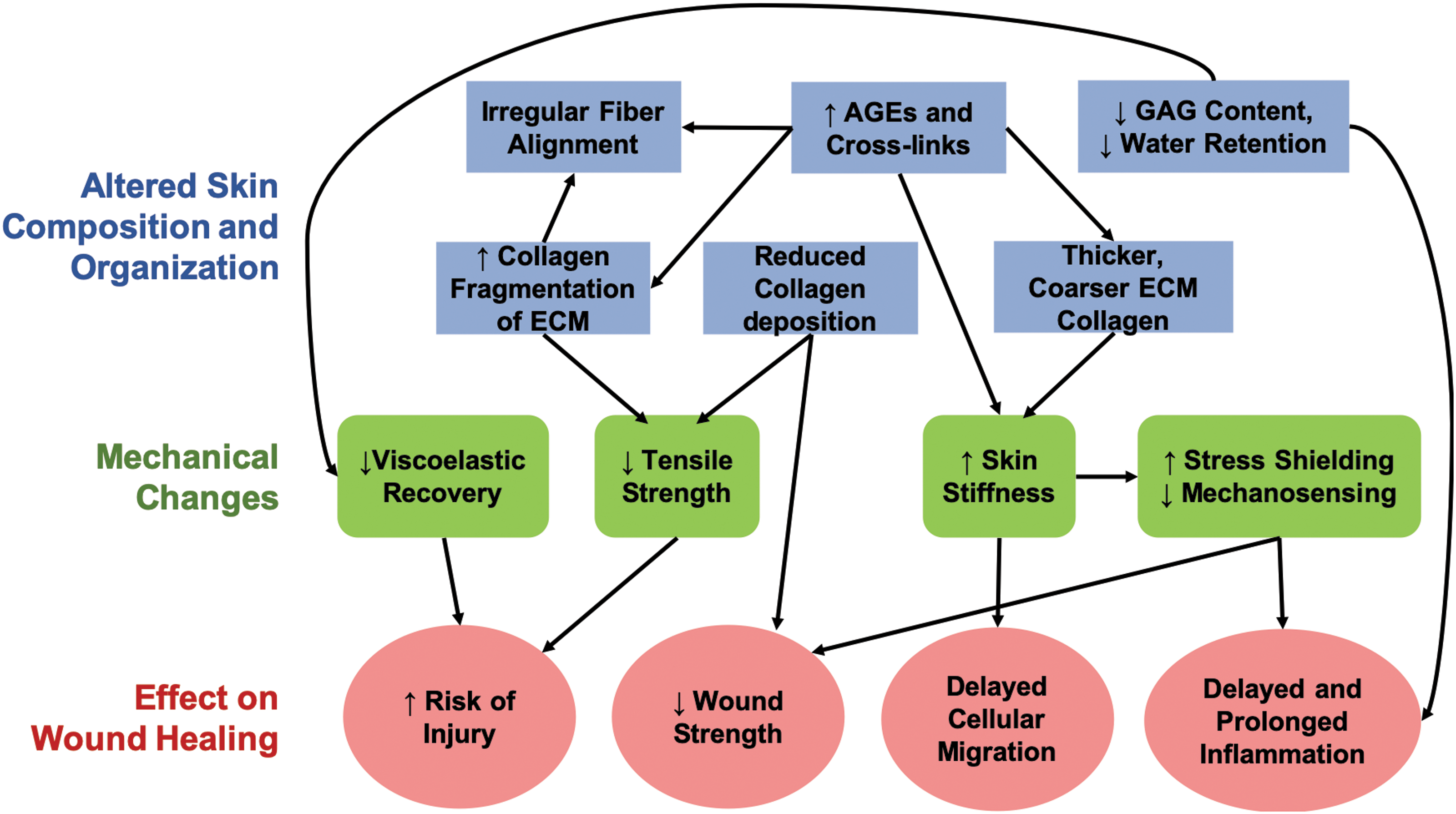
Graphical representation of the biological changes in aged skin that cause downstream effects on ECM structure leading to altered mechanical properties. Color images are available online.
Once a wound develops, the ability of aged skin to regenerate and repair itself is significantly decreased due the biological and biomechanical effects of aging. Stiffer aged skin can stress shield the wound, which may lead to a delayed and prolonged inflammatory response as well as inhibiting fibroblast differentiation and migration. Irregularities in collagen alignment and deposition, along with altered GAG content, can further hinder the migration of cells into the wound site. Increased cellular senescence and upregulated MMP activity can protract inflammation, inhibit proliferation, and lead to a decreased fibrotic response making aged wounds mechanically weaker after closure. Any combination of these factors can result in delays to different phases of healing, slower rates of wound closure, and increasing risk of wound chronicity.
Future directions and outlook
Additional work is needed to elucidate the complex relationships between tissue structure, mechanical function, and cell mechanobiology that change with advanced age in the skin. Future research with human volunteers or patients are needed, but the range of comorbidities with advanced age make it difficult to study the effects of solely intrinsic aging on the wound healing process. It is also important that this future work accounts for other biological factors, such as sex, when characterizing age-related changes in the skin. Additional work is also needed to better characterize the elastic response of skin and evaluate how the nonlinear stress–strain response and mechanical anisotropy change with age.
Furthermore, studies of the relationship between skin microstructure and mechanical function are generally limited to nondestructive mechanical tests. Despite these challenges, emerging research into elastography utilizing noninvasive clinical imaging modalities, such as ultrasound, 124 optical coherence tomography, 125 –127 and magnetic resonance 65,128 with mechanical testing, is expected to aid in understanding heterogeneity in the mechanical properties of the different layers of the skin and wounds.
Preclinical wound healing research using aged animal models will also be necessary to understand the failure properties of both skin and wounds, as well as their underlying relationship to the tissue composition and organization. Advanced noninvasive imaging techniques such as SHG allow for 3D evaluation of the tissue microstructure under load.
Recent studies have used SHG to evaluate metrics such as collagen density and coarseness, 26 as well as intensity between aged and young patients. Entire dermal collagen networks have even been imaged in vivo during uniaxial stretching to quantify changes in fiber alignment between aged and young tissue to provide direct comparison of microstructural changes during applied load. 61 SHG also has the added benefit of being acquired simultaneously with techniques like TPEF and FLIM that can provide information about underlying changes in cellular metabolism during healing 129,130 and collagen crosslinking status. 131 Combining quantitative measures of tissue microstructure with advanced modeling techniques 132 may yield additional insights into the relationships between structural changes and altered mechanical function at the micro- and macroscale.
With our emerging understanding of the mechanical properties of the skin through advanced imaging techniques, we also need to elucidate and verify the relationships between the mechanical microenvironment and cellular responses during healing. Through studies investigating the role of the FAK–ERK pathway, 122 fiber alignment before and during healing, 61 and cellular responses to mechanical stress, 62 there is an increased appreciation for the role mechanobiology plays in aging and wound healing. The effect of age-specific mechanical changes, such as increased stiffness and decreased viscoelastic recovery, on wound healing outcomes requires further exploration. Moreover, despite a higher prevalence of wound reopening in the elderly, 116 there is a dearth of information pertaining to skin and wound strength due to the destructive nature of this type of mechanical testing. Given the changes in collagen fiber density, organization, and crosslinking with advanced age, noninvasive microscopy techniques may be useful in predicting wound strength and the risk of reinjury.
Skin composition and organization changes with age.
Changes in the skin microstructure can affect its stiffness and alter the response of different cells involved in wound repair.
A prolonged inflammatory response and delayed reepithelialization in the wounds of aged skin may be related to an altered mechanical environment.
Aged skin is more prone to injury, and the strength of scars formed by aged skin is reduced compared with younger skin.
Footnotes
Acknowledgments and Funding Sources
This work was funded by NIH grant number R01AG056560 as well as the Arkansas Biosciences Institute.
Author Disclosure and Ghostwriting
No competing financial interests exist. The content of this article was expressly written by the authors listed. No ghostwriters were used to write this article.
About the Authors
Funding Information
This work was funded by NIH grant number R01AG056560 as well as the Arkansas Biosciences Institute.