
Abstract
Significance:
Since the last Food and Drug Administration (FDA) approval of a wound healing therapeutic in 1997, no new therapeutic candidate (excluding physical therapies, devices, dressings, and antimicrobial agents) has advanced to clinical applications. During this period, the FDA drug approvals for tumors, which have been referred to as “wounds that do not heal,” have reached a total of 284 (by end of 2018). Both political and scientific factors may explain this large discrepancy in drug approvals for the two seemingly related and equally complex pathophysiological conditions.
Recent Advances:
Using the current research funding ratio of 1:150 for wound healing to cancer and the 5% FDA drug approval rate for oncology, we reach a crude estimate of a 0.03% success rate for wound healing therapeutics. Unless a drastic improvement of the current situation, we express a pessimistic outlook toward new and effective wound healing drugs.
Critical Issues:
We argue that successful development of wound healing therapeutics will rely on identification of wound healing driver genes (WDGs), and the focus should be on WDGs for the wound closure phase of wound healing. Therefore, WDGs must be both necessary and sufficient for wound closure; the absence of a WDG disrupts wound closure, while its supplementation alone is sufficient to restore full wound closure. Successful translation of a WDG into therapeutics requires availability of well-defined animal models with a high degree of relevance to humans. This review discusses the main hurdles faced by the wound healing research community behind the development of so-called “rescuing drugs” for wound healing.
Future Directions:
Given the lack of new wound healing drugs for the past 23 years, there is a need for a wide range of fresh, innovative, and thorough debates on wound healing drug development, including an organized movement to raise public support for wound healing research.


Scope and Significance
The general public and patients may assume that the wound healing research community receives an equitable share of government funding for their research endeavors. To understand why advances in wound healing drug development have lagged behind advances in most other human chronic diseases, such as cancer, we present both political and scientific arguments, for discussion and debate. Using cancer research as a relevant comparison, this article highlights the funding inequities and previously overlooked obstacles for development of new wound healing therapeutics.
Translational Relevance
We call for focus of the already limited R&D resources on identification of wound healing driver genes (WDGs) for the wound closure phase of wound healing. We present our view on the reasons behind the failure of growth factors as wound healing drugs over the past few decades. To achieve the goal, we need animal wound healing models relevant for corresponding (acute or chronic) human wounds.
Clinical Relevance
Wound healing is a common part of the recovery phase in many human disorders. When this part fails to complete on time, acute pathological conditions become chronic, causing long-term health problems for patients. The three major types of chronic skin wounds in humans, venous ulcers (VU, >70%), pressure ulcers (PU, 15%), and diabetic (neuropathic) foot ulcers (DFUs, 8%), pose a major medical, social, and economic problem in the United States and a far worse conundrum in developing countries.
Background and Overview
150:1 research funding ratio for cancer versus wound healing
Cancer is a family of diseases with an incidence rate of 1.7 million and mortality rate of 0.6 million per annum in 2018. The National Cancer Institute (NCI) was founded in 1937. Guided by the National Cancer Act in 1971, which greatly broadened its scope and responsibilities, the NCI has expanded from its initial 15 cancer centers/laboratories to more than 70 research facilities across the United States. The NCI receives 12–18% of the total annual NIH budget in funding as well as funding from numerous other government and non-government agencies: Centers for Disease Control, the National Institute for Environmental Health Sciences, and the American Cancer Society, just to mention a few. Grant applications for cancer research have also been on the rise, with a 46% annual increase compared to less than 10% for the remaining 18 human disease categories. 1 Despite the NCI's massive funding support over the decades, it has faced challenges in delivering on its mission and meeting its own timeline to virtually eradicate cancer, as to date, no drugs have been approved for prevention of tumor metastasis, the main cause of death in cancer patients. The majority of the existing 284 oncology drugs target cell proliferation of primary tumors, extending patients' survival, but often losing efficacy after several months of treatment due to new mutations in the drug target genes in patients. 2 The widely recognized five common steps of cancer progression, including gene mutations, hyperplasia, dysplasia, primary tumor, and metastasis, are highly complex and, therefore, may justify the magnitude of political and financial support that cancer research has received from the public. However, many of these disease mechanisms are by no means wholly unique to cancer. In fact, the mechanisms involved in wound healing have considerable overlap with tumor development, and these two pathologies have widely been considered equal in terms of their complexities. More than 30 years ago, such a connection between tumor environment and wound healing environment was pointed out by Professor Harold Dvorak, who famously described tumors as “wounds that do not heal.” 3 Like tumors, wounds occur in many different organs in both acute and chronic forms. Mechanistically, wound healing shares a similar degree of complexity with cancer progression, including an intricate series of steps consisting of hemostasis, inflammation, proliferation, and migration, and microenvironmental remodeling, as schematically illustrated and compared in Fig. 1. The best-known example underlying the concept of tumors as wounds that do not heal is perhaps the progression of nonalcoholic liver cancer: fatty liver > liver fibrosis > liver cancer. 4,5 However, mechanisms of tumor progression vary greatly among different types of tumors. Our point here is not to downplay the milestones made in cancer research. Instead, we want to raise awareness of the potential higher degree of difficulty for developing wound healing therapeutics (specifically defined here as “gene target-based” therapeutics).
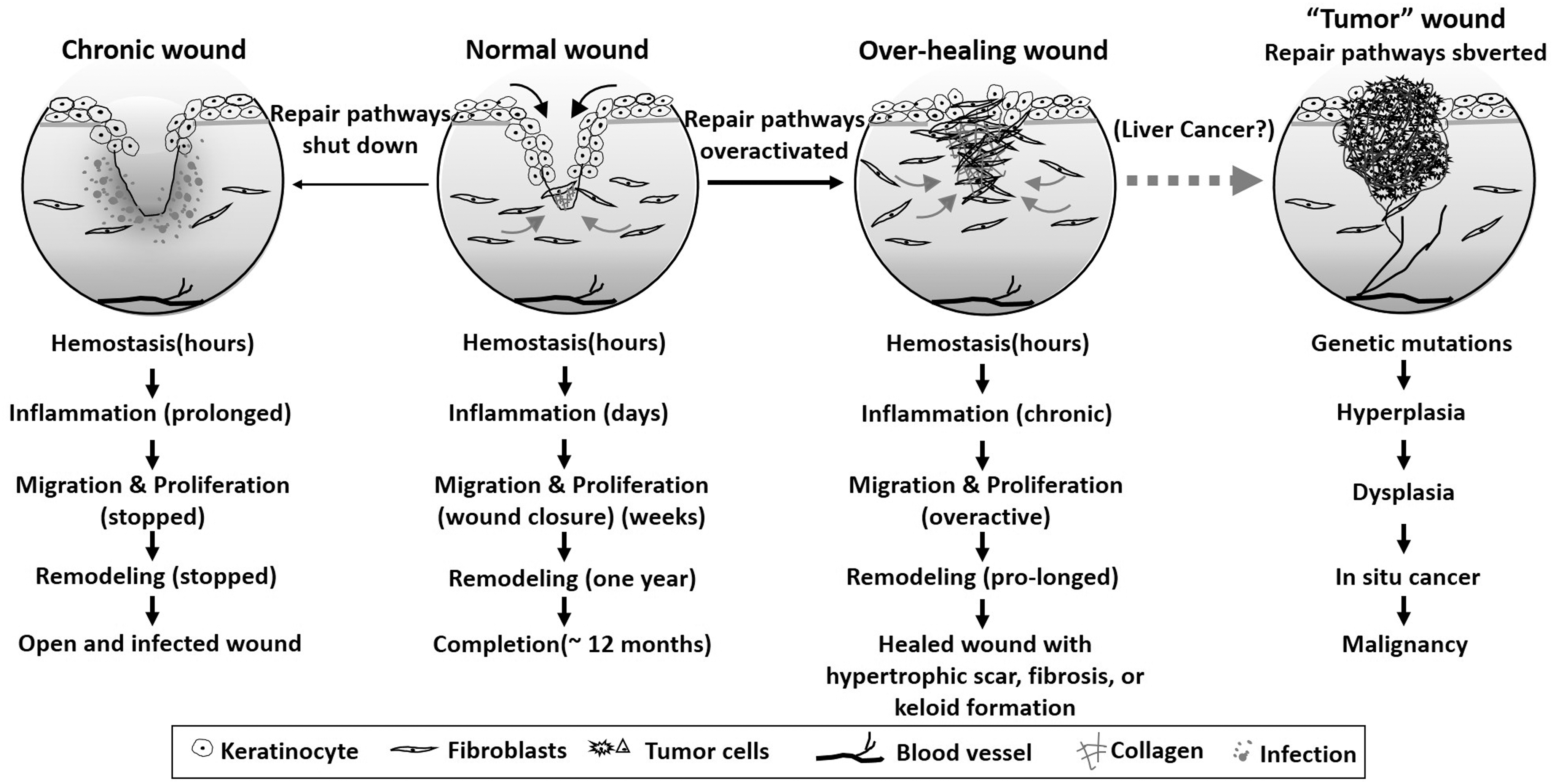
Pathology of wound healing and tumor progression. When the normal wound healing process becomes derailed by various systemic pathological conditions, the healing process may either be inhibited (left) or overactivated (right) leading to scarring and fibrosis. Tumors organize and utilize a similar “wound” microenvironment to support growth, invasion, and metastasis. Both processes share many common mechanisms and similar degrees of complexity.
Chronic wounds affect millions of Americans and cost the health care system $28.1–96.8 billion annually, without counting the costs associated with traumatic, burn, chemical, infectious, radiation, and postsurgical wounds, and disproportionately affect sick veterans, the elderly, and the poor. 6 Clinical data on some chronic skin wounds, such as neuropathic diabetic ulcers, reveal an even higher reported 5-year mortality rate compared with many common cancers. 7 –9 Nonetheless, wound healing is not listed as a specific area of research by any of the 11 agencies in the NIH. Several studies led by Professor Robert Kirsner reported rather chilling data on wound healing research funding in the United States. Their findings show that cutaneous wound healing accounts for only 0.1% of the NIH budget and 0.15% of the total medical research funded by the non-NIH federal sector. 10,11 Therefore, the NIH with or without other government agencies has been investing 150 times more in cancer research than in wound healing research for the past four decades or longer. Considering the consensus in the scientific community that cancer progression and wound healing share similar degrees of complexity, as well as patient morbidity and financial burden to health care systems, it is hard to comprehend how these two entities have been viewed with fundamentally opposite attitudes. Accordingly, the repeated calls for increased research funding by wound healing investigators often fall on deaf ears. Their grant applications have been viewed as non-mainstream subjects, which must often be incorporated into applications in various NIH study sections that focus on diseases other than abnormal wound healing. Public recognition of wound healing as a major medical challenge is virtually absent. For instance, a large portion of the 350 million people in the United States knows about “diabetes” and perhaps its kidney complications, but few have knowledge about the health, social, and economic burdens of DFU, including it being (i) the #1 cause of diabetes-associated hospitalizations, (ii) the main cause of 45–70% of all lower limb amputations, especially among minorities and the poor, (iii) a major reason for leg loss every 30 s in the world, 12 (iv) the etiology behind 50% of 2 year postamputation deaths, 13 and (v) the main factor contributing to a spike in amputation rates, 14 a problem compounded by the soaring costs of insulin—a top priority of the American Diabetes Association. 15 Like cancer, DFU must be caught early and treated to prevent continued progression and patient suffering. With timely screening, in-depth assessment of symptoms and thorough physical examinations, millions of amputations could have been prevented. 16 Moreover, DFUs only represent less than 8% of all chronic wounds in the United States. The hope is that finding a cure for DFU may serve as a starting point for other major types of chronic wounds, such as VU and PU.
Thomas et al. from BIO (the world's largest trade association representing both academic institutions and the biotechnology industry), BioMedTracker (a company that tracks the clinical development and regulatory history of investigational drugs to assess its likelihood of approval [LOA] by the Food and Drug Administration [FDA]), and Amplion (the leading biomarker business intelligence company) recently conducted a study to determine the FDA's LOA for various drugs. Their comprehensive approach gathered data from 9,985 clinical and regulatory phase transitions from 2006 to 2015—representing 7,455 company-sponsored and FDA-registered development programs across 1,103 companies. Their analysis shows that the overall FDA's LOA starting from Phase I for all developmental candidates was 9.6%, with the highest LOA of 26.1% for hematology and the lowest LOA of 5.1% for oncology, as illustrated in Fig. 2. 17 Although unknown, the LOA for wound healing therapeutics is probably similar to that of oncology, considering the similar degrees of complexity in pathophysiology. While many other factors besides LOA and funding could influence drug development, it would be reasonable to assume a dire future for new wound healing therapeutics if the current status quo is upheld. As an old Chinese idiom puts it, “巧妇难为无米之炊,” meaning, “Even the cleverest housewife cannot cook without rice.” At the end of the day, it would not be possible to convince industries to invest in a drug development program that has less than a 0.03% chance for FDA approval.
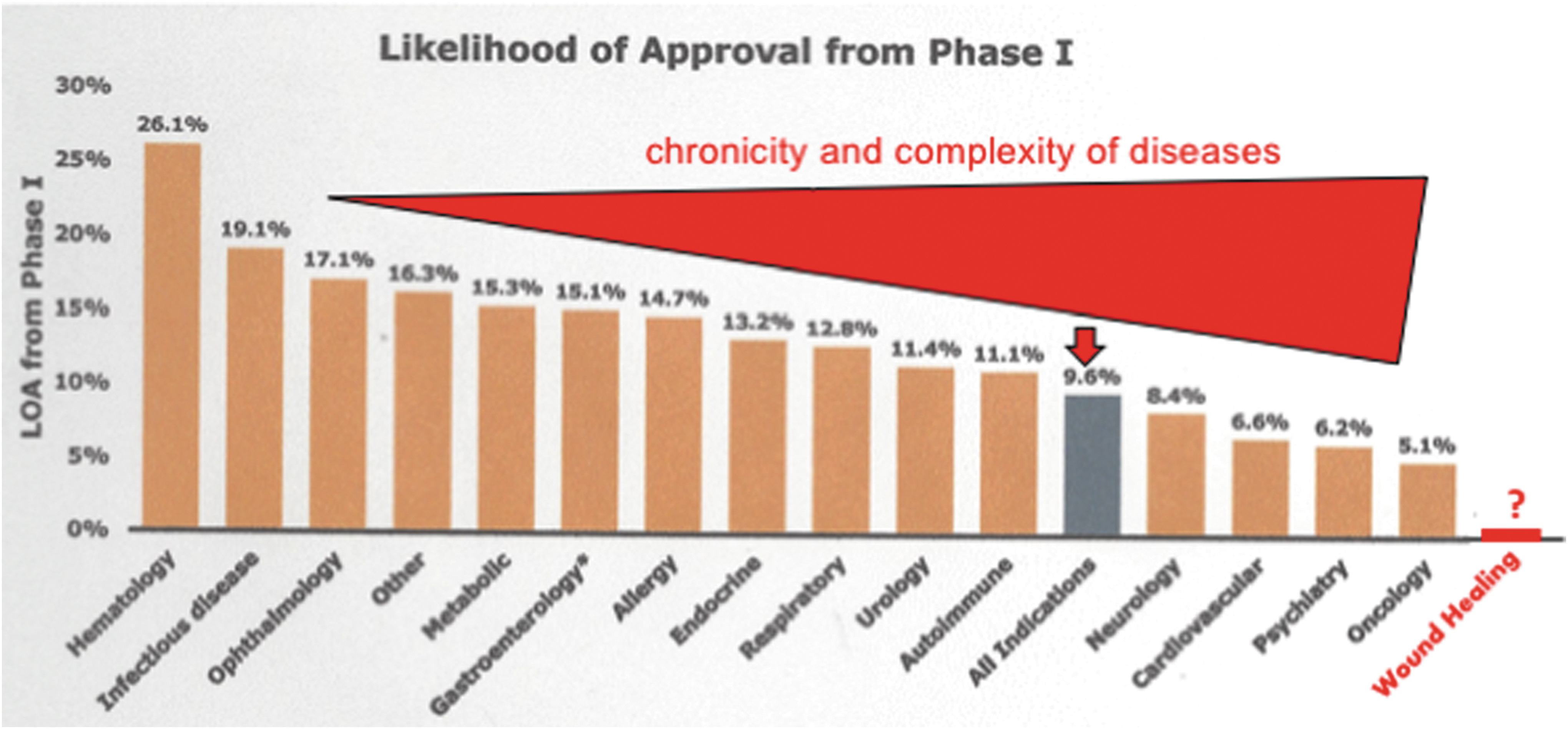
Chart of LOA from highest to lowest by disease area. This figure is from the original fig. 2 of the report “Clinical Development Success Rates 2006–2015” by Thomas et al. 17 and adapted with the authors' permission. The “wound healing” as a disease area and the complexity bar were added by authors of this article. From an LOA of 26.1% in hematology to 5.1% in oncology, there seems to be a negative correlation between success rate and disease complexity. Since the complexity of wound healing is equivalent to that of tumor progression, but the ratio of NIH funding for cancer research versus wound healing research is 150:1, the projected success rate of wound healing drugs is close to zero. LOA, likelihood of approval.
Cancer drugs are a “blocker,” but WDGs must act as a “rescuer”
Many genes have been proven essential for a given pathophysiological process, but few turned out to be sufficient by themselves to drive the biological process to completion. In cancer, a so-called cancer driver gene (CDG) is defined as a gene which when mutated increases net cell growth under the specific in vivo cellular microenvironment. 18 A typical solid tumor contains hundreds or thousands of genetic alterations, point mutations, small insertions, or deletions, but only a few of the mutations are “drivers,” conferring a selective growth advantage to the cancer cells. The remaining thousands of mutations are “passengers” that coincidentally arise during the large number of cell divisions associated with the neoplastic process. 19 –22 However, according to the initial “Two-Hit” 23 and later improved “Multi-Hit” 24 theory of cancer causation, no single oncogene is able to cause cell transformation and tumorigenesis. Instead, cancers develop because of randomly induced series of multiple mutations in otherwise normal genes, together with mutations in tumor-suppressor genes, which normally function to apply the brakes to cellular growth. In other words, these oncogenes or tumor suppressor genes are necessary, but not sufficient, by their own individual capacity to cause cancer. A typical example is the ras oncogene (v-ras), which has been detected in more than 30% of human cancers. 25 However, overexpression of v-ras failed to transform primary cells, instead causing cell senescence. 26,27 Activation of a single oncogene or inactivation of a single tumor suppressor gene, by itself, is not sufficient to drive cancer progression. 28 Mechanistically, many of these CDGs are key components in a signaling network. Therefore, loss of function of any one key component could change the functionality of the entire network. Taking a scaffold structure as a comparison, missing any individual tube or coupler will cause catastrophic collapse of the entire scaffold. CDGs act like these individual tubes or couplers in signaling scaffolds and knocking off any one of them could nullify the entire signaling network. Accordingly, the sole qualification for a large portion of anticancer drugs is to act as a “blocker” that interrupts a key signaling component of a constitutively activated signaling network in cancer cells, even if all other components of the same signaling network remain functional. In contrast, a gene product with therapeutic potential for promoting wound healing must not only be essential for the multiple-signaling component networks but also be sufficient by itself to activate the functionally halted networks, such as in chronic wounds, to jumpstart the repair process. Specifically, the requirements for an effective wound healing therapeutic candidate are (i) to play an essential role, similar to a CDG, and (ii) to act as a “rescuer” that “wakes up” a minimum number of the signaling components in the networks to reinitiate the disease-halted healing process. Therefore, unlike a CDG, which can be an intracellular kinase, a metabolic enzyme, a cell cycle regulator, or a transcriptional factor, an effective wound healing factor has to be a more upstream activator of all the above.
To raise awareness of the higher hurdles surrounding the development of wound healing therapeutics, in this study, we propose the concept of WDGs and the requirements for completing the wound closure phase. A WDG must be not only necessary, like a CDG during cancer progression, but also sufficient by itself to promote completion of the most critical phase of wound healing—wound closure. An unanswered question is whether any gene product or combination of gene products qualifies as a WDG or a “rescuer” during wound healing. Eming et al. have argued that no single tissue component, growth factor, extracellular matrix (ECM), or cell type is sufficient on its own to resume the healing process in a chronic wound. Instead, a combination of multitarget therapies is required. 29 This notion of multitarget comprehensive therapies may well reflect the reality, not only for chronic wound healing but also for other chronic diseases. Based on this argument, we would first need to obtain a comprehensive understanding of how different components act together spatially and temporally to restore injury. However, if this grandiose objective is treated as the be-all and end-all, causing investigators and drug companies to reject any single molecule drug candidate, it is not hard to predict that the paucity of new wound healing therapeutics will continue for several foreseeable generations. This is because the development of a single compound, including toxicity analysis, pharmacodynamics, pharmacokinetics, and efficacy in humans, would already take more than a decade, cost several billion dollars, and yet has a very low success rate. Attempting to develop a “comprehensive” and multicomponent regimen in one fell swoop would certainly be beyond the maximum operating capacity of any academic or industrial entity. Having already suffered from the lack of new and effective chronic wound therapeutics over the past several decades, during which single-molecule drugs have steadily received FDA approval for many other chronic disorders, chronic wound patients have a right to demand an answer. Therefore, we must keep the door open and the hope alive.
Focus on WDGs that promote wound closure for therapeutic development
Normal wound healing is a year-long process that progresses through several overlapping phases: blood clotting (hemostasis), inflammation (immune response), tissue growth (cell migration and proliferation), and dermal tissue remodeling (ECM replacement and neovascularization). 30 The first three phases are normally complete within a few weeks and are collectively known as the “wound closure phase,” widely recognized as the ultimate outcome for wound healing research, clinical trials, and practice. The U.S. FDA definition for complete wound closure is 100% wound surface reepithelialization without discernible exudate, drainage, and dressing requirements confirmed at two consecutive study/clinical visits 2 weeks apart. 31,32 The subsequent remodeling phase following wound closure lasts over a year. Presently, there are few animal models and methodologies available to evaluate deep tissue integrity. 33 The development of wound healing therapeutics will remain a long and uphill battle due to the difficulty of finding, designing, and developing a “rescuer” drug. We propose focusing on WDG candidates that directly promote wound closure. As listed in Fig. 3, we suggest that only those WDG candidates that meet all five criteria may expedite advances in wound drug development. Another argument to concentrate on wound closure rather than remodeling arises when we consider that wound healing in humans occurs largely through a repair process rather than regeneration. Regeneration is the natural process of replacing or restoring damaged or missing cells, tissues, or organs to full function in plants and animals. In humans, however, except for a few appendages, such as hair, nails, and the top epidermal layer, the vast majority of cutaneous wounds are healed by repair, which will not give rise to the original architecture before the injuries occured. 34 Skin regeneration is another frontier currently being explored by scientists. However, new WDG drug candidates that drive wound closure should be prioritized, given that regenerative therapies may take much longer to be developed and to become widely available for clinical use.
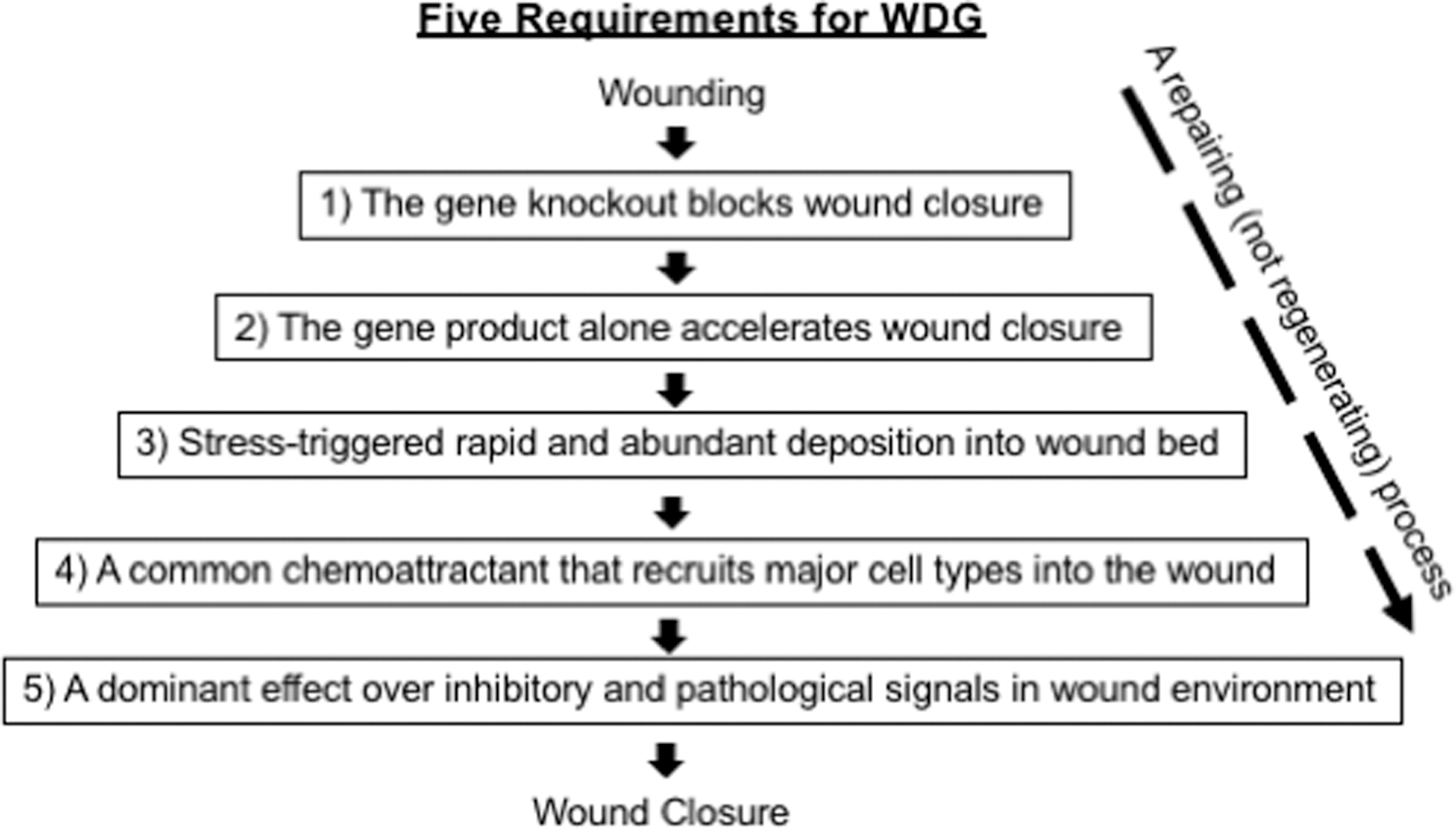
Requirements for a WDG candidate before drug development. (1) Genetic studies of the gene indicate necessity; (2) the correction of delayed wound closure by gene product supplementation alone in various animal models suggests sufficiency; (3) rapid accumulation of the gene product in the wound bed in response to wound stress signals, especially ischemia; (4) directional migration of the wound-surrounding cells, including immune cells, toward the wounded area must precede first for wound closure; and (5) the newly released WDG(s) must be functionally stable and minimally affected by the hazardous wound environment. WDG, wound healing driver gene.
WDGs cannot come from the circulation
In analyzing the environmental changes between unwounded and wounded skin, our studies revealed a critical regulatory factor. In intact skin, the epidermal and dermal cells are bathed in interstitial fluid, largely a filtrate of plasma from the circulation. Following acute cutaneous injury, the most dramatic microenvironmental change that occurs once intact blood vessels are damaged, is the influx of platelet-degranulated serum from vessel to wound bed. The cells at the cut edge of the wound experience the transition from an environment of plasma to an environment of serum in the wound milieu for the first time. As the wound heals, the cells are transitioned from serum environment back to plasma environment. We found that human serum promotes human keratinocyte migration significantly more than human plasma in vitro. 35 This finding suggests that the promotility factor for keratinocytes, which is responsible for wound surface reepithelialization, becomes available only after transition in the environment from plasma to serum. Triggered by this surprising observation, we went on to test the effect of human plasma versus human serum on migration and proliferation of all three major human skin cell types: epidermal keratinocytes, dermal fibroblasts, and microvascular endothelial cells. We reported that human plasma strongly promotes motility of the two dermal cell types. To our surprise, human serum halts migration of the two dermal cell types, while it selectively promotes migration of keratinocytes. Moreover, while human plasma promotes proliferation of all three cells types, human serum inhibits proliferation of all the skin cell types. 36 We further identified the “blocking signal” in human serum that selectively prevents dermal cell migration and inhibits proliferation of all three cell types as transforming growth factor-beta3 (TGFβ3) (not TGFβ1 or TGFβ2), which is detected in human serum, but not in human plasma. To understand the mechanisms behind the differential actions of TGFβ3, we demonstrated that the selective sensitivity of human dermal cells to TGFβ3 inhibition is due to their 10-fold higher expression levels of the TGFβ type II receptor (TβIIR) compared to human keratinocytes. Reduction of the TβIIR levels in dermal cells to the levels in keratinocytes alters dermal cell proliferation and motility responses to match that of keratinocytes in human plasma or serum. In reverse, increasing TβIIR expression in human keratinocytes, even in human melanocytes, to the levels in human dermal cells sensitizes keratinocytes to the antimotility effects of serum and TGFβ3. Since the antimotility and antigrowth effects of the TGFβ family cytokines override the promotility and promitogenic effects of growth factors, 37,38 these findings suggest that the driving force for proliferation of all three skin cells and migration of the dermal cell in the wound cannot come from the circulation. Growth factors, such as PDGF-BB, FGF, and VEGF, may be indeed present in the wound environment, but are unable to execute their functions due to the presence of TGFβ3. We interpret the biological significance of TGFβ3 in wound healing as that of a “traffic controller,” in which TGFβ3 orchestrates the orderly migration of epidermal (immediately) and dermal (several days later) cells throughout the naturally occurring plasma > serum > plasma transition during wound healing. 36 This interpretation was recognized as an “Editor's Choice” by Science (312:162, 2006).
Conventional growth factors cannot be WDGs
Since the discovery of the first growth factor more than 60 years ago, 39,40 locally released growth factors in wounds have since been believed to be the main driving force behind wound healing. 41 Growth factors are thought to be responsible for inducing wound reepithelialization by promotion of the lateral migration of epidermal keratinocytes and then act as a chemoattractant to promote the inward migration of dermal fibroblasts (which remodel damaged dermis) and microvascular endothelial cells (which revascularize the neodermis). 42 Under this not-evidence-based assumption, more than 30 growth factors have been subjected to extensive preclinical and clinical studies alone or in combination over the past several decades. 30,43 Epidermal growth factor (EGF) first entered clinical trials for treating partial-thickness skin graft wounds in 1989. 44 Subsequently, numerous growth factor trials were conducted, including (i) EGF treatment for traumatic corneal epithelial defects, 45 chronically perforated tympanic membranes, 46 and advanced DFUs 47,48 ; (ii) basic fibroblast growth factor (bFGF) treatment for partial-thickness pediatric burn wounds, 49,50 second-degree burns, 51 and DFUs 52 ; (iii) acidic FGF treatment for partial-thickness burns and skin graft donor sites 53 ; (iv) granulocyte and macrophage colony-stimulating factor plus bFGF treatment for PU 54 and venous leg ulcers 55 ; and (v) platelet-derived growth factor-BB (PDGF-BB) treatment for chronic pressure and DFUs, 56 –59 just to mention a few. While most of the double-blinded trials initially reported promising clinical efficacies, only recombinant human PDGF-BB received approval from the U.S. FDA as a therapeutic for treatment of DFUs (becaplermin gel with 0.01% hrPDGF-BB [∼100 μg/mL]; Regranex™, Ortho-McNeil Pharmaceutical, Raritan, NJ). Following the initial approval, subsequent multicenter, randomized, and parallel trials showed that becaplermin gel improved wound closure modestly better than placebo; the studies found a 50% improvement rate with becaplermin gel treatment versus 36% with placebo. 60 Many argued that this level of improvement should not have been considered cost-effective for clinical practice.61– 63 In 2008, the FDA added a black box warning to becaplermin gel regarding an increased risk of cancer mortality in patients who require treatments of three (15 g) tubes or more. 64 This significant side effect may not be surprising to cancer researchers, since it had been known for years that overexpression of PDGF-BB (c-sis) or autocrine signaling of its viral form, v-sis, can cause cell transformation. 65 More intriguingly, the formulated concentration of PDGF-BB in becaplermin gel (100 μg/g gel) is several 1,000 times higher than the physiological PDGF-BB levels in human circulation, based on three previous reports that the mean PDGF-BB concentrations in normal human serum range from 10.56 to 17.5 ng/mL, compared to concentrations in normal human plasma of less than 1 ng/mL. 66 –68 Nevertheless, after repeated petitions by the manufacturer, the FDA has recently removed the cancer warning from Reganex, 69 based mainly on opposing evidence from two studies. 70,71 No other recombinant growth factors have since been granted FDA approval as wound healing therapeutics.
While the “growth factor-wound healing” paradigm has served as widely accepted guidance for preclinical and clinical wound healing studies over the past several decades, in retrospect, there has been little direct experimental evidence to support such statements as (conventional growth factors) playing an “essential role,” a “vital role,” a “requirement,” or a “driving force” in wound healing, although these phrases continue being used in wound healing literature. It is fair to say that the march to develop growth factor drugs promoting wound healing has been arduous, expensive, and unproductive. Revisiting the preclinical studies on hrPDGF-BB that ultimately led to FDA approval of becaplermin gel, we find that numerous laboratories participated in those studies and investigated a wide range of wound healing parameters, including granulation tissue formation, epithelialization, contraction, and wound strength. The researchers used animal models, including a guinea pig partial- and full-thickness wound model, 72 –74 a pig partial- and full-thickness wound model, 75,76 a normal or diabetic (streptozotocin [STZ] induced) rat excision wound model, 77 a rat implantation wound model, 78,79 a rabbit ear skin excision wound model, 80 –83 a rat corneal wound healing model, 84,85 and a normal and db/db diabetic mouse wound model. 86 –89 While results of these studies were highly variable, the consensus was that hrPDGF-BB/becaplermin gel (i) enhances granulation tissue formation and (ii) has little effect on wound epithelialization or wound closure and contraction, except in db/db mouse model and rabbit ear skin excision wound model. Notwithstanding the unclear evidence from these preclinical studies, clinical trials with becaplermin gel proceeded with a leap of faith assumption that enhanced granulation tissue formation may indirectly correct the damage caused by chronic diabetic wounds. The overall clinical outcomes with becaplermin gel treatments of DFUs showed variable and modest results, as previously mentioned.
Three potential hurdles for becaplermin gel/PDGF-BB to be a WDG
Joining wound healing research from a long-time growth factor laboratory of our own, we disbelieved the disappointing outcomes of growth factor therapies and decided to use PDGF-BB as a representative conventional growth factor to investigate what prevents it from being a WDG. We identified three possible hurdles. First, among human keratinocytes, dermal fibroblasts, and microvascular endothelial cells (the three cell types critical during wound closure), PDGF-BB only directly affects the PDGF-receptor-positive dermal fibroblasts, since neither keratinocytes nor endothelial cells express any PDGF receptors. 90 –93 Second, as previously mentioned, the co-presence of TGFβ3 in the wound could completely nullify the effectiveness of PDGF-BB. 35,90 Third, pathological conditions in chronic wounds, such as hypoxia (prevalent in wounds due to vascular disruption) and hyperglycemia in diabetics, further compromise the effectiveness of PDGF-BB. 90,94 For instance, vascular injury and obstruction in wounded tissue cause ischemia, creating a hypoxic wound microenvironment and impairing growth factor delivery. These three limitations, as schematically illustrated in Fig. 4 and restated here, could greatly compromise the effectiveness of PDGF-BB and perhaps other growth factors as well. These findings may explain why the several 1,000-fold higher concentrations of PDGF-BB (100 μg/g gel) in becaplermin gel compared to its physiological concentration in circulation (10–15 ng/mL) 66 –68 was needed to achieve a modest efficacy. An important question remains: What role do conventional growth factors play in wound healing and at what stage of the process? We speculate that growth factors play a critical role in dermal granulation and more importantly during the wound remodeling phase after wound closure.
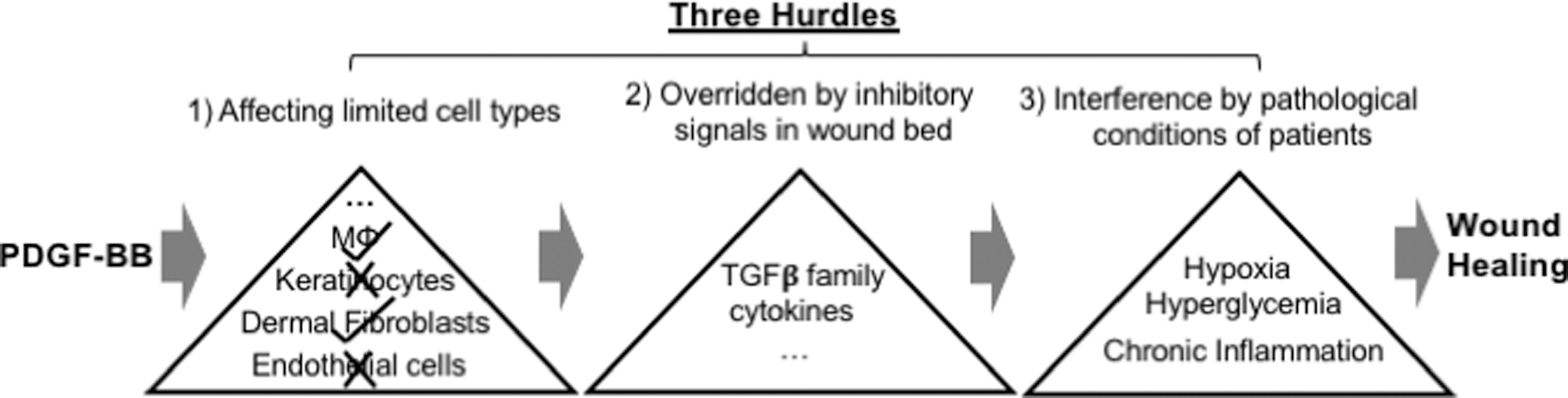
Three hurdles facing growth factor therapy. The three major skin cell types, keratinocytes, dermal fibroblasts, and endothelial cells, involved in wound healing are indicated. An example of a conventional growth factor, PDGF-BB, is as shown. Growth factors face three major hurdles, which prevent them from being primary drivers of wound closure: (1) only selective cell type(s) express the cognate receptor; (2) growth factors are sensitive to the antiproliferation and antimigration effects of TGFβ3, and (3) their efficacy is compromised under pathological conditions. PDGF-BB, platelet-derived growth factor-BB; TGFβ3, transforming growth factor-beta3.
Suggestion for a three-step establishment of a WDG
Among the five suggested requirements for a WDG, as listed in Fig. 3, the gene's necessity and sufficiency for wound closure in animal models are the foundational requirements. To successfully develop a WDG candidate, we propose three sequential lines of experimental approach: (i) in vitro studies of the mechanisms of action by a WDG candidate, (ii) a genetic animal model to prove the WDGs' essentialness for wound closure, and (iii) a wound healing animal model(s) relevant to the mechanism of human wound healing to evaluate the WDGs' sufficiencies for promoting wound closure. Finally, quantitative benchmarks of the wound closure measurements that define success or failure of a WDG candidate need to be established.
In vitro studies of a WDG candidate for molecular parameters
Cultured homogenous cells are necessary to define important parameters of a potential WDG product, such as its target cell types, binding target, binding affinity, binding specificity, signaling, and mechanisms of action. This information may be immediately available if a newly identified WDG candidate has already been well characterized from previous studies in a different context. For instance, EGF was originally described as a secreted peptide found in the submaxillary glands of mice and in human urine. PDGF-BB, however, was initially isolated from normal rat serum and was one of the first growth factors whose characterization in cultured cells led to the current understanding of signal transduction by many other growth factors. 95 What wound healing researchers would have needed to do was to study whether the same parameters are present in the skin cells involved in wound healing, before studies in animal models. In retrospect, investigations of PDGF-BB parameters in wound healing were not properly completed during earlier, preclinical studies, despite the FDA approval of becaplermin gel as a therapeutic drug for wound healing. For instance, earlier studies already showed that neither keratinocytes nor endothelial cells express mRNAs for any of the two PDGF receptors. 91 –93 Consistently, by Western immunoblotting analysis of the three main human skin cell types, our laboratory demonstrated that only dermal fibroblasts express the receptors for PDGF-BB. 91 The absence of PDGF receptors would prevent keratinocytes and endothelial cells from responding, at least not directly, to the biological effects of PDGF-BB/becaplermin gel, which were described as essential for promoting wound healing.
Genetic animal models to prove the WDGs' essentialness for wound closure
The necessity of a WDG candidate for wound closure can only be established using genetically engineered mouse (GEM) models. 96 Over the last several decades, GEM models have provided researchers with opportunities to investigate more than 100 genes for their potential importance in one or more aspects of skin wound healing. 97 In these mice, three kinds of full-thickness excision wounds are commonly utilized, as shown in Fig. 5, including (i) nonrestricted cover-held or “loose” wounds, (ii) adhesive bandage-held or “contained” wounds, and (iii) adhesive bandage-held and edge-sutured or “splinted” wounds. Among the three, 1 × 1 cm “loose” wounds reached complete closure first, by day 10 (upper panels). The “contained wounds” achieved complete closure by day 14 (middle panels). The “splinted wounds” showed the slowest wound closure around day 17 (lower panels). Quantitation of the wound closure and hematoxylin and eosin (H&E) staining measurements showed a significant difference in wound reepithelialization among the three types of wounds, in which splinted wounds showed the longest distance of keratinocyte migration like wound healing in humans. 98 For this reason, the splinted wound model has been preferred by many researchers, because it offers relatively more wound reepithelialization and less wound contraction in mice. Fuchs laboratory showed that keratinocyte growth factor-1 (KGF-1)/fibroblast growth factor-7 (FGF-7) gene knockout in mice had little effect on wound healing. 99 In essence, these results already disqualifies KGF-1 as a possible WDG, even though KGF-2/FGF-10 potentially could have substituted the role of KGF-1. On the other hand, Grose et al. showed that keratinocyte-specific knockout of β1 integrin in mice caused a retardation in wound reepithelialization. 100 Chmielowiec et al. reported that keratinocyte-specific knockout of c-met (hepatocyte growth factor receptor) caused a 30% delay in wound closure. 101 Moreover, Meyer et al. showed a 10–20% delay in wound closure in a keratinocyte-specific knockout model of the FGF receptor 1 and 2 genes in mice. 102 However, none of these gene knockouts altered the time for completion of wound closure. Because epidermal growth factor receptor (EGFR)-null mice do not survive beyond a few days after birth, Repertinger et al. examined wound healing using EGFR null- and wild-type skin grafted onto male athymic nude mice. They reported 60% delay in healing and 7-day delay in closure under these “hybrid” conditions. 103 Most of the findings served as supporting data for their clinical trials, as previously described, but none had advanced to clinical applications.
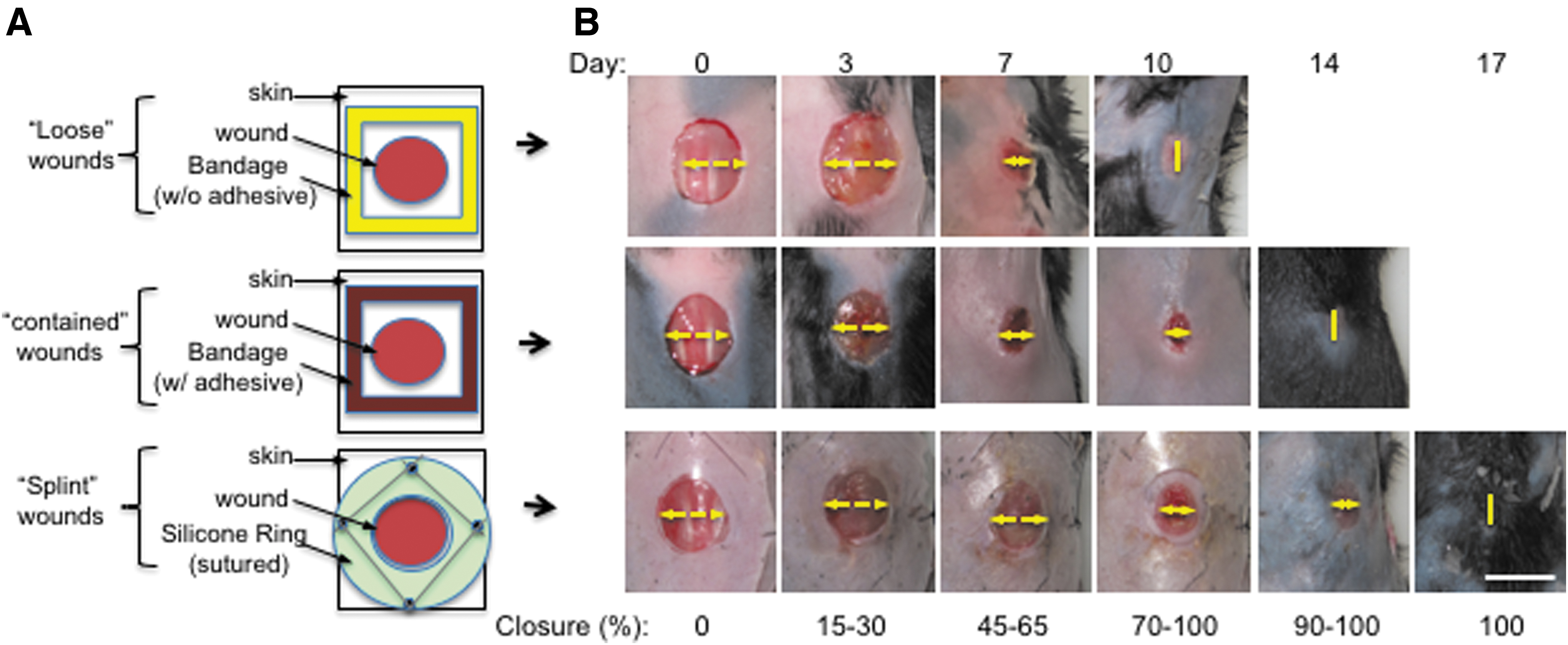
Comparison of wound closure rates among three types of commonly used wounds in mice.
Despite criticisms of mouse models due to the fundamental differences between mice and humans, GEM models, such as germline- or tissue-specific gene knockouts, remain an irreplaceable mammalian platform to identify the definitive functions for genes of interest. However, advancing directly to clinical trials based on mouse wound healing data may be premature and unproductive. Again, taking cancer research as an example, GEM has played an irreplaceable role in the progression of cancer research and contributed much to our conceptualization of how tumorigenesis takes place in humans. However, when it comes to translation of knowledge from mice studies to treatments of human cancers, we have witnessed four decades of delay from the initial announcement of the National Cancer Act in 1971 that optimistically announced to virtually eradicate cancer by 1976. While the complexity of human cancer may have surely been underestimated at the early times, it is hard not to blame the fundamental irrelevance of mice to humans in so many aspects of physiology. As famously stated by Rangarajan and Weinberg, “Mice are not small people.” 104 Among multiple genetic parameters, including metabolic rates, telomerase activities, susceptibilities to single versus multiple gene mutations, germ-line genomic alterations, and wide-scale mutations, the most significant challenges in using mouse models of human diseases are the species-specific differences, in terms of size, life span, organ morphology, and physiology. 105,106 Therefore, despite their indispensable advantages in biomedical research, the usefulness of GEM models for drug discovery has proven to be limited. Mouse models have additional disadvantages for developing skin wound healing drugs, because mice are loose-skin animals and heal skin wounds predominantly by the mechanism of wound contraction, instead of the mechanism of keratinocyte migration-led reepithelialization in human skin wound healing. Finally, researchers should also be aware of the extraordinarily costly and time-consuming business of drug development, which also carries a very low success rate. A joint study from a group of economic researchers at Tufts University, Duke University, and the University of Rochester reported a $2.6 billion cost estimate for developing a prescription medicine for FDA approval. If including the costs for postapproval research and development (tests for new indications or new formulations and long-term safety monitoring), the final cost is projected to be $3 billion. 107 –109
Using pig wound healing model(s) to evaluate the WDGs' sufficiencies for promoting wound closure
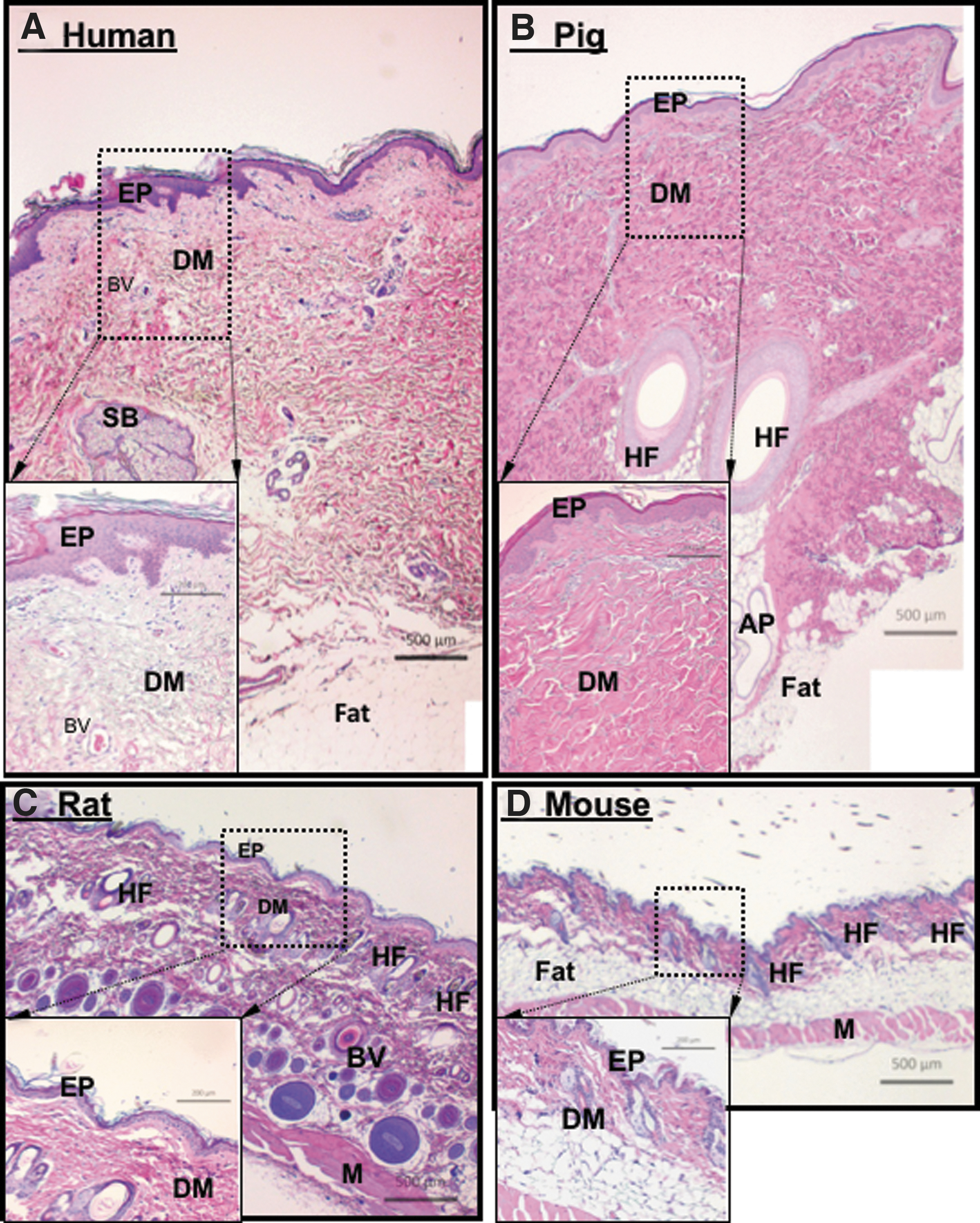
We suggest that the third step in testing the sufficiency of a WDG candidate is using a human-relevant animal model before rushing into the time-consuming and money-burning roller coaster of IND (investigational new drug) application, multiphase clinical trials, and FDA approval of an NDA (new drug application). 110 The good news is that a topically applied external drug for wound healing has an advantage over a systemically administered internal drug. For testing of topical drug candidates, pigs are widely considered the preclinical model of choice, despite limitations such as lack of transgenic models, higher costs, and more complex surgical and bandaging procedures. Pigs, like human beings, are tight-skinned animals and heal skin wounds with a larger component of reepithelialization (i.e., the lateral migration of keratinocytes across the wound bed) and a much smaller component of wound contraction than mice. Moreover, multiple groups of replicate wounds can be created in the same pig, another advantage of the pig model for testing topical agents. Randomized wound healing studies reported a high concordance of results between pigs and humans. 111,112 A comparison of H&E-stained sections of normal mouse, rat, domestic pig, and human skin shows a clear similarity between human and pig skin, in comparison to the rodent skin (Fig. 6). Recently, our laboratory has investigated, in more detail, how to properly utilize pigs to test a therapeutic candidate. These efforts allowed us to identify several previously overlooked wound healing parameters in pigs, which could determine the reproducibility and validity of experimental outcomes.
Differences in locations of the wounds
Previous pig wound healing studies that compared placebo with a treatment or even different dosages of the same treatment on wounds created on the same side of the pig torso may not have been the correct way of testing. Our investigation revealed that the skin, where wounds are made along the same side of a pig's torso, differ in elasticity, thickness, and hair follicle density from top to bottom and from left to right, even within a few centimeters apart. For instance, the epidermis becomes softer and more pliable going from top to bottom of the torso. As demonstrated in Fig. 7, two wounds that were located 4–5 cm apart on the same side of the torso, either vertically (A and B, panels a–d) or horizontally (C and D, panels e–h), showed significantly different closure rates. However, we found that two wounds created on opposite sides of the animal's torso, but at corresponding locations, underwent wound closure at similar rates (E, panels i–l). This finding may provide a solution for another long-recognized technical problem. Due to constant movements of the animal (standing and lying down, running around, and scratching against fences), exchange of added materials by diffusion among neighboring wounds on the same side of the torso, such as between drug-treated and placebo-treated wounds, could occur. We suggest that a therapeutic treatment and its placebo control should be applied to wounds that are created at corresponding spots on two opposite sides of the pig's body, to achieve more reliable and reproducible results. 94 Other factors, such as side for sleep preference, may further complicate comparisons of placebo- and drug-treated wounds and should also be considered in the study design. In all, the art is definitely more than skin deep.
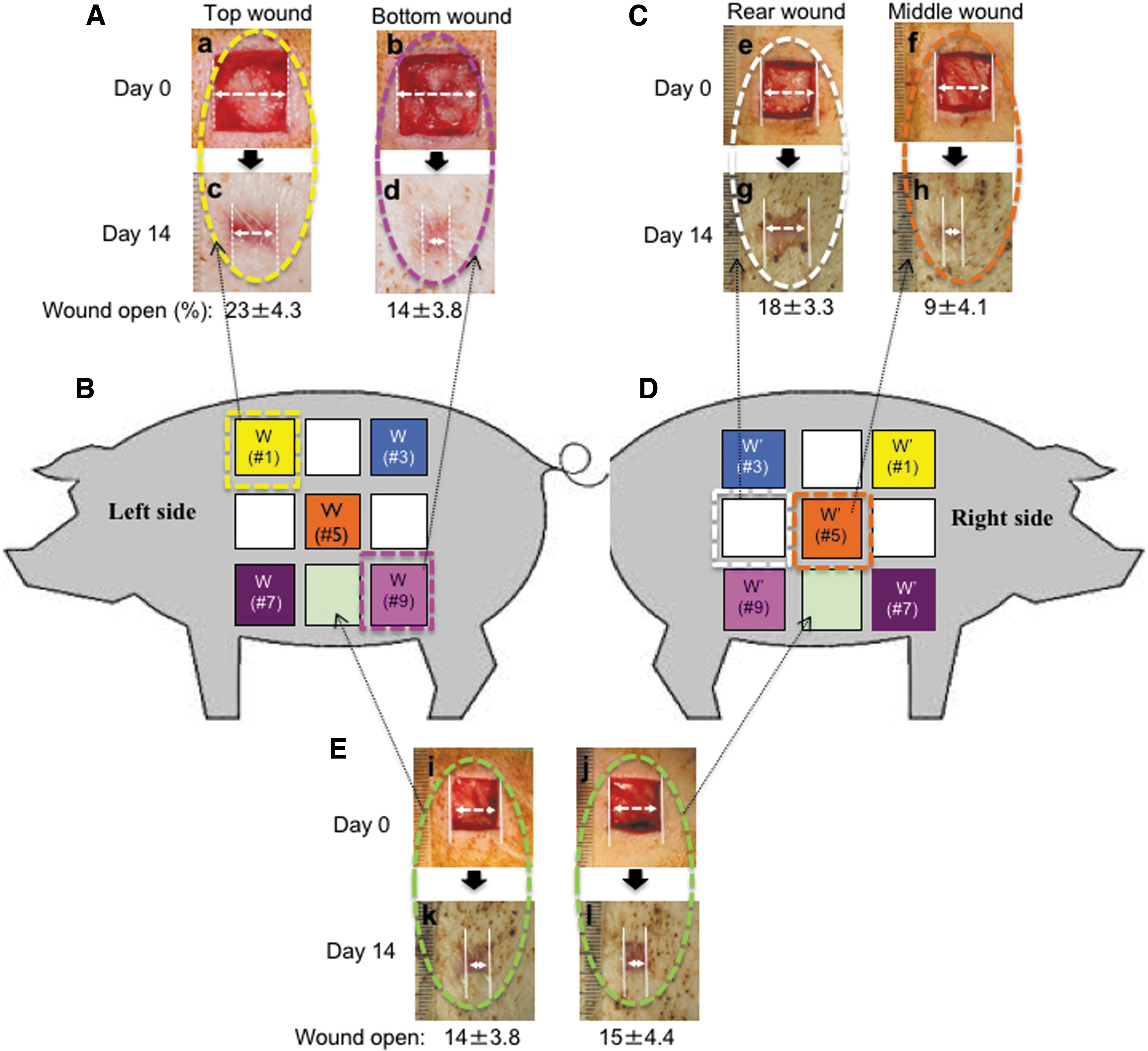
Wounds on the same side of a pig torso heal with variable rates.
Hyperglycemia and delay in wound closure
There has still been lack of any widely accepted animal model for PU and VU studies. In terms of DFU, chemically induced and transiently elevated plasma glucose levels in pigs are not equivalent to the chronic hyperglycemia in human diabetic patients and cannot cause a compatible level of pathological change in these animals. STZ enters through the β cell glucose transporter GLUT2, and causes the beta cells to undergo destruction by necrosis, resulting in higher circulating glucose levels in the animal. Eriksson's group first used an STZ-induced diabetic pig model in wound healing studies and reported 4 to 6 days of delay in skin wound closure following a 2- to 3-week period of STZ treatment. 113 Bergmann et al. compared partial-thickness wounds in normal pigs and STZ-treated pigs and reported little difference in wound closure rates. 114 Many similar studies followed suit with short-term STZ treatment and few presented direct evidence indicating changes in wound closure rates. 114 –121 We initially also followed the reported procedures of STZ-induced diabetic pigs, but were unable to detect any significant delay in wound closure. After repeated attempts of well-designed experiments and analyses, we came to the conclusion that the previously called (STZ induced) “diabetic pig wounds” lacked the most important clinical signatures of diabetic ulcers in humans—chronic hyperglycemia-associated delay in wound closure.
To reinvestigate whether or not there is indeed a correlation between duration of hyperglycemia and degree of delay in wound closure in pigs, we conducted time-course experiments over a period of 3 months following an initial one-time STZ injection. Six Yorkshire pigs (2–3 months old and 20–25 kgs in weight) were injected with STZ to induce high blood glucose levels, which were kept between 200 and 350 mg/dL by dietary control and insulin injection (if necessary) for 20, 45, and 90 days (during which the A1c levels rose from 4.1 to 5.5), before wound healing experiments. As illustrated in Fig. 8, 1.5 × 1.5 full-thickness wounds in a control pig closed around day 14, as expected. Wounds in pigs with a 20-day history of hyperglycemia did not show any detectable delay in wound closure. Interestingly, pigs in which hyperglycemia was kept for 45 and 90 days exhibited a 7- and 15-day delay in wound closure, respectively. This is the first validation of the common clinical observation that the greater the duration of hyperglycemia, the more pronounced the delay in diabetic wound closure. 94

Duration of hyperglycemia correlates with degree of delay in wound closure in STZ-treated pigs. The delay in wound closure was examined in pigs injected with STZ 20, 45, and 90 days before wound surgery.
A need for establishment of a chronic diabetic pig wound healing model for drug tests
Adult age and years of accumulated damage from pathological conditions are the signatures of chronic wounds in humans, during which many components for wound closure may have been damaged. Similar signatures of various chronic human disorders have long posed as major challenges for scientists attempting to create relevant animal models. In turn, this decade-long and still ongoing conundrum may have directly contributed to the overall low rate of drug development for chronic diseases. Among the three main types of chronic wounds in humans, VU, PU, and DFU, perhaps the most appropriate or relevant chronic wound animal model is the minipig model with an (minipig life span based) adult age and chronic hyperglycemia.
Pigs used in biomedical research belong to two main groups, domestic pigs and miniature pigs. Domestic pigs have rapid growth rates and grow from 1 kg at birth to 100 kg within 4 months (without any dietary control), and typically reach an adult weight of >200 kg within 1 year. The rapid growth rate and large size of domestic pigs pose a logistical problem for research, since 50 kg is the limitation for animal facilities on most U.S. university campuses. Due to this limitation, most previous wound healing studies had to expedite the development of hyperglycemia in their animal model and could not keep animals long enough to induce chronic hyperglycemia. Typically, these studies used animals of <3 months of age and chemically induced (acute) hyperglycemia for only 2–3 weeks. In humans, a majority (90%) of type 2 diabetes mellitus (T2D) patients exhibit an adult onset (∼40 years old) of disease and years, if not decades, of chronic hyperglycemia. If we take 15 years as the average lifespan for domestic pigs, 4-year-old pigs are reflective of 40-year-old human beings. Therefore, the farm pig model cannot feasibly meet the two critical parameters of T2D in humans: an adult age and chronic hyperglycemia (i.e., accumulated damages by tissue glycation). Unlike domestic pigs, however, miniature pigs grow to 10–25 kg within 4 months of age and reach their adult weight of ∼50 kg (fully grown) by the end of their first year of life. Since the life span of miniature pigs is 10–12 years, minipigs greater than 3 years old, which have been subjected to 1 year of hyperglycemia, may model the typical course of diabetes in humans. These numbers make minipigs more suitable for research projects studying chronic diseases such as diabetes. For example, Yucatan miniature swine look like traditional domestic pigs, except they are much smaller in size. Their light skin, relatively light hair coat, and low-fat content make these pigs an excellent candidate for topical drug studies. 96,112 Sinclair Bio-Resources (Columbia MO) offers adult and diabetic minipigs between 1 and 7 years of age with chronic diabetic conditions (induced with alloxan at 7 months of age). These animals have been used to study various complications of diabetes, such as kidney failure, but have not been fully characterized for wound healing studies. A limitation with these “real” diabetic minipigs is their very high cost (average cost of $8,000 per animal vs. $1,500 per normal control animal). A recent study by Abadir et al. used this adult age and chronic hyperglycemia minipig model (36-month-old animals with 29 months of alloxan-induced hyperglycemia) to study the role of angiotensin 2 receptor in wound healing. Unfortunately, these authors did not look into any diabetic wound parameters such as the degree of delay in wound closure. 122 We believe there is a need to fully characterize and establish a diabetic minipig model for wound healing research and drug screening, as it can be used to assess for sufficiency of a WDG drug candidate, and it is a relevant model for human disease.
Stress-triggered, keratinocyte-secreted, and exosome surface-bound heat shock protein-90alpha: a WDG candidate or not quite?
Previous wound healing research has covered a wide range of potential wound closure candidates, including mesenchymal stem cells, nucleotides such as micro-RNAs, proteins such as growth factors and cytokines, peptides such as angiotensin II, extracellular vesicles such as exosomes, and small molecules such as wnt pathway modulators, not to mention various kinds of bioengineered materials and devices, hoping to achieve the proposed capabilities of a WDG. While continued investigations may result in breakthroughs, the scope of this review does not allow us to discuss each of those possibilities in further detail. Nonetheless, to date, there has not been a single gene product(s) that has met all five of the requirements for a WDG, as previously illustrated in Fig. 3.
Under the microscope, keratinocyte migration occurs almost immediately following skin injury, whereas inward migration of dermal cells is not detected until 4 days afterward. 36 One possible mechanism is that skin injury triggers keratinocytes to produce and deposit a laminin-5 isoform into the basement membrane of the provisional matrix, so that keratinocytes themselves migrate into the wound bed within hours. 123 How is the timing of these sequential events regulated? It is known that TGFβ family cytokines act as negative regulators of cell proliferation and migration. In Smad 3 knockout mice, wound epidermal cells are unable to transduce the TGFβ signal, resulting in accelerated reepithelialization of wounded skin. 124 A 2006 study from our laboratory provides a possible explanation. Our findings suggest that the appearance of TGFβ3 in the wound selectively halts dermal cell migration and proliferation and lets keratinocyte migration proceed first. 36 It is known that a migrating cell does not proliferate and a proliferating cell does not migrate. This antagonistic behavior of cell migration and cell proliferation was first referred to as the migration/proliferation dichotomy or “Go-or-Grow” behavior, 125 and has been supported by numerous in vitro and in vivo observations. 126 –129 These studies clearly indicate that proliferation and migration are mutually exclusive behaviors.
During wound closure, cell migration appears to precede and subsequently trigger cell proliferation. In their NEJM article, Singer and Clark wrote, “One to two days after injury, epidermal cells at the wound margin begin to proliferate behind the actively migrating cells. The stimuli for the migration and proliferation of epidermal cells during reepithelialization have not been determined, but several possibilities exist. The absence of neighboring cells at the margin of the wound (the ‘free edge’ effect) may signal both migration and proliferation of epidermal cells.” 30 Our view is that, as cells at the wound edge move toward the wound bed, they inevitably leave empty space behind them. Cells adjacent to the empty space lose contact inhibition with the front migrating cells, and proliferation is induced. The stimuli responsible for this later stage of cell proliferation are likely from diffusion of plasma growth factors from surrounding unwounded blood vessels, where TGFβ3 levels are low or undetectable. Further studies are needed to provide more definitive evidence for this important concept. On the other hand, more recent studies suggested “collective cell migration,” instead of separate individual cell migration at the wound edge. These studies showed that wound keratinocytes migrate as a coherent layer with a loosened adhesion between cells. Then, cell proliferation occurs behind the migrating sheet to fill the empty space. The authors argue that collective cell migration during wound healing is relevant to the wound clinic. 130 –134
When we launched a search for potentially new WDGs over a decade ago, we argued that the WDGs must be able to drive both epidermal and dermal cell migration against TGFβ3 inhibition in the wound bed. Furthermore, we reasoned that this WDG product(s) does not come from HS following wounding, because TGFβ3 in HS inhibits human dermal cell migration and growth. 34,35 Instead, we postulated that the WDG candidate(s) originates from secreted factors by stressed epidermal and dermal cells around the wound edge. Following collecting several liters of the cells' serum-free conditioned media, protein purification led to identification of the secreted form of heat shock protein-90alpha (Hsp90α). 134,135 We demonstrated that the secreted form of Hsp90α is able to overcome the three hurdles that rule out most conventional growth factors as WDGs. First, secreted Hsp90α is a common promotility factor for all three types of human skin cell types involved in wound healing because all three cell types express a compatible level of LRP-1 (LDL receptor-related protein-1), the receptor required for extracellular Hsp90α signaling. 90,129 Second, even in the presence of TGFβ3, secreted Hsp90α remains equally effective in promoting migration of all three types of human skin cells. 94,135,136 To our knowledge, secreted Hsp90α is the first molecule identified that is able to override the inhibitory effects of the TGFβ family cytokines. 137 Third, while hyperglycemia reduces growth factor-stimulated human skin cell migration, secreted Hsp90α not only promotes cell migration in normal glycemic conditions but also “rescues” migration of cells cultured in hyperglycemic conditions. 94
The next question is whether or not keratinocyte-secreted Hsp90α is necessary for wound closure. Dr. Udono's group generated a unique mouse model where the intracellular function of Hsp90α is selectively removed by a C-terminal deletion to prevent its binding to client molecules. However, the extracellular functional part within the Hsp90α-Δ mutant, the so-called F-5 region, remained intact. 90,98 These mice are phenotypically indistinguishable from their wild-type counterparts 138 and, therefore, allowed us to specifically test Hsp90α's non-chaperone and extracellular functions during wound closure. We found that the chaperone-defective Hsp90α-Δ mutant mice showed wound closure rates similar to Hsp90α-wt mice, suggesting that the secreted Hsp90α-Δ deletion mutant protein is fully functional in promoting wound closure. However, when we selectively inhibited the extracellular Hsp90α-Δ protein function by adding a neutralizing antibody to wounds, a significant delay in wound closure was detected in these mice, indicating the necessity for secreted Hsp90α. 98 To test if secreted Hsp90α alone is sufficient to drive normal and chronic wound closure, we demonstrated that topical application of human recombinant (hr) Hsp90α protein or its F-5 peptide accelerates full-thickness excision, burn, and diabetic skin wound closure in mouse and pig models. 90,94,98,139 These findings indicate that keratinocyte-secreted Hsp90α meets the two most critical qualifications of a WDG of being necessary and sufficient for wound closure. In addition, it is secreted under conditions of cellular stress, resistant to the inhibitory functions of TGFβ and effective on multiple cell types. Our current understanding of secreted Hsp90α in leading and expediting the initial phase of wound closure is schematically depicted in Fig. 9. We have been actively pursuing the possibility that keratinocyte-secreted Hsp90α is a critical driving force during the early phase of wound closure.

A model of how secreted Hsp90 promotes reepithelialization and recruits dermal cells into the wound bed during wound closure. (Step 0) Uninjured intact skin with little detectable TGFβ3, cell migration, or stress; (Step 1) injury triggers release of TGFβ3, an immediate immotile to motile transition of keratinocytes and release of conventional growth factors. Growth factors are not able to recruit dermal cells to the wound bed due to the presence of TGFβ3; (Step 2) stressed keratinocytes at the wound edge release/secrete Hsp90α (round dots). Secreted Hsp90α reaches the threshold concentration of >0.1 μM and drives inward migration of dermal cells; (Step 3) keratinocyte migration closes the wound, TGFβ3 disappears, and the migrated dermal cells start to remodel the wound (taken with permission of all the authors for Ref. 90). Hsp90, heat shock protein-90.
Summary
Based on the current status quo, the outlook on the development of new, effective wound healing therapeutics remains pessimistic. The demand for wound healing drugs to rescue the aberrant healing process, which may consist of multiple dysfunctional components, poses a significant obstacle. It is conceivable that no single molecule or WDG is capable of promoting wound closure. On the other hand, R&D of multicomponent therapeutics presents additional technical and economic challenges. Finally, the ongoing deficiency of government funding for wound healing research will continue to leave researchers struggling and patients devoid of optimized therapies.
Take-Home Messages
The disproportionately low level of government support for wound healing research, in comparison to support for cancer research, has been a major rate-limiting factor that prevents discovery of wound healing therapeutics.
Wound healing drugs must overcome additional and greater obstacles in comparison to cancer drugs. While a cancer drug needs its cellular target only to be necessary during cancer progression, a wound healing drug must be a factor that is both necessary and sufficient to reactivate a halted repair process to close the wound.
Wound closure should be the primary focus for drug development.
Conventional growth factors are not WDGs for promoting wound closure, although they have been recognized as such for decades.
Animal models that share similar systemic and chronic parameters of human disorders, such as adult onset and chronic hyperglycemia for T2D patients, will be critical to break the glass ceiling of the decade-long paucity of new wound healing treatments.
Footnotes
Authors' Contributions
W.L. planned, conceptualized, and orchestrated the writing; D.G.A. verified the accuracy and clinical support of key statements; X.T. made figures; M.H. helped the writing and critically edited the article; C.C. helped making figures and gathering all references; A.B. and K.O. synthesized the data; and M.C. helped putting all items in places and checked for correctness.
Acknowledgments and Funding Sources
We thank those who gave us permissions to use their published data in this review article. This study is supported by NIH grants GM066193 and GM067100 (to Wei Li) and grant W81XWH-1810558 from the Congressionally Directed Medical Research Program (to Mei Chen).
Author Disclosure and Ghostwriting
Authors claim no competing financial interests. All contents of this article were constructed and written by the authors listed. There were absolutely no ghostwriters used for the writing.
About The Authors