
Abstract
Objective:
To design and validate a novel murine model of full-thickness (FT) vaginal wound healing that mirrors postinjury tissue repair and underscores the impact of estrogen signaling-driven healing kinetics, inflammation, and neovascularization.
Approach:
Five-week-old female CD1 mice were subjected to two 1-mm FT wounds. To assess wound healing kinetics, vaginas were harvested at 6, 12, 18, 24, 48, and 72 h and 7 days postinjury. Wounds from all time points were analyzed by hematoxylin and eosin and trichrome to, respectively, assess the rate of wound closure and tissue deposition. Inflammatory leukocyte (CD45), neutrophil (Ly6G), and macrophage (F480 and CD206) infiltration was examined by immunohistochemistry (IHC) and the resulting anti-inflammatory M2 (CD206)/total (F480) macrophage ratio quantified. Neovascularization (CD31) and estrogen receptor-α (ERα) expression levels were similarly determined by IHC.
Results:
We observed rapid healing with resolution of mucosal integrity by 48 h (p < 0.05), and overall neutrophils and polarized type 2 macrophages (M2) apexed at 12 h and reduced to near control levels by day 7 postinjury. Tissue repair was virtually indistinguishable from the surrounding vagina. CD31+ vessels increased between 12 h and day 7 and ERα trended to decrease at 12 h postinjury and rebound at day 7 to uninjured levels.
Innovation:
A proof-of-concept murine model to study vaginal wound healing kinetics and postinjury regenerative repair in the vagina was developed and verified.
Conclusion:
We surmise that murine vaginal mucosal repair is accelerated and potentially regulated by estrogen signaling through the ERα, thus providing a cellular and molecular foundation to understand vaginal healing responses to injury.

Introduction
Vaginal atresia, congenital adrenal hyperplasia, obstructive Mullerian anomalies, and Mayer–Rokitansky–Kuster–Hauser syndrome (vaginal aplasia) are prevalent disorders in young girls and adolescents, which ultimately require vaginal surgery with an incidence that ranges from 1/5,000 to 1/2,500. 1 –3 Notably, surgical vaginal reconstruction leads to increased tissue fibrosis within the mucosal canal in 73% of patients, 4,5 which inevitably causes lifelong scarring sequelae or requires reoperation. What determines whether postsurgical vaginal wound healing undergoes regenerative or fibrotic tissue repair is not known and mechanisms that govern vaginal wound healing remain to be elucidated, which requires development of robust in vivo models.
Previous research in vaginal wound healing has mainly utilized rats, rabbits, and guinea pigs as model animals to simulate full-thickness vaginal wounds 6 –8 where initial work compared full-thickness (FT) wounds (injuries that breach the muscularis) between vaginal and dermal tissues. 9 For instance, Abramov et al. showed that FT vaginal wounds in rabbits follow the same histological sequence of dermal wound healing, but at an accelerated pace, 9 which equally leads to a similar fibrosis pattern. Interestingly, while the mRNA profiling of FT vaginal wounds followed similar gene regulation as dermal wounds, the mucosal wound mRNA profiles reflected molecular differences underscoring reduced proinflammatory (TNFα, IL1α, and IL1β) and profibrotic (TGFβ) gene expression. 10 However, rabbits do not experience an estrus cycle, thereby undermining the comparison with human vaginal tissue repair pathophysiology. 9 On the other hand, Balgobin et al. 7 systemically replaced estradiol in oophorectomized guinea pigs and created FT vaginal injuries to study the effects of estrogen on vaginal tissue at 4 and 21 days postinjury. While estradiol appeared to increase smooth muscle and thickness of the vagina with increased total and cross-linked collagen, the authors did not assess vaginal healing kinetics or inflammatory and angiogenic profiles. Furthermore, while rabbits and guinea pigs provide adequate vaginal size to create full-thickness injury models, their use is limited for in vivo applications given the cost, federal regulations, and lack of transgenic lines. Moreover, subsequent studies on FT vaginal (epithelium and stroma) injury models in rats from the introitus to the cervix have been previously described. 11 Although these studies demonstrated the role of acute postoperative administration of vaginal estrogen on epithelial and stromal healing in the context of biomechanical properties of the healed vaginal tract, the impact of immune cells and healing kinetics was overlooked.
Recent literature has highlighted the effects of estrogen on vaginal wound healing where postoperative vaginal estrogen increases tissue compliance through promotion of matrix components such as collagen types I and III. 11 The observations are supported by the strong effects of estrogen on the vaginal mucosa such as the increase in cellular proliferation and differentiation. 12 Notably, other studies showed that low-dose conjugated equine estrogen (Premarin; Pfizer, New York, NY) could improve vaginal weight through increased vaginal thickness in ovariectomized rats while enhancing epithelial layer proliferation and inhibiting mucosal proteases. 6 In this study, we sought to understand the dynamics of estrogen signaling and vaginal physiological responses to injury using an innovative, in vivo murine model of mucosal wound healing to investigate the mechanistic relevance of estrogen in vaginal tissue repair.
The primary objectives of this project were, first, to establish a murine model of FT vaginal wound healing to define vaginal cell and molecular reactions to injury in the context of kinetics, inflammatory burden, and angiogenic events. Second, in view of recent literature, which highlights the effects of estrogen on progressive stages of vaginal wound healing, including proliferation, epithelial closure, and matrix turnover, 6,7 we evaluated the significance of estrogen receptor α (ERα) density during wound healing.
Clinical Problem Addressed
The high incidence of postsurgical fibrosis is at odds with the notion that vaginal mucosa heals regeneratively. There are no established postoperative procedures to reduce vaginal tissue fibrosis. Therefore, understanding how the vagina heals and uncovering the mechanisms that dictate the direction of vaginal tissue repair toward either regenerative or fibrotic wound healing are fundamental to improve patient care and surgical outcomes.
Materials and Methods
Ethics statement
Female CD1 pre-estrus mice between 4 and 5 weeks old were purchased from the Center for Comparative Medicine at Baylor College of Medicine and used for these experiments according to the ARRIVE (Animal Research: Reporting In Vivo Experiments) guidelines. 13 Animals were socially housed under a 12-h light–12-h dark cycle with free access to standard food and water. These studies were carried out in strict accordance with recommendations in the Guide for the Care and Use of Laboratory Animals of the National Institutes of Health (NIH). All animal research was conducted according to protocols approved by the Institutional Animal Care and Use Committee (IACUC) at Baylor College of Medicine (assurance number AN-7475).
In vivo FT vaginal wound
We first established and validated an in vivo, FT vaginal wound model. Mice were sedated using isoflurane at a rate of 0.5 L/min and underwent a bilateral, FT vaginal wound broaching the vaginal smooth muscle layer using a 1-mm biopsy punch at the dorsolateral aspects of the vaginal canal at 500 μm distal to the introitus by inverting the vaginal mucosa using smooth flat forceps (Fig. 1). Injuries were marked using a 29-gauge insulin needle tip dipped in green tissue dye (Davidson's Histology Tissue Dye, Catalog No. 1008-P; Bradley Products, Bloomington, MN). Mice were allowed to recover after surgeries and heal postoperatively. Mice were euthanized at 0, 6, 12, 24, 48, and 72 h and 7 days postinjury. Vaginal tissue was explanted and fixed in 10% neutral buffered formalin (n = 2–4 animals per time point comprising biological and technical replicates).
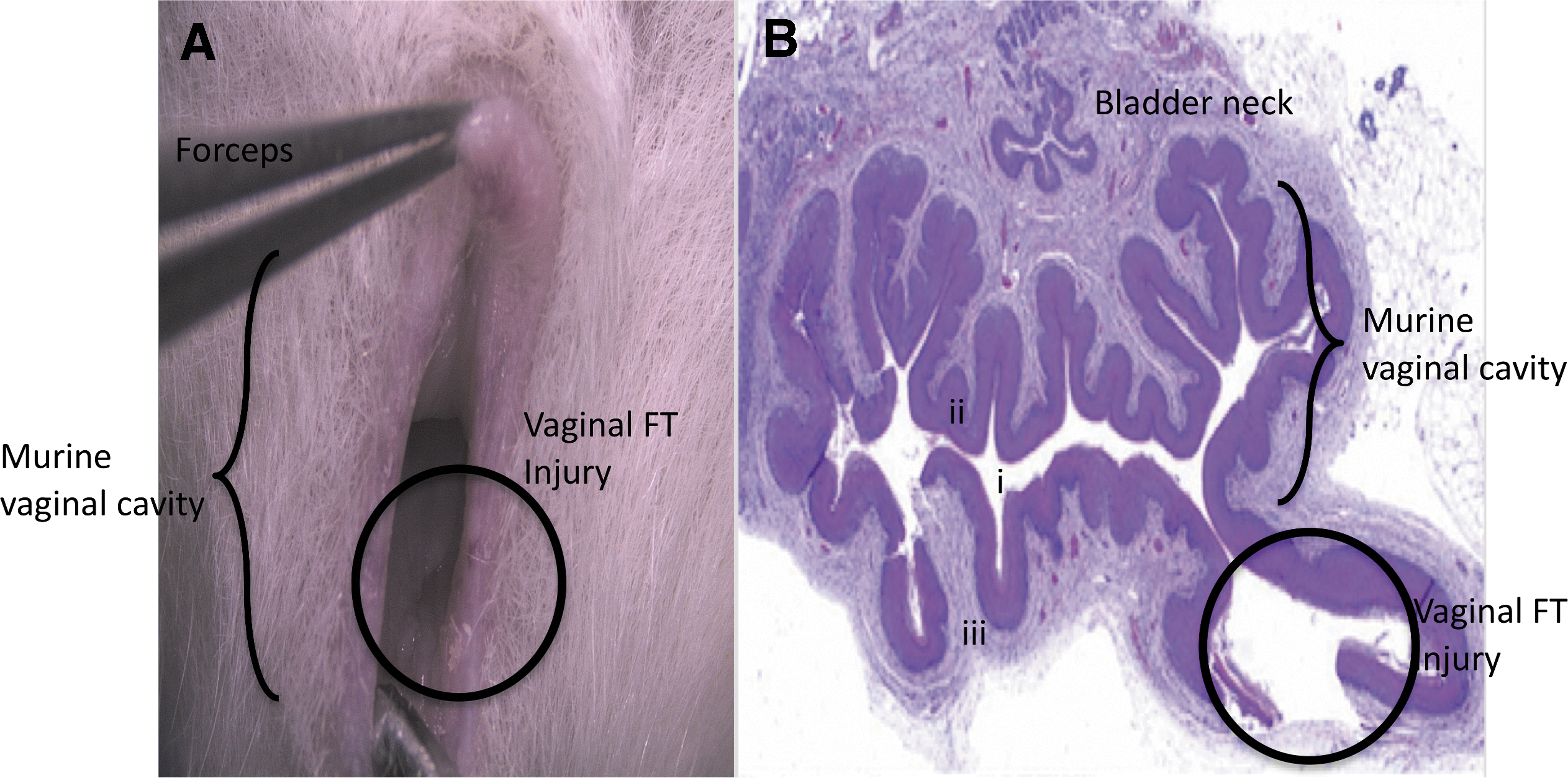
Murine vaginal wound model.
Histology of wounds
Formalin-fixed paraffin-embedded (FFPE) sections were cut to 5-μm thickness, rehydrated, and stained with hematoxylin and eosin (H&E) and trichrome staining on a Leica autostainer (ST5020). Vaginal tissue sections were double-blind examined by two pathologists for assessment of vaginal injury depth for validation of our injury model.
To determine wound healing kinetics, H&E-stained wound sections were imaged at 5 × using a Leica DM2000 microscope with a Leica DFC450 camera (Buffalo Grove, IL) and Leica Application Suite X (LAS-X). Measurements (in μm) of the epithelial gap between encroaching epithelial margins were taken using LAS-X software. ImageJ software (NIH, Bethesda, MD) was used to determine total cell density within the injured area of the submucosa using four 40 × images taken around the injury, which was determined to be equal across treatment groups (Supplementary Fig. S1). To evaluate extracellular matrix deposition, 5-μm sections were stained with trichrome and four 40 × images were taken per slide and quantified using ImageJ as the area of submucosal tissue above a threshold, which remained constant for all images.
Immunohistochemistry staining
Immunohistochemistry (IHC) was used to evaluate angiogenesis and inflammation following FT wounding. FFPE sections were cut to 5 μm and rehydrated, followed by antigen retrieval using either citrate buffer at pH 6.0 or low-pH antigen solution (K8005; Agilent, Santa Clara, CA) and the PT Link antigen retrieval system (Dako). IHC was completed either by hand or by using an autostainer (Dako Autostainer Link 48; Agilent). The following primary and secondary antibodies were used for IHC analysis: CD31 (550274, 1:100; BD Pharmingen, San Jose, CA), CD45 (ab10558, 1:2,000; Abcam, Cambridge, MA), CD206 (ab64693, 1:500; Abcam), F4/80 (MF480000, 1:5,000; Thermo Fisher, San Francisco, CA), ERα (ab32063, 1:50; Abcam), Ly6G (551459, 1:200; BD Pharmingen), anti-rabbit (K4009; Agilent), and anti-rat (D35-110; GBI Labs, Bothell, WA). 3,3′-Diaminobenzidine (DAB) or aminoethyl carbazole (AEC) was used to detect positive staining and hematoxylin was used as a counterstain. Four 40 × images per slide were taken using a Leica DM2000 microscope outfitted with a Leica DFC450 camera using Leica Application Suite X software. ImageJ was used to count positive cells within the submucosal tissue, reported as number of cells/submucosal area.
Statistical analyses
Microsoft Excel (v16.22) and Prism (version 6.0e) were used to conduct all statistical analyses. One-way analysis of variance with multiple comparisons post-testing between groups was performed using Dunnett's correction to determine the significance between noninjured control tissue and injured tissue at different time points. Tukey's tests were performed to determine differences between time points. Sample size was considered adequate to determine the difference from controls, for 0 and 48 h postinjury for epithelial gap and 0 and 12 h postinjury for cellular fluctuations, calculated for 95% confidence and power of 80%. Outliers were removed using Q test analyses.
Results
Murine model validation for intravaginal FT wounding
In this study, we established a murine, FT, vaginal wound healing model using an intravaginal minimally invasive approach. One-millimeter punch biopsy wounds were created in the vagina. Grossly, H&E staining of the vaginal cross section at the injury site revealed tissue breaches extending through the epithelial, submucosal, and smooth muscle layers (Fig. 1B), which were consistent across all the animals tested.
Vaginal wounds in a murine model are regenerated by 48 h after injury
We observed that in the majority of mice evaluated, the epithelial layer of vaginal tissue is completely closed and regenerated by 48 h postwounding (Fig. 2A, B). The epithelial gap was widest (p < 0.05) at 12 h and showed progressive decrease between 12 and 72 h. The submucosal tissue layer regenerated more slowly, with large tissue breaches persisting until 72 h postwounding. Despite regeneration of the submucosal tissue defect at 7 days postinjury, with healing indistinguishable from the surrounding uninjured vaginal tissue, there were no statistically significant differences in the total (quantified) collagen presence over time in the injury reaction area (Fig. 2A, C). See Supplementary Figure S1 for H&E and trichrome image acquisition localization and quantification of uninjured vaginal tissue and vaginal tissue at 12 h and 7 days postinjury.

Murine vaginal wound healing.
Inflammation is resolved and vascular lumen density is restored by day 7 postwounding
We analyzed inflammatory cellular infiltration and vascular lumen density at 12 h and 7 days postinjury as these were the time points that corresponded to highest and minimal epithelial gaps, respectively. We compared inflammation and vascular density at 12 h and 7 days with that of the uninjured control, no injury (NI) vaginal tissue.
We first evaluated angiogenesis by staining for CD31, a marker for endothelial cells. By counting the number of vessels (within the submucosal area) that were positive for CD31, we found that there was a steady, but nonsignificant, increase in the number of CD31+ vessels between the control tissue and injured tissue both 12 h and 7 days postinjury (Fig. 3A, B).

Inflammatory and angiogenic changes over time in murine vaginal wound resolution.
We next quantified the overall cellular inflammatory burden and macrophage subpopulations using IHC. CD45 was used to reveal total leukocyte populations near the FT wound, and Ly6G was used to detect infiltrating neutrophils. Following tissue injury, we observed that while the number of CD45+ cells was significantly decreased around the wound bed (p = 0.0061), the population recovered by 7 days postinjury (Fig. 3A, C). We noted the opposite pattern for first-responder neutrophil Ly6G+ cells, with the greatest Ly6G+ influx seen at 12 h postinjury (p = 0.0044), which decreased to near-uninjured tissue levels at 7 days postinjury (Fig. 3A, D). Moreover, the total F480+ macrophage population and anti-inflammatory CD206+ M2 macrophages exhibited a similar trend as the overall CD45+ leukocyte population with a significant decrease at 12 h postinjury (CD206, p = 0.0131), which returned to nearly uninjured levels by 7 days (Fig. 3A, E, F). The ratio of M2 (CD206+)/total macrophages (F480+) showed opposite trends and increased markedly at 12 h postinjury and decreased to near-uninjured levels at 7 days postinjury (Fig. 3G). Taken together, these data support the concept that there is a population of resident vaginal leukocytes (CD45+) and that upon injury, there is an acute and rapid influx of neutrophils (Ly6G+) to the vaginal space. Although the overall macrophage population decreases at 12 h, the M2 burden appears to increase at 12 h and supports a shift toward a prohealing environment based on F480 and the CD206:F480 ratio at the earlier time point.
Estrogen receptor expression levels change during vaginal wound healing
To investigate whether estrogen signaling plays a role in vaginal wound healing, we compared ERα levels between uninjured controls and mice at 12 h and 7 days postinjury. We found that at 12 h postinjury, there was an almost 60% reduction in the number ERα-positive cells within the submucosa (Fig. 4A, B), which returned to uninjured control values at 7 days postinjury, thus revealing a transient injury-driven reduction of ERα signaling. This suggests that while estrogen is known to play a role in vaginal wound healing, signaling through ERα could be transient in the healing process since ERα levels returned to uninjured control levels by 7 days postinjury.

Estrogen plays a role in murine vaginal wound healing. ERα density measuring vaginal wound resolution between uninjured mice and mice at 12 h and 7 days postinjury, respectively, illustrated by IHC images
Our collective data support two important and novel concepts, namely that the 1-mm, murine, FT vaginal wound heals in an accelerated and regenerative manner by day 3 postinjury and that injury impacts inflammatory and angiogenic cell populations, which could be influenced by estrogen signaling.
Discussion
The present study established an unappreciated wound repair model of murine vaginal mucosa, which uncovers unique healing kinetics, inflammatory patterns, and angiogenic profiles that connect estrogen signaling with vaginal responses to injury.
Model, kinetics, and extracellular matrix
We devised and validated a minimally invasive intravaginal approach of FT vaginal wounding, which enables a robust means to inquire how the vagina and other mucosal tissues such as oral and gastrointestinal microenvironments heal. The model is innovative, minimally invasive, and cost-effective while circumventing the limitations of previous larger animal models, opening new options for state-of-the-art, genetically engineered research approaches and generation of unique reagents for virtually unlimited experimental manipulations. We also demonstrate that murine vaginal wound healing occurs in an accelerated manner with primary epithelial closure long before submucosal integrity is restored. This mirrors established knowledge regarding healing in other mucosally lined tissues, with an absence of the hyperproliferative epithelial appearance seen in dermal tissue repair.
Role of inflammation and angiogenesis in wound healing
When a new wound is created, destruction of the extracellular matrix (ECM) and introduction of microbes lead to an influx of polymorphonuclear leukocytes (PMN or neutrophils), which compose the primary responders of innate immunity. The initial influx of neutrophils to the site after wounding with subsequent reduction follows established patterns of inflammatory postwounding behavior. Neutrophils are recruited to the site immediately after injury to reduce tissue damage associated with the release of proinflammatory mediators, 14,15 reactive oxygen species, proteases, and antimicrobials. Consistent with this notion, we demonstrated that the overall cellular inflammatory burden from neutrophils was highest at 12 h postinjury and returned to near-uninjured tissue levels by 7 days postwounding. Interestingly, we did note decreases in total leukocytes and total and anti-inflammatory M2 macrophages at 12 h, with the levels returning to near-uninjured tissue levels by day 7. Our findings are further supported by other tissue injury models 16,17 that have observed a similar shift in leukocyte:neutrophil ratios with comparable M1 to M2 macrophage polarization after injury and over time. Moreover, M1 to M2 polarization from early phase wound repair to later healing processes in dermal wounds has been confirmed in alterations implicated in the pathogenesis of chronic wounds. Furthermore, although neutrophil decontamination of the wound bed in the early phases of repair is critical, their efficient removal by macrophages is vital to nondisordered wound healing. 16 Importantly, neutrophils are noted to be absent or markedly reduced in nonscarring tissues such as fetal skin, oral mucosa, and Acomys nonscarring mice. 17,18 Notably, emerging evidence suggests that macrophages are the central arbiters in all stages of the wound healing response, which exhibit remarkable polarization plasticity present at varying levels during all stages of wound healing. 19 The initial proinflammatory environment at 12 h postinjury observed in our murine model of vaginal wound healing undergoes polarizing M1 to M2 shifts into a prohealing stage by 7 days postinjury.
Role of angiogenesis in wound healing
The intertwined nature of inflammation and angiogenesis can be understood through the dynamics of our vaginal model of wound healing. Migration and proliferation of endothelial cells within the proliferative phase of the vaginal tissue repair responses to injury are critical to formation of new capillary beds for nutrient and waste transport and subsequent resolution of the wound site. During the wound healing cycle, macrophages release vascular endothelial growth factors to recruit endothelial cells into the granular tissue. 20,21 This continues throughout the proliferative phase and into the latent stages of vascular remodeling, with wound healing progressing in coordination with an increase of M2 macrophages. While we observed no significant rise in the number of CD31+ vessels near the wound, there is a slight upward trend between 12 h and 7 days, suggesting that similar to other wound healing models, angiogenesis follows the inflammatory repair phase, which promotes wound resolution.
Wound healing as an estrogen-dependent phenomenon
The role of estrogen is a potent anti-inflammatory mediator, which has been studied in specific mechanisms of wound healing that include angiogenesis, M1 to M2 macrophage polarization, and fibroblast inhibition. Consistent with those studies, patients undergoing hormone replacement therapy exhibit improved cutaneous wound healing, through reduced PMN recruitment, decreased elastase production, and subsequent increase in collagen deposition. 22 –25 Exposure to physiological levels of 17β-estradiol (E2) reduces PMN migration by inhibiting chemotaxis through an estrogen receptor-dependent mechanism. 26 In this context, estrogen receptors (ERs) are important in many cellular functions, and the expression levels of these receptors concomitantly vary with systemic estrogen levels in a dose-dependent manner. 27,28 However, what remains to be elucidated is how the fluctuations in endogenous estrogen affect ER sensitivity, location, and/or density, which are known to be affected by injury. 29 In this study, we found that ERα density decreases immediately after injury and rebounds to uninjured levels when injured tissues are regenerated at day 3. Our collective data are supported by previous studies, which show that ERα density is inversely correlated with wound healing in rodent vaginal tissue. 6,30 –32 Future studies will not only evaluate the connection of ERα density but also unveil how estrogens regulate vaginal mucosa responses to injury as they relate to inflammation, angiogenesis, and collagen deposition. In sum, the emerging concepts from our work and that of others surmise that ERα signaling is mechanistically influenced by injury, which warrants further exploration to leverage our discoveries on the bench with bedside applications that improve regenerative wound repair, standard of clinical care, and patient's quality of life.
Conclusion
The generation, validation, and study of a mouse model for FT vaginal wound healing are vital to understand how estrogen and ER interactions transduce signals initiated by vaginal cell and molecular responses to injury. In this study, we found that our minimally invasive model heals regeneratively, setting a foundation to decipher how vaginal mucosa mediators regulate regenerative tissue repair, which will inevitably lead to identification of next-generation biomarkers to translate into clinical practice. Future FT vaginal wound healing studies of oophorectomized and genetically engineered, conditional cell-specific ERα and/or estrogen receptor beta (ERβ)-deficient mice (commercially available) could uncover genetic and epigenetic mechanisms of estrogen and ER signaling of vaginal wound healing. The proposed studies address the following: (a) a critical knowledge gap underpinning estrogen-driven vaginal responses to injury and (b) unmet clinical needs for the treatment of vaginal scar tissues. We conclude that our present data on postinjury variations in ERα density, dynamics, and sensitivity underscore an opportune postoperative window for the application of estrogen-targeted adjuvant therapies to optimize wound healing outcomes of pediatric, adolescent, and adult female patients.
Innovation
The mechanisms underpinning the cross talk of estrogen-dependent and inflammation–angiogenesis-driven vaginal wound repair are largely unknown. Their elucidation could lead to creation of personalized precision therapies to improve wound healing, promote regenerative healing, and reduce fibrosis by establishing vaginal wound healing kinetics using our groundbreaking murine model. We have validated histopathological methods of quantifying vaginal fibrosis and investigated the relationship of estrogen signaling in wound progression. These discoveries will shed new light onto the pathogenesis of vaginal fibrosis, which will lead to the identification and design of personalized, biomarker-guided precision therapies for vaginal wound healing.
Key Findings
Murine vaginal wound healing occurs in an accelerated manner compared with dermal wound healing, with primary epithelial closure (by day 3 postinjury) occurring long before submucosal integrity is restored.
There is an initial vaginal proinflammatory environment at 12 h postinjury that shifts to a prohealing state by 7 days postinjury.
Overall expression of ERα is directly related to wound resolution progression.
Footnotes
Acknowledgments and Funding Sources
The authors thank the TCH Office of Surgical research, especially Drs. Hector Martinez-Valdez and Monica Farenholtz, for their editorial support. Additionally, the entire CV Pathology Team at the Texas Heart Institute (especially Drs. Vela and Segura) was instrumental in this work. This work was supported, in part, through seed funding by the Department of Obstetrics and Gynecology at Baylor College of Medicine, the Clinical Scientist Training Program (CSTP) at Baylor College of Medicine, and through the Texas Children's Hospital Basic Science Surgical Research Clayton Award.
Author Disclosure and Ghostwriting
No competing financial interests exist. No ghostwriters were enlisted.
About the Authors
Supplementary Material
Supplementary Figure S1
Abbreviations and Acronyms
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.