
Abstract
Objective:
After injury, humans and other mammals heal by forming fibrotic scar tissue with diminished function, and this healing process involves the dynamic interplay between resident cells within the skin and cells recruited from the circulation. Recent studies have provided mounting evidence that external mechanical forces stimulate intracellular signaling pathways to drive fibrotic processes.
Innovation:
While most studies have focused on studying mechanotransduction in fibroblasts, recent data suggest that mechanical stimulation may also shape the behavior of immune cells, referred to as “mechano-immunomodulation.” However, the effect of mechanical strain on myeloid cell recruitment and differentiation remains poorly understood and has never been investigated at the single-cell level.
Approach:
In this study, we utilized a three-dimensional (3D) in vitro culture system that permits the precise manipulation of mechanical strain applied to cells. We cultured myeloid cells and used single-cell RNA-sequencing to interrogate the effects of strain on myeloid differentiation and transcriptional programming.
Results:
Our data indicate that myeloid cells are indeed mechanoresponsive, with mechanical stress influencing myeloid differentiation. Mechanical strain also upregulated a cascade of inflammatory chemokines, most notably from the Ccl family.
Conclusion:
Further understanding of how mechanical stress affects myeloid cells in conjunction with other cell types in the complicated, multicellular milieu of wound healing may lead to novel insights and therapies for the treatment of fibrosis.


INTRODUCTION
After injury, the excessive production and accumulation of extracellular matrix (ECM) proteins during wound healing leads to fibrosis and permanent scar tissue. 1 Several organ systems can suffer from pathologic fibrotic responses, such as pulmonary fibrosis in lungs, scar formation after myocardial infarction, and hypertrophic scar (HTS) formation after cutaneous injury. 2,3 Fibrosis and its related complications contribute significantly to morbidity and mortality, and furthering our understanding of the underlying molecular mechanisms may yield novel therapeutic strategies.
Recent studies have provided mounting evidence that the impact of external mechanical forces on intracellular signaling pathways—termed “mechanotransduction”—critically drives various cellular processes. 4 Most studies to date have focused on studying mechanotransduction in fibroblasts, the principal mesenchymal cells of the dermis that deposit and modulate collagen and other ECM proteins to form scar tissue following injury. 5,6 Fibroblasts adhere to and interact with their environment via focal adhesion kinase (FAK) complexes, and increased mechanical strain upregulates inflammatory and profibrotic transcriptional programs. 7
However, repair of damaged skin involves a dynamic interplay between not just resident cells, but also cells recruited from the circulation. 8,9 Myeloid cells, such as monocytes and macrophages, are derived from hematopoietic precursors and migrate to sites of injury where they modulate all stages of wound healing and scar formation. 10 The subpopulations of immune cells and their cellular interactions mediating skin repair and regeneration are complex and incompletely understood. While there is mounting evidence that macrophages respond to mechanical cues in certain situations, these studies have yielded conflicting results, with mechanical strain inducing both pro- and anti-inflammatory responses. 11,12
In this study, we characterized the effect of mechanical strain on myeloid cell differentiation and transcriptional signatures at a single-cell level. We utilized a three-dimensional (3D) in vitro culture system to precisely manipulate mechanical strain applied to myeloid cells and analyzed their transcriptional response and differentiation using single-cell RNA-sequencing (scRNA-seq). Our findings suggest that mechanical strain may drive myeloid cells to differentiate into macrophages and fibrocytes with proinflammatory and fibrotic transcriptional programs. Further understanding of how mechanical stress affects myeloid cells may lead to novel insights into fibrotic scarring and ultimately to new avenues in the therapeutic treatment of fibrosis.
INNOVATION
Developing therapies to promote wound healing and prevent scarring is of critical relevance to affected patients, especially those who suffer from debilitating scars and scar contractures. Clinically effective pharmacotherapies that can address this need are not currently available. Recent work has investigated the ability of disrupting mechanotransduction in fibroblasts using small-molecule inhibitors to reduce fibrosis and promote tissue regeneration. 6,7,13 Improving our understanding of how mechanotransduction affect other cell types involved in wound healing is of critical importance for the development of targeted therapeutics to mitigate fibrosis after soft tissue injury (Fig. 1).

Innovation schematic. Color images are available online.
CLINICAL PROBLEM ADDRESSED
The effect of mechanical stress on cellular differentiation and transcription affects the development of fibrosis after cutaneous injury. Understanding how to mitigate mechanotransduction across various cell types may help us develop therapies to encourage regeneration after injury.
METHODS
Cultivation of bone marrow-derived macrophages and cell seeding
We performed bone marrow isolation to culture myeloid cells according to previously published protocols. 14,15 In brief, we explanted the femurs and tibias from five C57/BL6 mice after euthanasia, specifically removing the epiphyses on both ends of the long bones. We collected the bone marrow by (1) flushing the bones with 5 mL RPMI media (ThermoFisher) and (2) crushing the epiphyses using a mortar and pestle. Red blood cells were lysed with Ammonium-Chloride-Potassium (ACK) lysing buffer (ThermoFisher). We cultured the myeloid cells in 10 mL RPMI containing 10% fetal bovine serum (FBS), 100 U/mL penicillin, 0.1 mg/mL streptomycin, 2 mM glutamine, 50 μM 2-mercaptoethanol, and 200 ng/mL murine granulocyte-macrophage colony-stimulating factor (GM-CSF) (Peprotech, Frankfurt, Germany). After 3 days, we added an additional 10 mL to the culture. To change the media, we would collect 10 mL of media, spin down the solution, and resuspend the cells in fresh media.
Myeloid-populated 3D collagen scaffold experiments
The primary myeloid cell cultures were then used to create 10 cell-populated collagen hydrogels at a final concentration of 200k cells/mL and 2 mg/mL collagen (PureCol, Advanced Biomatrix, San Diego, CA), following our previously published protocols. 16 We formulated collagen scaffolds in a cruciform shape, with sponges in the arms, and in Petri dishes with a poly-dimethylsiloxane (PDMS; Sylgard 184 Silicone Elastomer Kit; Dow Corning) coating (∼4 mm) on the bottom. We pushed pins through the arms to constrain the scaffolds in both directions for a 24 h preculture period before either No Strain or 10% equibiaxial strain conditions.
We imposed strain by manually stretching the hydrogel arms and pushing the pins back through to hold the strained position. We applied nine Titanium(IV) oxide paint dots (Sigma-Aldrich) on the surface of the hydrogel to track and quantify the imposed strains. Photographs of each marker's position were used to compute a single homogenous deformation gradient tensor
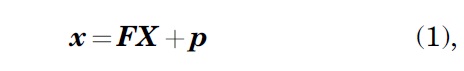
where

Single-cell barcoding, library preparation, and scRNA-seq
After 2 days of either No Strain or Strain conditions, we microdissected the collagen scaffolds and enzymatically digested them in 20 mL Collagenase, Type I (ThermoFisher) in phosphate-buffered saline (PBS) at a concentration of 5 mg/mL. 17 We subsequently mixed and rotated the cell-digest suspensions for a total of 1 h at 37°C, before using a vortex mixer (VWR) to physically disrupt the remaining collagen tissue. We then used a 100 and 70 μm Nylon cell filter (Fisher-Scientific) to filter the cells and quenched the digest in 10% FBS Dulbecco's modified Eagle medium (DMEM). Solutions were spun at 300 g for 8 min at 4°C in a centrifuge to pellet the cells and then resuspended to a final cellular concentration of 1,200 cells/μL in 0.04% bovine serum albumin (Sigma-Aldrich) in PBS, in accordance with the maximum capture concentration short of overloading, per specifications from 10 × Genomics (Pleasanton, CA). This cellular suspension was then submitted for droplet-based microfluidic scRNA-seq at the Stanford Functional Genomics Facility (SFGF) using the 10 × Chromium Single-Cell platform (Single Cell 3’ v3, 10 × Genomics). We combined a droplet of the cell suspension, reverse transcription master mix, and partitioning oil to process on the Chromium Controller. Reverse Transcription was performed at 53°C for 45 min. We amplified cDNA for 12 cycles total (Bio-Rad C1000 Touch thermocycler) with cDNA size selected using SpriSelect beads (Beckman Coulter) and a 0.6 ratio of SpriSelect reagent volume to sample volume. We used an Agilent Bioanalyzer High Sensitivity DNA chip to analyze cDNA for qualitative control purposes and then fragmented cDNA in the proprietary fragmentation enzyme blend for 5 min at 32°C, followed by end repair and A-tailing at 65°C for 30 min. cDNA were double-sided size selected using SpriSelect beats. Sequencing adaptors were ligated to the cDNA at 20°C for 15 min. cDNA was amplified using a sample-specific index oligo as primer, followed by another round of double-sided size selection using SpriSelect beads. Final libraries were analyzed on an Agilent Bioanalyzer High Sensitivity DNA chip for qualitative control purposes. cDNA libraries were sequenced on a HiSeq 4000 Illumina platform aiming for 50,000 reads per cell.
Data processing, FASTQ generation, and read mapping
Base calls were converted to reads using the Cell Ranger (10 × Genomics; version 3.1) implementation of mkfastq and then aligned against the mm9 (mouse) genome using Cell Ranger's count function with SC3Pv3 chemistry and 5,000 expected cells per sample. 18 Cell barcodes representative of quality cells were delineated from barcodes of apoptotic cells or background RNA based on a threshold of having at least 200 unique transcripts profiled, less than 10,000 total transcripts, and less than 10% of their transcriptome of mitochondrial origin.
Data normalization and cell subpopulation identification
Unique molecular identifiers (UMIs) from each cell barcode were retained for all downstream analysis. Raw UMI counts were normalized with a scale factor of 10,000 UMIs per cell and subsequently natural log transformed with a pseudocount of 1 using the R package Seurat (version 3.1.1). 19 Highly variable genes were identified, and cells were scaled by regression to the fraction of mitochondrial transcripts. Aggregated data were then evaluated using uniform manifold approximation and projection (UMAP) analysis over the first 15 principal components. 20 Automated cell-level annotations were ascribed using the SingleR toolkit (version 3.11).
Generation of characteristic subpopulation markers and enrichment analysis
Cell-type marker lists were generated with Seurat's native FindMarkers function with a log fold change threshold of 0.25 using the ROC (receiver operating characteristic) test to assign predictive power to each gene. The 100 most highly ranked genes from this analysis for each cluster were used to perform gene set enrichment analysis against pathway databases in a programmatic manner using EnrichR (version 2.1). 21
Overrepresentation analysis using Genetrail3
Using GeneTrail 3, 22 an overrepresentation analysis (ORA) was performed for each cell using the 500 most expressed protein coding genes with the gene sets Gene Ontology and WikiPathways. The p-values were adjusted using the Benjamini-Hochberg procedure, and gene sets were required to have between 2 and 1,000 genes.
CytoTRACE analysis
The CytoTRACE algorithm was used with default parameters to compare cellular differentiation states among the myeloid cells in our dataset. 23 CytoTRACE analyzes the number of uniquely expressed mRNA features per cell, as well as other factors such as distribution of mRNA content, to calculate a score assessing the differentiation and developmental potential of cells.
RNA velocity analysis using scVelo
RNA velocity analysis was performed using the dynamical model of the scVelo package. 24 Partition-based graph abstraction (PAGA) was performed using the sc.tl.paga function in scVelo.
To find genes with differentially regulated transcriptional dynamics compared to all other clusters, a Welch t-test with overestimated variance to be conservative was applied using the sc.tl.rank_velocity_genes function. Genes were ranked by their likelihood obtained from the dynamical model grouped by treatment.
Pseudotime analysis using Monocle
Monocle 3 (v0.2.1.2) was applied to construct pseudotime trajectories for individual cells along their aggregate spatial manifold. 25 The Louvain community detection algorithm was then used to group mutually similar cells and assign them to pseudotime paths and trajectories of individual cells and branches. 26 –28 We manually chose the root of origin (pseudotime = zero node) using RNA velocity analysis (location where RNA velocity = zero).
Electronic notebook
No electronic notebook was used.
RESULTS
scRNA-seq identifies multiple myeloid cell populations in a 3D culture system
To specifically interrogate how macrophages respond to changes in mechanical strain, we used a 3D culture system that permits the precise manipulation of mechanical strain (and therefore stress) applied to cells (Fig. 2a
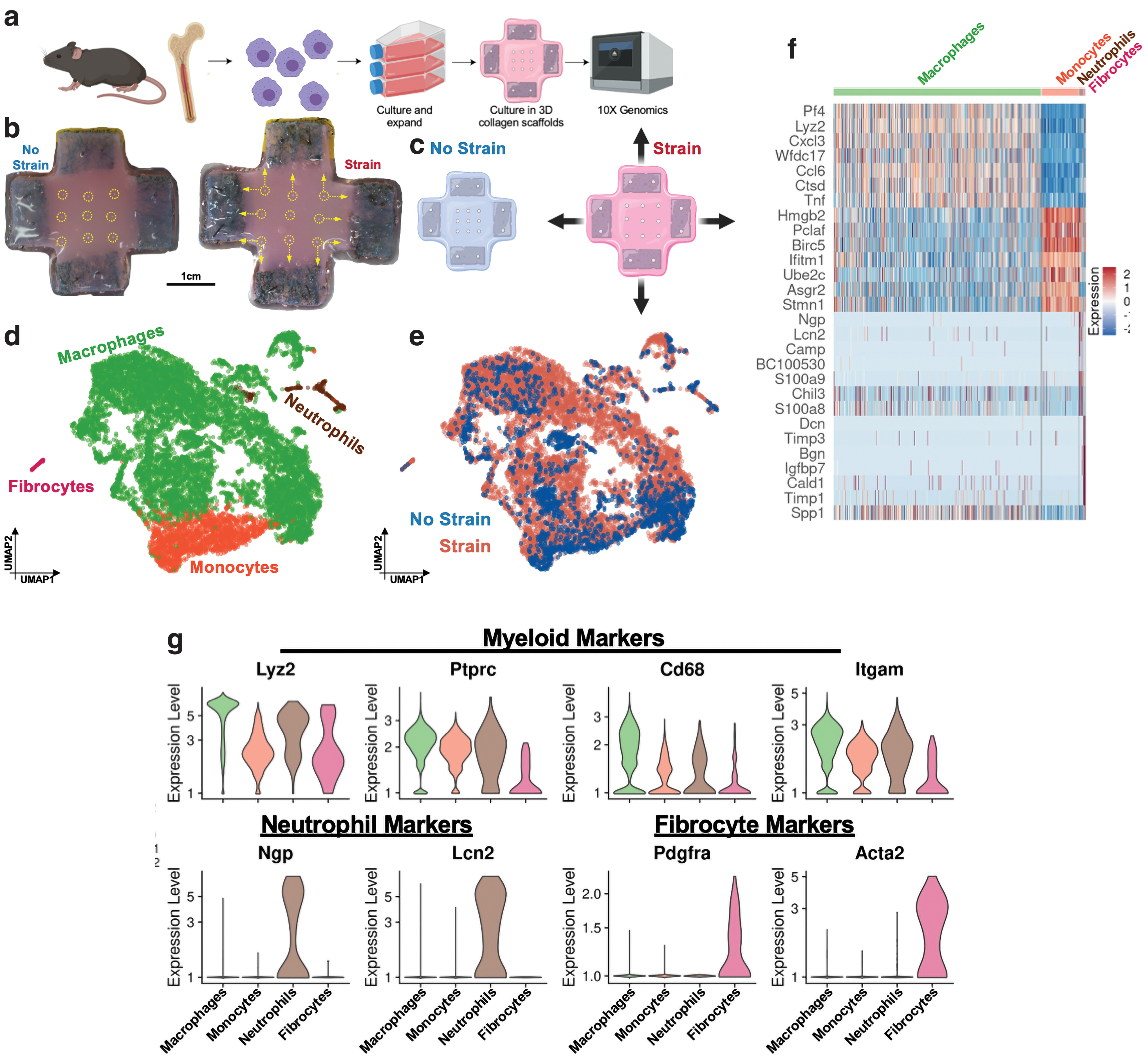
A total of 11,801 cells were captured, with 1,757 monocytes, 9,796 macrophages, 180 neutrophilic granulocytes, and 68 “fibrocytes” identified using semisupervised cell type annotations through the SingleR package and evaluation of cell type-specific marker genes (Fig. 2e). All cells expressed common hematopoietic markers Lyz2 and Ptprc (encodes for CD45) (Fig. 2h). Neutrophils expressed specific genes such as neutrophil granule protein (Ngp) and neutrophil gelatinase-associated lipocalin (Lcn2) (Fig. 2g, h). “Fibrocytes” were initially categorized as fibroblasts due to expression of Pdgfra and Acta2 (Figs. 2g, h and 3a); however, the coexpression of the common hematopoietic markers Ptprc (CD45) and Lyz2 indicated that these cells were indeed derived from the hematopoietic lineage, which is in line with our myeloid cell culture protocol. Therefore, we classified these cells as fibrocytes, according to previous studies that have highlighted the role of this rare population of hematopoietic collagen producing cells in connective tissue remodeling and fibrosis. 30,31 Fibrocytes also highly expressed ECM markers Col1a1 and Dcn (encodes for Decorin), as well as Timp1/3, which regulates collagen deposition and degradation.
Mechanotransduction promotes monocyte differentiation into proinflammatory macrophages
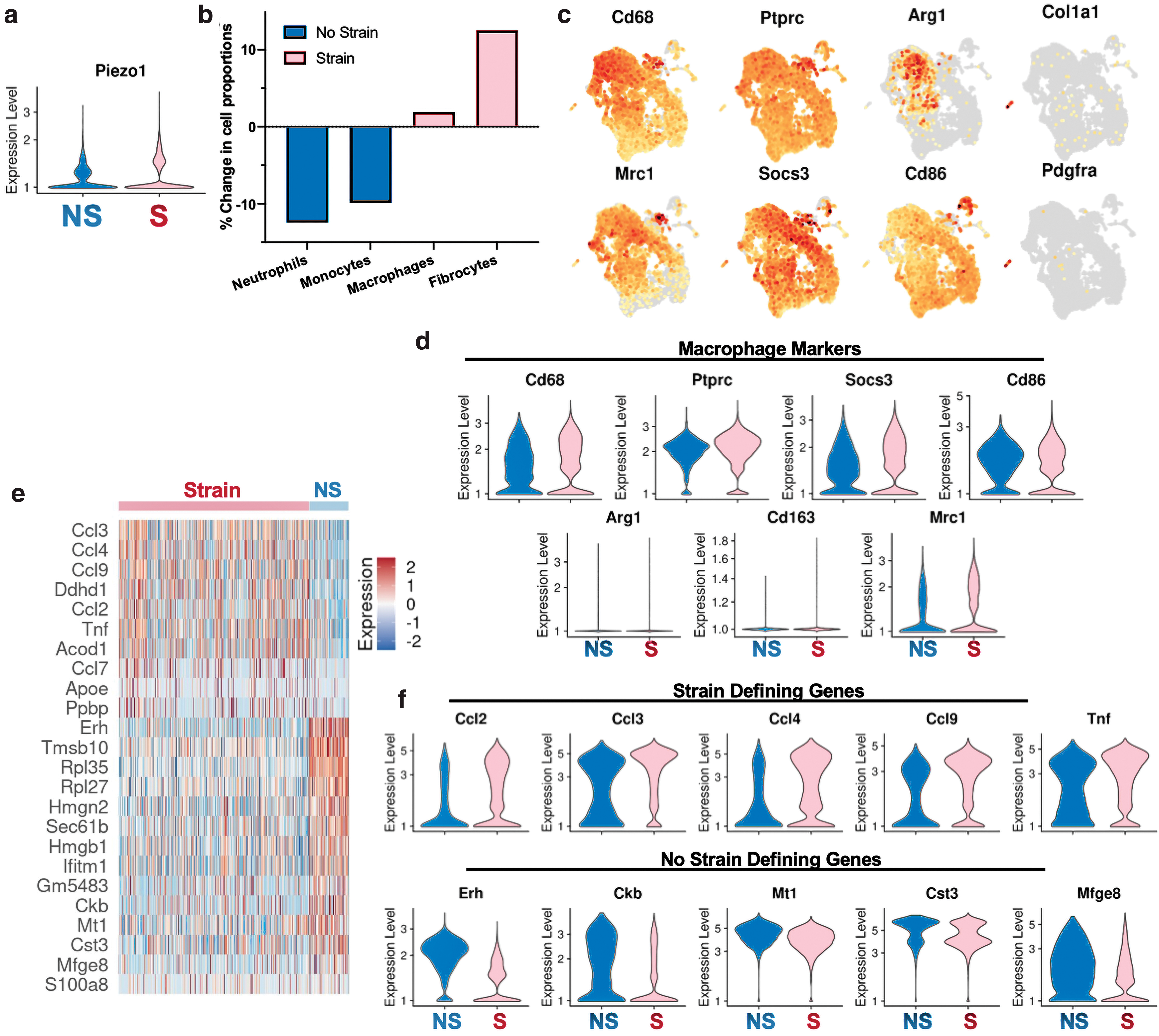
We captured 2,005 “No Strain” (NS) cells and 9,796 “Strained” (S) cells and observed that mechanically strained cells demonstrated a clear shift in transcriptional programs (Fig. 2f). First, mechanical strain activated Piezo1 expression, which has been recently implicated in regulating macrophage mechanotransduction 32 (Fig. 3a). The NS cells had relatively more neutrophils and monocytes, while strain activated and increased the relative number of macrophages and fibrocytes (Fig. 3b). When looking at common myeloid markers (Cd68, Ptprc) and classical M1 and M2 macrophage markers (Socs3, Cd86, Arg1, Cd163, and Mrc1), we observed that expression of these markers was relatively evenly distributed in both NS and S cells (Fig. 3c, d). Thus, traditional activation of M1 and M2 markers did not correlate with mechanical strain in this system.
We observed that strain activated a family of chemokine (C-C motif) ligand (Ccl) genes, all representing increased inflammatory responses (Fig. 3e, f). Ccl3, Ccl4, and Ccl9 are chemoattractants secreted by macrophages and myeloid cells and involved in recruiting other inflammatory and immune cells, such as monocytes or NK cells. 33,34 These genes encode for macrophage inflammatory proteins (MIP) 1-alpha, 1-beta, and 1-gamma, respectively, which have been specifically linked to upregulating proinflammatory behaviors. Mechanotransduction also upregulated Ccl2, encoding for monocyte chemoattractant protein (MCP-1), which recruits monocytes, memory T cells, and dendritic cells to the site of inflammation. Strain also upregulated Tnf, which has been extensively studied as a secreted macrophage cytokine that plays a significant role in the development of fibrosis. 35
Mechanotransduction promotes transcriptional diversity among myeloid cells
We identified nine transcriptionally distinct clusters (c0 to c8) in our dataset (Fig. 4a). Mechanical strain strongly altered transcriptomic profiles and shifted macrophage transcriptional programs primarily into c0, c5, c6, and c8 (Fig. 4b). Cells in c0 were defined by increased expression of Thbs1, a gene linked to macrophage activation and subsequently increased inflammation (Fig. 4c, d). The family of Thbs genes encode for thrombospondin proteins, which facilitate cell binding to ECM, associate with increased fibrosis, and have antiangiogenic functions. 36 c5 cells increased expression of an array of Ccl and Ccr genes, including Ccl5, Ccr7, and Ccl22, while c6 cells increased expression of Cxcl10, a chemokine that mediates macrophage-associated inflammation. c8 cells were identified as fibrocytes. We then used Genetrail3, a computational pipeline for ORA of specific gene sets on a single-cell level, to further investigate differential regulation of cellular signaling pathways. 22 We found that strained macrophages exhibited induction of gene sets for cellular response to tumor necrosis factor (GO-BP), positive regulation of inflammatory response (GO-BP), and lung fibrosis (Wiki Pathways; WP) (Fig. 4e). Together, these data demonstrated that mechanically strained macrophages upregulated a cascade of inflammatory Ccl genes, resulting in an increased expression of various inflammatory and fibrotic pathways. Interestingly, traditional inflammatory M1 markers such as Cd86 did not correlate with mechanical strain.
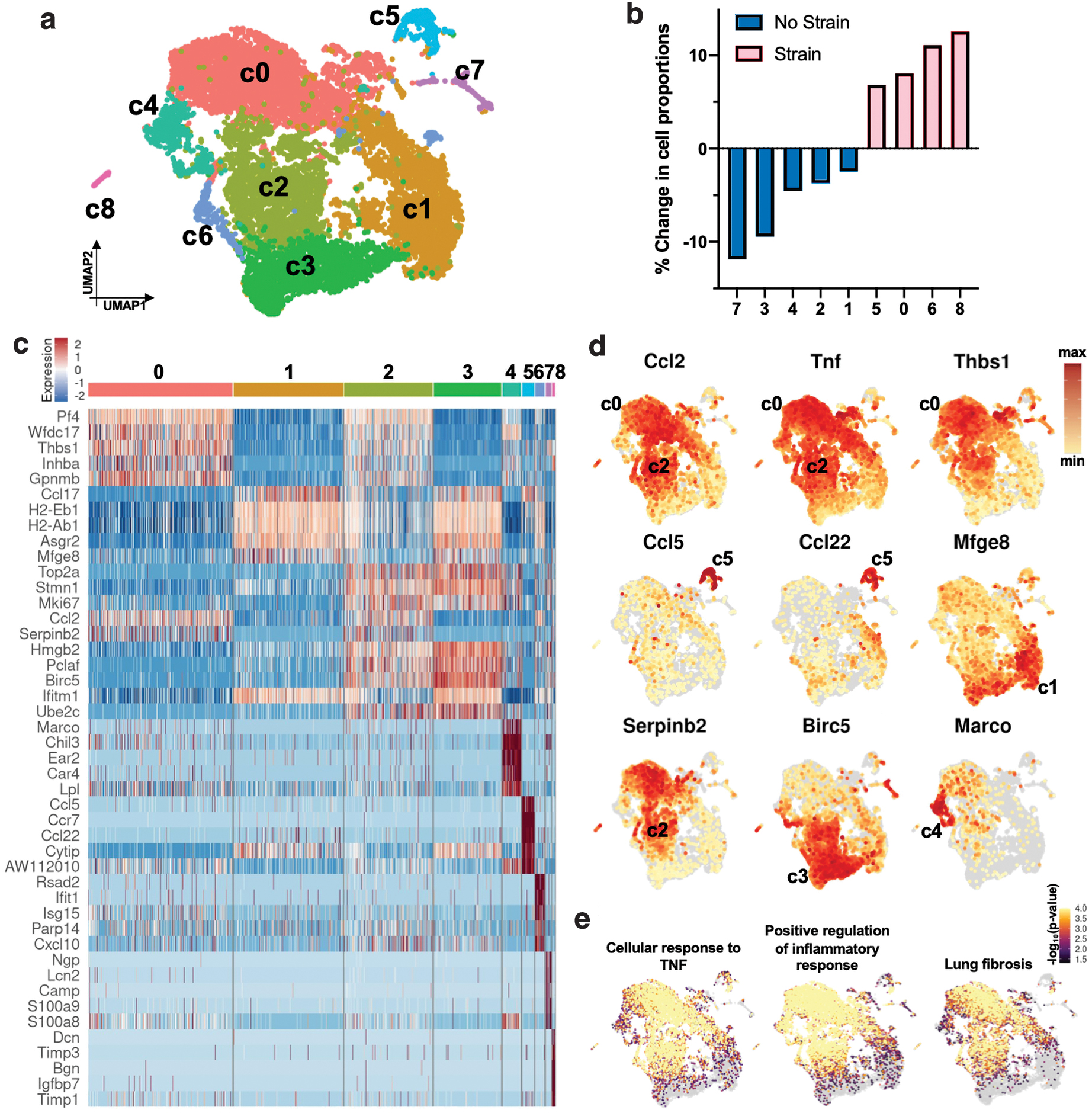
By contrast, unstrained cells aggregated primarily in clusters c1, c2, c3, c4, and c7 (Fig. 4b). c1 cells demonstrated increased expression of both Ccl17, another inflammatory marker of the Ccl family, as well as Mfge8 (Fig. 4c, d) (encoding for milk fat globule-EGF factor 8 protein or lactadherin), which decreases fibrosis by reprogramming macrophages to anti-inflammatory phenotypes and promoting wound closure through fibroblast migration. 37 c2 cells demonstrated increased expression of Serpinb2, which importantly reduces inflammation and fibrosis 38 ; c3 upregulated Birc5, which promotes macrophage function by mediating apoptosis 39 ; and c4 increased expression of Marco (encoding for “macrophage receptor with collagenous structure”), which binds to pathogens and promotes macrophages to scavenge and phagocytose pathogens. 40 c7 cells were identified as neutrophils.
Mechanical signaling encourages a unifying trajectory of monocyte differentiation
Traditional differential expression analysis only provides a snapshot of mRNA expression. We therefore used RNA velocity analysis using the scVelo package to explore the comparative abundance of spliced and unspliced pre-mRNA transcripts in these myeloid cells. 24 By analyzing mRNA splicing states and transcriptional dynamics in an unbiased manner, 24,41 we identified a unifying trajectory of monocyte differentiation, moving from NS to Strained cells (Fig. 5a, b). The monocytes (c3) were identified as the origin of this velocity stream. Correlating with our velocity analysis, CytoTRACE analysis also identified the monocytes as the root cells of differentiation, with the highest differentiation potential (Fig. 5b). 23
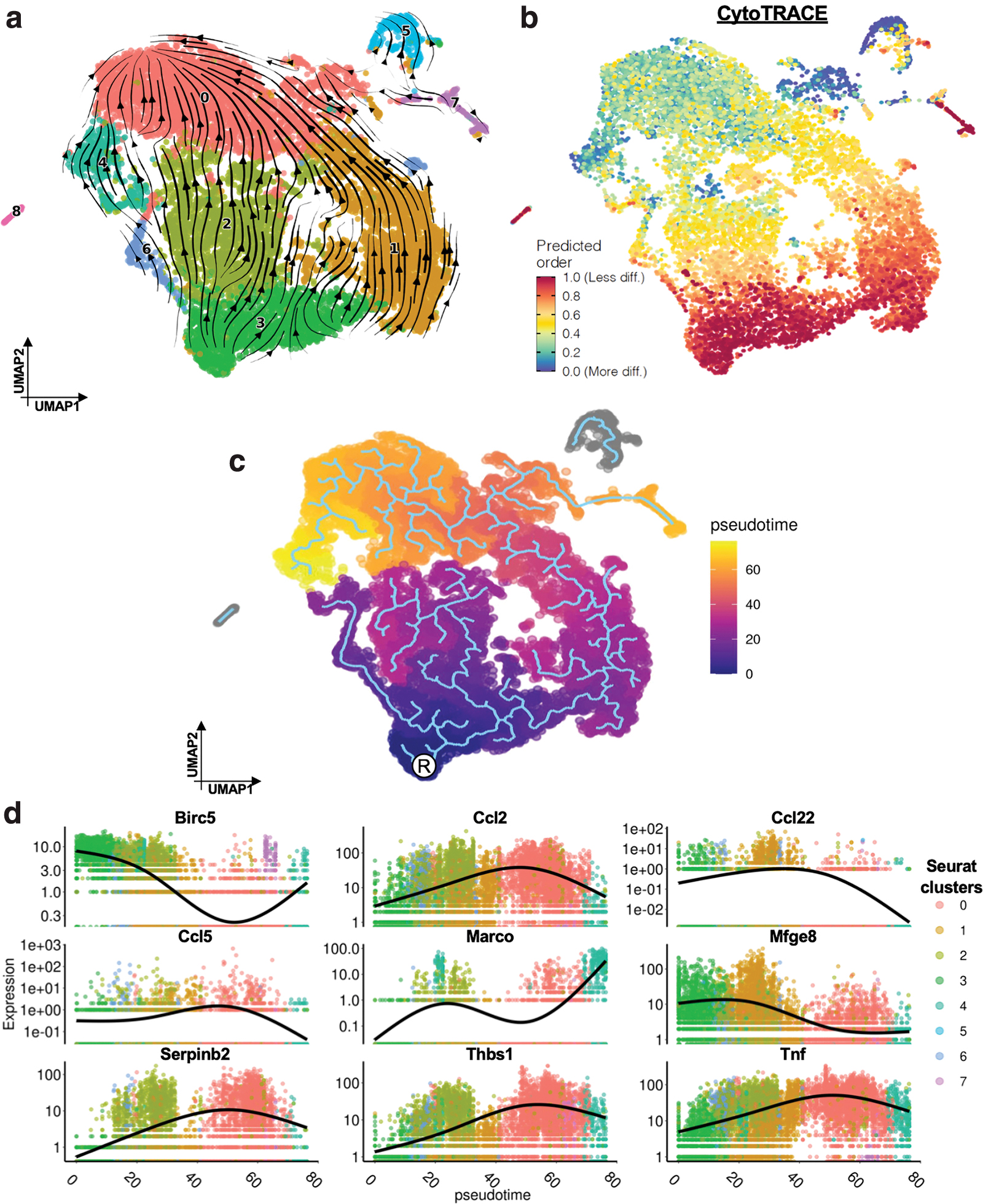
To further understand the transcriptional shifts observed in our single-cell data, we used the Monocle3 tool to calculate and assign a “pseudotime” value for each of our cells. 42 In this analysis, pseudotime is used as a surrogate measure to determine the transcriptional similarities or differences between cells. Cells that are more transcriptionally similar to each other have more similar pseudotime values, and transcriptional trajectories can be inferred to predict how different subpopulations of cells may differentiate into each other. We found significant pseudotemporal divergence between mechanically strained macrophages and NS cells (Fig. 5c). We identified an inflammatory trajectory from the root cells (root, R) to the mechanically activated cell clusters and observed that the differentiation of macrophages as well as inflammatory gene expression (Ccl2, Tnf, etc.) correlated with pseudotime (Fig. 5d). The trajectory progressed from c3 (monocyte cluster), through c1 and c2 (regenerative myeloid cells), to c0 (strained, activated cells).
DISCUSSION
Mechanotransduction underlies a number of critical physiological processes in the body, including proprioception, touch, balance, and hearing. 43 Within the context of wound healing, recent studies have highlighted that upregulated mechanotransduction signaling may affect the cellular ecology and transcriptional dynamics and alter healing capacity. 44 While we know that aberrant mechanotransduction activates fibroblasts to excessively produce collagen and form scar, 4 the role of mechanotransduction on the inflammatory response is still incompletely understood. 45 We have previously shown that increasing mechanical strain within a murine model of HTS formation upregulates specific fibrotic and inflammatory signals such as FAK, ERK (extracellular signal-regulated kinase), and MCP-1. 7 However, these previous studies were limited by the available technologies at the time and were unable to fully examine the plethora of cellular signaling pathways, which are altered in response to mechanical stress. Recent advances in single-cell transcriptomics have increased our ability to comprehensively investigate these cellular responses for the first time. 17
Myeloid cells, which are critically involved in acute inflammation after soft tissue injury, as well as chronic fibrosis, have recently been identified as being mechanically sensitive. 10 Solis et al. demonstrated in 2019 that myeloid cells sense mechanical force through the pressure-sensitive channel PIEZO1. In macrophages exposed to pressure cycles, PIEZO1 signaling drove a proinflammatory expression profile. 46 In this study, we demonstrated that increased mechanical stretch also upregulated PIEZO1 expression. We have previously shown that fibroblasts integrate strains from both stretch and compression and align themselves in a direction that minimizes their strain field. 16 It seems likely that myeloid cells would also integrate stretch and compression, potentially through the PIEZO1 signal pathway. Future work will need to be done to interrogate the combinatorial effects of both stretch and compression.
While other studies have also investigated the effects of mechanical strain on myeloid cells, they have typically used stiff, silicone membranes to cyclically load macrophages to mimic other organs such as the vasculature or lungs. 12 In the context of wound healing, we used a 3D collagen matrix and static stretch profile that more physiologically mimics the wound environment and observed increased Piezo1 expression. 46,47 Normal human skin typically experiences a maximum strain of 20% at maximum extension. 48 Since it seems likely that a healing wound would not be subjected to maximal strains (i.e., because of pain), we used a strain profile of 10%. Previous research has found that static strain stiffens the collagen matrix, which then exhibits stress relaxation over time. 49 Thus, our investigations demonstrate a stabilized response from the macrophages, integrating both the initial static stretch along with subsequent matrix relaxation over the course of 48 h.
Surprisingly, we found that strain did not specifically activate macrophages to differentially upregulate genes related to classically defined M1 or M2 subsets. We utilized a standard culture protocol to differentiate myeloid cells from bone marrow precursors in the presence of GM-CSF. Further studies using different myeloid cell culture protocols are needed to fully understand the impact of mechanical force on macrophage polarization.
Instead, our results show that increasing mechanical strain correlated with a cascade of inflammatory Ccl and Tnf genes, demonstrating how these already inflammatory “M1”-like macrophages further increased their proinflammatory signal expression. Tumor necrosis factor-α (TNF-α) has been identified as a potential therapeutic target to combat fibrotic diseases. 35 Zhang et al. found that TNF-α released in macrophage supernatants positively correlated with increased inflammatory recruitment and ECM deposition. 35 We have previously found that increased mechanical strain in a murine model of HTS upregulated MCP-1 (protein form of Ccl2) expression in fibroblasts, further triggering inflammatory pathways to potentiate a fibroproliferative state.
Our collective findings indicate that increased mechanotransduction also upregulates expression of Ccl2 and other Ccl genes in myeloid cells, further contributing to this “fibroproliferation.” These findings indicate that mechanical activation increases inflammation in myeloid cells, agreeing with our previous findings in fibroblasts. 7 Within the physiologic tissue environment, other factors could also influence macrophage inflammatory responses, such as myofibroblast contraction of the surrounding tissue shifting macrophage polarization 50,51 or changes in the cytokine environment reversing inflammatory responses over time. 52,53 Future studies will need to investigate the ability of inflammatory responses to be reversed or how coculture of fibroblasts and macrophages could affect responses to mechanical activation.
Within our dataset, we also identified heterogeneous myeloid clusters. In general, clusters with low mechanotransduction (No Strain group: c1, c2, c3, c4, and c7) each expressed a subset of anti-inflammatory, anti-fibrotic, or angiogenic gene sets (e.g., Mfge8, Serpinb2, Marco). By contrast, strained clusters (c0, c5, c6, c8) each expressed a subset of inflammatory genes such as Thbs1 or additional Ccl and Ccr genes. We used various scRNA-seq tools (RNA velocity, Monocle3, and CytoTRACE) and found that they all independently identified the same origin of differentiation within the monocyte cell subpopulation and majority of the NS cells. From this root, we observed that monocytes (NS cells) became activated by mechanical stress to primarily differentiate into heterogeneous macrophage clusters. While these subsets of myeloid cell types have been confirmed by other groups, 22,36 –40 our dataset holistically captures these heterogeneous myeloid populations together to provide insight on how they each contribute to the development of fibrosis.
We also observed a small population of fibrocytes, which have been previously identified in small quantities in vivo. Various studies have investigated the ability of myeloid lineage cells to differentiate into fibrocytes that may contribute to excessive fibrotic outcomes. 30,31 These cells have been found to initially exhibit both inflammatory properties of macrophages as well as tissue remodeling properties of fibroblasts, initially migrating to sites of injury to secrete chemokines before transitioning to lay down ECM, proliferate, and contribute to tissue repair. 54,55 Studies have shown that these cells may be specifically responsive to CCL genes and the CXCL12-CXCR4 axis and that targeting these pathways to reduce fibrocyte numbers effectively reduces pulmonary fibrosis. 56,57
In this study, we observed that mechanical strain caused a relative increase in fibrocyte numbers, indicating that mechanical activation may play a role in triggering fibrocyte differentiation from myeloid precursors. To our knowledge, this relationship has not been previously explored, and future studies should further interrogate the effect of mechanical loading on fibrocyte differentiation. However, there are conflicting reports as to the exact origin of these cells and their relationship to mesenchymal stromal cells. Some studies claim that fibrocytes originate from mesenchymal progenitors, 58 while others claim that CD14+ monocyte progenitor cells may differentiate to fibrocytes. 57
Overall, our study provides a comprehensive characterization of the effects of increased mechanical forces on myeloid cell transcriptional fates. We demonstrate that monocytes are activated by mechanical forces and differentiate toward proinflammatory macrophage and fibrocyte subpopulations, characterized by an induction of chemokine signaling and profibrotic gene expression. Our work lays the groundwork for future studies to further interrogate this mechanically driven myeloid differentiation process in animal models of wound healing and scar formation. A better understanding of the underlying mechanisms driving inflammation and fibrosis will improve future therapies to promote healing and prevent fibrosis.
KEY FINDINGS
Myeloid cells are mechanoresponsive.
Mechanically strained monocytes differentiate into macrophages with increased inflammatory transcription, primarily from Tnf and Ccl family genes.
Mechanotransduction increases myeloid differentiation into fibrocyte populations.
DATA AVAILABILITY
The scRNA-seq data discussed in this publication have been deposited in NCBI's Gene Expression Omnibus 59 and are accessible through GEO Series accession number GSE175460.
Footnotes
AUTHORS' CONTRIBUTIONS
K.C. and G.C.G. designed the study. K.C., D.H., D.S., C.A.B., M.G., H.C.K., and A.A.T. performed the animal and cell culture experiments. K.C., D.H., C.A.B., and M.G. performed the single-cell RNA sequencing experiments. K.C., D.H., and M.J. performed the bioinformatics analyses. K.C., D.H., D.S., M.J., and G.C.G. wrote the article. J.P., D.C.W., and M.T.L. helped revise and edit the article.
ACKNOWLEDGMENTS AND FUNDING SOURCES
We thank Theresa Carlomagno for administrative support. Single-cell sequencing was supported by the Stanford Functional Genomics Facility (SFGF) with funds from the NIH (S10OD018220 and 1S10OD021763). This work was supported by the Center for Dental, Oral, & Craniofacial Tissue & Organ Regeneration Interdisciplinary Translational Project Awards supported by the National Institute of Dental & Craniofacial Research (U24 DE026914).
AUTHOR DISCLOSURE AND GHOSTWRITING
No competing financial interests exist. None declared.
ABOUT THE AUTHORS