
Abstract
Significance:
The aberrant inflammation during wound healing results in pathological scarring, such as hypertrophic scars and keloids. This adversely affects the quality of life of patients due to the disfiguring appearance as well as the symptoms of itch and pain. This review summarizes the up-to-date knowledge of the immunopathogenesis and treatment options for pathological scars.
Recent Advances:
With the advent of new technologies, combined with in vitro and in vivo wound models, several inflammatory cells have been shown to have both direct and indirect effects on both wound healing and pathological scarring.
Critical Issues:
Expansion of pro-fibrotic immune cells such as M2 macrophages, dendritic cells, mast cells, and Th2 cells leads to fibroblast transition to myofibroblasts via transforming growth factor-β1 signaling pathway. Appropriate management of such inflammatory responses during wound healing remains a critical issue during clinical practice.
Future Directions:
Regulating inflammation response during wound healing may be a potential therapeutic option for avoiding or reducing pathological scars.

SCOPE AND SIGNIFICANCE
Inflammation triggered by both innate and adaptive immune cells plays a key role in the early phase of wound healing. The dysregulation of inflammation can promote prominent pathological scarring by activating fibroblasts and increasing extracellular matrix (ECM) deposition. Our goal is to summarize the biological functions of immune cells, including macrophages, dendritic cells, neutrophils, mast cells, and T cells, involved in wound healing and pathological scarring. We also summarize the molecular mechanisms involved in cell-cell interactions between immune cells and stromal cells, and we highlight the contemporary therapeutic approaches.
TRANSLATIONAL RELEVANCE
Pathological scars substantially affect the patient's quality of life. Understanding the molecular mechanisms involved in the inflammation during wound healing and scarring can help us to identify the potential preventive and therapeutic targets.
CLINICAL RELEVANCE
More than 100 million patients have developed scars after trauma or surgery, and roughly 15% of those patients with scars are prone to hypertrophic scar or keloid. 1 The resulting disfigurement from scarring not only leads to cosmetic nuisances but also places significant physical and psychological burdens on patients. 2 However, there are no effective preventions or mono-therapeutic treatments for keloid, highlighting the great difficulty in treating this condition. 3
Inflammation responses are regarded as a key factor in the formation of hypertrophic scars and keloids. 4 –7 Combining immunomodulators with other therapies seems to be a promising strategy for the treatment or prevention of pathological scars.
BACKGROUND
Wound healing
Cutaneous wound healing is a repair process after skin injury and has three overlapping but distinct stages, namely: the hemostasis/inflammatory phase, the proliferation phase, and the remodeling phase (Fig. 1). 8 During the hemostasis/inflammatory phase, which occurs immediately after tissue damage, fibrin clots form in conjunction with platelets to control bleeding and prevent the tissue from being exposed to external spaces.
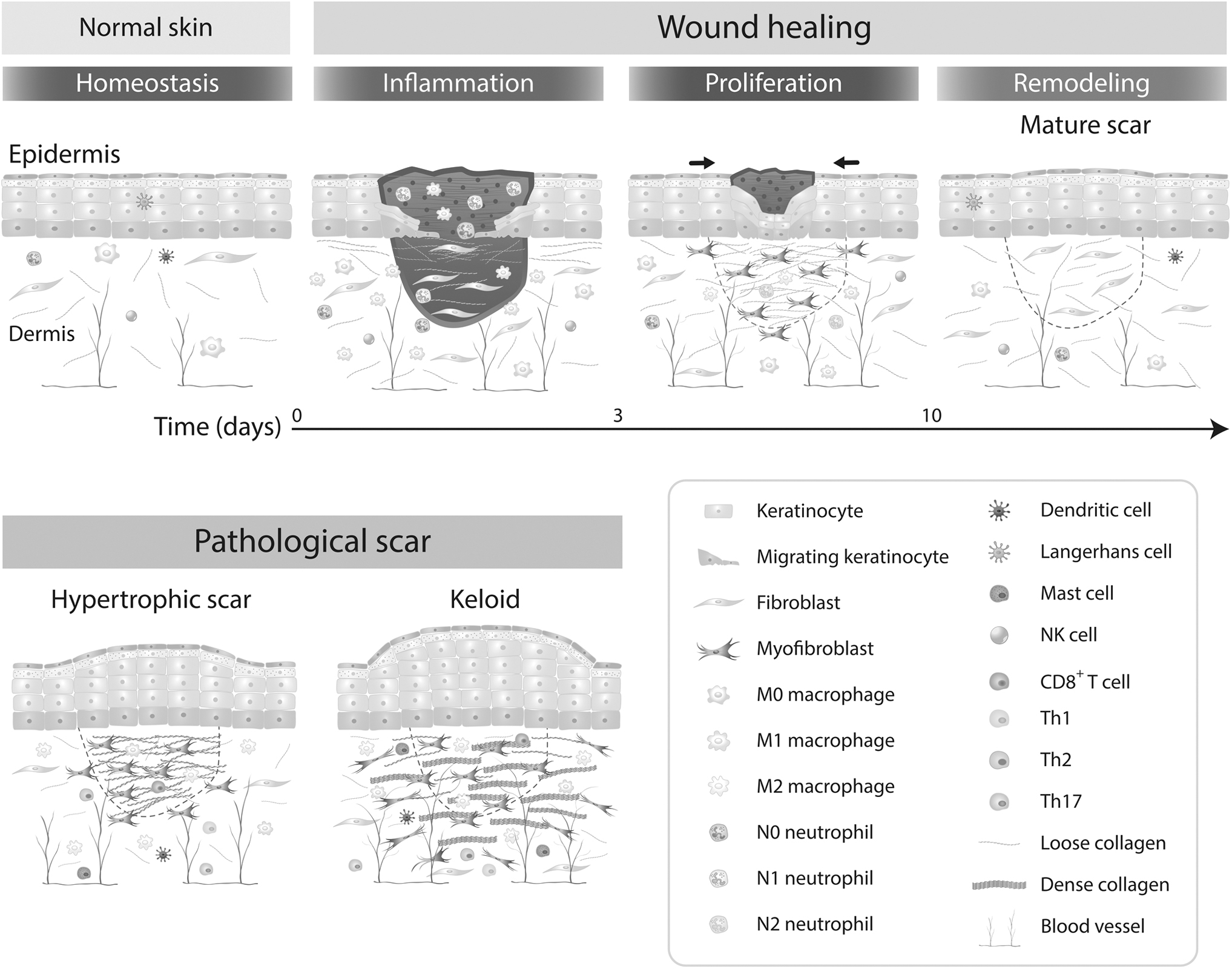
Wound healing and pathological scars. The complex process of wound healing can be grouped into three phages—“inflammation,” “proliferation phase,” and “remodeling phase.” In pathological conditions, myofibroblasts persist and continue to produce excessive extracellular matrix. Keloids are characterized clinically by fibrotic expansion beyond the boundary of the original wound, whereas hypertrophic scars are elevated scars that remain within the boundaries of the wound.
Histamine, bradykinin, and leukotrienes derived from resident mast cells increase vascular permeability and recruit immune cells. In response to stress signals, neutrophils and monocytes transmigrate from the vasculature to the wounded tissue within minutes to perform phagocytosis, antigen presentation, and cytokine production. 9 Pathogen colonization activates the innate immune cells, including neutrophils, monocytes, natural killer cells, gamma-delta T cells, and invariant natural killer T cells. 10
Next, in the proliferation phase, macrophage polarization toward the alternative (M2) phenotype facilitates fibroproliferation, re-epithelialization, and neovascularization in wound healing. 11 Fibroblasts that proliferate and migrate into the wound bed play a major role in granulation tissue formation. Transforming growth factor-β (TGF-β) and platelet-derived growth factor (PDGF)-BB secreted by macrophages stimulate the transformation of fibroblasts into myofibroblasts, causing wound contraction.
These myofibroblasts are also responsible for the synthesis of the various components of the ECM, which includes collagen, fibronectin, elastin, and glycosaminoglycans. 12 Epidermal growth factor family members trigger local keratinocyte migration to the upper layers of the granulation tissue, to aid in wound closure. Meanwhile, endothelial cells proliferate and migrate in response to vascular endothelial growth factor (VEGF) to promote angiogenesis. Newly synthesized capillary sprouts are formed to support local cells with nutrients and oxygen. 13
Lastly, in the tissue remodeling phase, specific cell populations in the granulation tissue undergo the apoptosis or quiescence and the secretion of enzymes to remodel the matrix. 14
In human wound healing, the ratio of collagen type I (COL1A) and collagen type III (COL3A1) plays a critical role in scar formation. The ratio of COL3A1:COL1A1 decreases from the early to late phase of wound healing. COL1A1, therefore, serves a dual function of maintaining stable tension in the wound area but may also promote scarring. 15 Myofibroblasts from hypertrophic scars and keloids have an increased capacity to resist apoptosis in comparison to normal wound myofibroblasts, which may imply an abnormal tissue remodeling phase in pathological scars.
Pathological scarring
Pathological scars contain hypertrophic scars and keloids that feature excessive ECM deposition and fibroblast proliferation (Fig. 1). 16 Keloids grow beyond the margins of the original wound and do not spontaneously regress. Histopathological observation of hypertrophic scars shows vertically oriented blood vessels, a horizontally fibrous bundle, and higher α-smooth muscle actin (α-SMA) expression. Keloids are characterized by fibrous fascicles, tongue-like protruding edge underneath the papillary dermis, and thick hyalinized eosinophilic collagen fibers (keloidal collagen) in the dermis. 17
The association between the immune system and keloid has been previously illustrated. 18 Innate and adaptive immune cells play a role in myofibroblast transdifferentiation indirectly by secreting cytokines or growth factors, which contribute to excessive ECM production. 4 Several pieces of evidence have demonstrated that macrophages can secrete many types of collagens, which may further modulate the cell-cell and cell-matrix interaction. 19
These findings propose a direct and indirect role for immune cells in ECM synthesis during pathological scarring. Some studies suggest that the likely origin of myofibroblasts is from circulating bone marrow-derived cells. 20 Importantly, the population of bone marrow-derived cells such as fibrocytes, pericytes, and macrophages are increased in pathological scarring. 21 Fibroblasts from the reticular layer have been implicated in the pathogenesis of fibrotic skin diseases. 22
In addition, chronic inflammation is significant in the reticular dermis during scar formation. 5 These results implicate that reticular fibroblasts may interact with immune cells and trigger the progression of fibrosis.
Inflammation
Inflammation, orchestrated by inflammatory cells, mediators, and vasodilation, is a complex biological response to various factors, such as pathogens, irritants, and injured cells. 23 The complex human immune response can be roughly separated into two categories: innate immunity and adaptive immunity (Fig. 2). 24 The innate immune system is the first-line defense against pathogen penetration into the body surface.
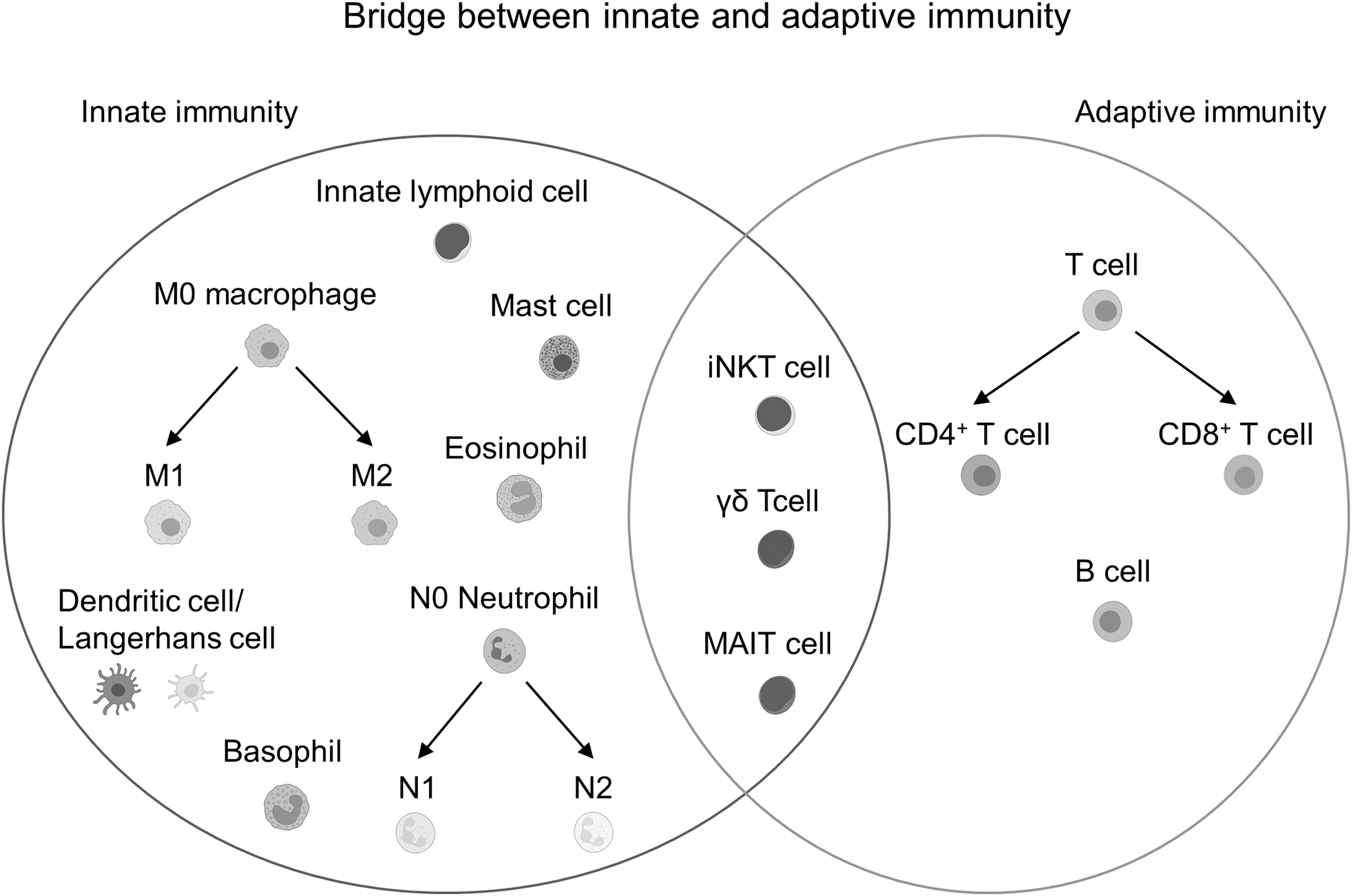
Immune cells in the innate and adaptive responses. Macrophages, dendritic cells/Langerhans cells, neutrophils, mast cells, and innate lymphoid cells belong to innate immunity, whereas T cells and B cells are attributed to adaptive immunity. Innate-like T cells, including iNKT cells, γδ T cells, and MAIT cells, play a role in both the innate and adaptive immune systems. Cell template is created by
The main functions of innate immunity include the recruitment of immune cells, the release of cytokines, complement cascade, the removal of pathogens, and the presentation of antigens to the adaptive immune system. However, the adaptive immune system features immunological memory that enables it to accurately recognize targets and immediately respond when encountering the same pathogenic stimuli again. T cells and B cells are the main populations of the adaptive immune system.
Keloid initiation has been shown to be associated with genetic predisposition, skin tension, and inflammation. 25 In addition, scarless healing and reduced inflammatory cell infiltration are hallmarks of human fetal wound healing, suggesting that the inflammation response of immune cells in adults plays a critical role in scar formation. 26 In this review, major cellular and molecular mechanisms by which inflammatory cells regulate wound healing and pathological scars are discussed.
The detailed involvement of inflammatory cells regarding innate and adaptive immunity in wound healing and pathological scar formation is described individually. Finally, we summarize novel insights into potential therapeutic interventions that could regulate specific inflammatory mediators responsible for pathological scarring.
DISCUSSION
Macrophages
Wound healing
Macrophages play a role in the homeostasis and repair of the skin by interacting with stromal cells and activating adaptive immune responses. Macrophages can be further sorted into a classical phenotype (M1 macrophages) and an alternative phenotype (M2 macrophages). CCR2+/Ly6C+ M1 macrophages, induced by lipopolysaccharide/interferon-γ, exert their inflammatory function by producing pro-inflammatory cytokines such as interleukin (IL)-1β, IL-6, and tumor necrosis factor (TNF)-α. 27
CD163+ (scavenger receptor)/CD206+ (mannose receptor) M2 macrophages, stimulated by IL-4/IL-13 or apoptotic neutrophil, promote proliferation of keratinocytes and fibroblasts by secreting PDGF, TGF-β, insulin-like growth factor (IGF)-1, and VEGF. 28
Monocyte-derived resident macrophages found in the dermis contribute to the wound-healing process. 29 The depletion of macrophages during different phases of wound healing results in different sequences of repair events. 30 After wounding, macrophages clear microbes and cell debris by opsonization and phagocytosis, which are activated by alternative complement pathways and phagolysosome formation respectively. 31
Chemokine receptors CX3CR1 and CCR2, for CX3CL1 and CCL2 respectively, are essential for the recruitment of circulating monocytes to the wound site. Blocking the CX3CR1-CXCL1 or CCR2-CCL2 axis delays the wound-healing process and results in the reduction of macrophage number, collagen deposition, and neovascularization, highlighting the important role of macrophages in the inflammatory phase of wound healing. 32,33
During the proliferative phase of wound healing, the phagocytosis of apoptotic neutrophils and cell debris promotes an M2 macrophage expansion which secretes TGF-β1 and VEGF-A, which further contributes to fibroblast differentiation and angiogenesis. 34 At the remodeling stage, M2 macrophages engulf residual ECM and debris through mannose receptor recognition of collagen and matrix metalloproteinase (MMP)-10-mediated collagen degradation. 35,36
Deficiency of Krüppel-like factor 4 (KLF4), a zinc-finger transcription factor for autophagy, in myeloid cells decreases M2 polarization and wound healing. 37 Extracellular ATP administration accelerates wound healing and M2 macrophage recruitment, suggesting that a metabolic change regulates macrophage programming. 38 Therefore, modulators of M2 macrophage expansion can greatly enhance the wound-healing process. 39
Pathological scarring
Wound bed macrophages expressing CD301b, a prototypical M2 marker that selectively activates proliferation of adipocyte precursors, 40 are considered to be the origin of myofibroblasts in large wounds. 41 This observation could imply that M2 macrophages may contribute to the increased number of myofibroblasts. As expected, the increased population of M2 macrophages relative to M1 is usually present in hypertrophic scars and keloids. 42,43
An increase in COL1A1+ (collagen type I)/CD204+ (macrophage scavenger receptor 1) M2 macrophages is also observed in peripheral blood mononuclear cells (PBMCs) from the patients with major thermal injuries. 44 Interestingly, IL-17 may induce the infiltration of macrophages to aggravate fibrosis through monocyte chemotactic protein (MCP)-dependent mechanism. 45 Meanwhile, human-specific gene CHRFAM7A gene expression is significantly lower in hypertrophic scars, and CHRFAM7A attenuates M2 polarization via the Notch pathway, which eventually mitigates scar formation. 46
Nevertheless, M2 macrophages are also found to have an anti-fibrotic effect via IL-6- and MMP-9-dependent mechanisms in in vitro and in vivo studies. 47,48 Fingolimod (FTY720), a sphingosine-1-phosphate receptor antagonist, improves the mechanical force-induced abnormal scars in mice by increasing M2 polarization during the proliferative stage. 49 Therefore, various regulated pathways and molecules could lead to the different effects of M2 macrophage on pathological scars.
Taken together, therapeutic strategies that utilize macrophage polarization might be applied to prevent or treat pathological scarring although further studies are necessary to clarify the molecular mechanisms of such treatments.
Dendritic cells and langerhans cells
Wound healing
Dendritic cells with CD1a and CD207 markers are a type of antigen-presenting cells that are responsible for activating adaptive immune responses. Langerhans cells, a subset of immature dendritic cells, are distinguished from other dendritic cells by the presence of cytoplasmic racket-shaped Birbeck granules and the residing in the basal and supra-basal layer of the epidermis. 50 Langerhans cells function as part of the first line of defense by aiding the degradation of intruding pathogens. 51
During steady states without epidermal injury or inflammation, Langerhans cells are characteristically maintained by proliferation in situ, 52 whereas myeloid dendritic cells are replenished by bone marrow-derived precursors. The migration of Langerhans cells from the skin to the draining lymph nodes occurs even during unwounded steady states and is enhanced during inflammation. 50
Studies on Langerhans cells have focused primarily on their migration and repopulation in the skin. When an acute injury occurs, Langerhans cells tend to emigrate from the skin. In a murine skin burn injury model, Langerhans cells and mast cells were found to disappear in injured skin. 53 In mice, epidermal Langerhans cells are found to be replenished in two waves during local inflammation.
During the first wave, monocytes were found to give rise to the short-term Langerhans cells repopulation, whereas the intrinsic steady-state proliferation in situ from precursor cells located at the hair follicles facilitates the long-term repopulation process. 54 In another mouse study, the depletion of langerin+ cells was found to result in a faster reduction in the wound area, 55 suggesting an inhibitory role for Langerhans cells in wound healing.
Inversely, wound closure was significantly delayed in dendritic cells-deficient mice that was associated with significant suppression of early cellular proliferation and granulation tissue formation. 56 Aberrant wound-healing process resulting from depletion of DCs suggests that specific DC subsets are essential for skin repair although this generalization cannot be extended for LCs.
Pathological scarring
Human hypertrophic scars exhibit higher numbers of CD1a+ Langerhans cells than normal scars. 57 Specifically, accumulations of Langerhans cells were reported in the hypoxic areas of hypertrophic scars. 58 However, IL-1α produced by Langerhans cells and keratinocytes was reduced in the epidermis of hypertrophic scars, 59 which could reduce the catabolism of the dermal matrix.
Although there were no significant differences in the numbers of Langerhans cells between keloids and normal tissue, 43 higher infiltration of dendritic cells expressing CD80, CD86, and CD11c was noted in keloid lesions than in non-lesional areas or normal skin. 60 The contrasted consequences of Langerhans cells and other dendritic cells in scar formation may be based on the types of pathological scarring. However, more studies are required for elucidating the functional and pathological roles of Langerhans cells and other subtypes of dendritic cells in wound healing and the formation of pathological scars.
Neutrophils
Wound healing
Neutrophils are among the first leukocytes to arrive at an injury in response to self-damage signals (damage-associated molecular patterns [DAMPs]) or pathogenic signals (pathogen-associated molecular patterns [PAMPs]). 9 In response to PAMPs, neutrophil recruitment is likely to function as part of protective immunity. PAMPs and DAMPs activate similar pro-inflammatory responses via pattern recognition receptors, which may recruit neutrophils to the injured sites and further cause severe tissue damage via releasing oxidants, proteases, and antimicrobial proteins. 61
Neutrophils play an important role in preventing microbes from spreading, via the formation of neutrophil extracellular traps (NETs). 62 The NET components include neutrophil elastase, myeloperoxidase, and MMPs. Though NETs play a protective role by trapping microbes, the excessive release of NETs can destroy wound matrix, angiogenesis, and repairing cells that lead to immoderate inflammation and delayed wound healing. 63,64
Therefore, the overabundant neutrophils in chronic wounds indicates that blocking the number or the activity of neutrophils may be beneficial for the treatment of vicious wounds. 65 In mice, proteases such as MMP-9 were found released from neutrophils, leading to VEGF activation and promotion of revascularization at injured sites. 66 Neutrophils can also directly release VEGF to promote wound healing. 67 Physiologically, the transient recruitment of neutrophils in the early phase of wound healing is important for wound healing by clearing wound debris, reducing infection, and secreting growth factors. 68
Pathological scarring
In a prospective observational study, very few neutrophils are found to be recruited to fetal wounds that correspond to scarless healing. 69 The possibility is that neutrophils adhere less to fetal endothelial cells due to lower P-selectin expression. 70 In addition, skin biopsies were taken directly post-wounding and 3 h later from patients who received elective cardiothoracic surgery.
The scars developing into hypertrophic scars 1 year after surgery showed significantly higher P-selectin mRNA expression than those developing into normal scars even though neutrophil numbers are increased equally in both groups. 71 The adhesion by P-selectin between leukocytes and endothelial cells is required for effective recruitment of leukocytes from the circulation and into injured regions. 72 Therefore, increased P-selectin expression could mediate the recruitment of other immune cells into hypertrophic scars.
As to the circulating neutrophils in the bloodstream, another prospective study revealed that the patients who developed hypertrophic scars had lower neutrophil counts on postoperative day 5 compared with those who developed normal scars. 73 Hence, there is a need for further investigation of the mechanisms underlying the circulating and tissue-enriched neutrophils that contribute to hypertrophic scars or keloids as compared with normal scars or skins.
Mast cells
Wound healing
Mast cells are resident inflammatory cells essential for allergic responses, which present in high numbers in organs exposed to the external environment, such as skin. 74 Besides their importance in homeostasis and defense against infections in the skin, mast cells play important roles at various wound-healing stages.
When skin injury occurs in mice, the absence of mast cells leads to reduced recruitment of neutrophils in the inflammation stage likely due to reduced histamine release, 75 but it has no effect on the proliferative biological functions of wound healing, including re-epithelialization, collagen synthesis, and angiogenesis. 76 Other groups have confirmed that the depletion of mast cells in mice reduces neutrophil extravasation and recruitment to the wounded regions and interrupts the wound-healing process by disrupting the histamine pathway. 77
However, in a simple coculture system, mast cells can induce α-SMA expression and contraction force of normal human dermal fibroblasts. Interestingly, histamine treatment only stimulates α-SMA expression, suggesting that other factors could be also involved in the activation process of fibroblasts. 78 In normal human skin wound healing, expression of MCP-1 and IL-4 is correlated with the recruitment of mast cells. 79 Those factors have been shown to play a role in contractile activity of fibroblasts. 80,81 Therefore, mast cell activation, histamine release, and cytokine secretion have critical roles for fibroblast activation and cutaneous wound healing. 78,82
Pathological scarring
Fetal cutaneous wounds usually heal without scarring owing to lower inflammatory response. 26 As compared with that at embryonic day 15 (E15), the increases in cell number and granularity of mast cells are found on unwounded and wounded fetal skin of mouse at E18. Injection of mast cell lysates isolated from E15 or E18 into E15 wounds blocks scarless healing, and wounds of mast cell-deficient E18 mouse heal with smaller scars.
These results reveal that mast cells promote the transition from scarless to fibrotic wound healing in mouse fetus. 83 In another study using a hypertrophic scar mouse model induced by a third-degree burn, mast cell number was also elevated. 84 The blockade of mast cell activation can reduce inflammatory cytokines IL-1α, IL-1β, and CXCL1 as well as scar formation of the healing wound in an adult mouse model. 85 In humans, mature scar showed increased levels of mast cells relative to uninjured skin. 86
Mast cells are also more abundant in hypertrophic scars than in normal skin and are known to promote cell proliferation, and collagen synthesis of hypertrophic scar fibroblasts by activating the TGF-β1/SMAD pathway. 87 Many studies have showed the increased number of mast cells in keloid relative to normal skin. 88,89 Degranulated and mature mast cells, which co-express tryptase and OX40 ligand, are found to be greatly increased in intralesional and perilesional sites of keloids compared with normal skins and normal scars. 43
Mast cell-derived chymase contributes to cell proliferation, collagen production, and TGF-β1 expression in keloid fibroblasts. 89 Intralesional cryosurgery flattens lesional sites of keloids with a reduction in the thickness of collagen fibril as well as the cell number of fibroblasts and mast cells. 90 These studies show that mast cells are a potential target for inhibiting the development of pathological scarring. However, several studies have also reported an opposite trend for mast cells involvement in keloids.
Comparison of mast cells in human keloids and exuberant granulation tissues in horse (which was argued to have similar gross appearance) shows less mast cells in the former, although a lack of normal/control skins does not allow further verification of these numbers. 91 Another study revealed that the occurrence of toluidine blue positive mast cells in the papillary and reticular dermis in keloids was largely lower than that in normal skins, mature scars, and hypertrophic scars. 92
Since toluidine blue displays metachromatic staining of mast cells due to the presence of heparin and histamine, other mast cell populations may not have been detected. It is possible that specific subsets such as tryptase or chymase positive mast cells could also play a leading role in pathological scarring. In addition, the method of collecting of skin samples from healthy and keloid subjects, and the distance of adjacent normal skins from keloids could result in different results.
The heterogeneity of keloids among different genetic backgrounds (races) can contribute to the conflicting findings regarding the association of mast cells in keloids. Therefore, advanced studies using novel techniques and good animal models are necessary to investigate the role of mast cells in the various stages during the progression of pathological scars.
T cells
Wound healing
T cells play a pivotal role in the adaptive immune response, especially in combating neoplasms and virus-infected cells. Generally, T cells are classified into two major subpopulations, CD8+ and CD4+, and they undergo further differentiation to gain distinct functions in the periphery with the unique expression T cell receptors through somatic gene rearrangement, based on the stimuli and the inflammatory microenvironment. Among the different subtypes, CD8+ tissue-resident memory T cells (Trm cells) mainly populate the human epidermis whereas CD4+ Trm cells localize in the dermis and possess more dynamic properties. 93
Highly activated FOXP3+ (forkhead box p3) regulatory T cells (Treg cells), which are a subset of CD4+ T cells, are significantly increased in wounded skin. Tregs can attenuate the accumulation of M1 macrophages and pro-inflammatory T cells such as Th1 and Th17. 94 The specific ablation of Tregs in wounds results in delayed wound re-epithelization, implying that Tregs can facilitate skin wound healing by dampening the inflammatory response. 95
Further, a group of skin commensal-specific CD8+ T cells that are devoid of cytotoxic features execute type 17 immunity and produce type 2 cytokines (IL-5 and IL-13) to promote skin homeostasis and wound healing. 96 Therefore, different subtypes of T cells commit to the defense against pathogens and the tissue repair process.
Pathological scarring
Th1 and Th2 cells belong to CD4+ T cells, whose related cytokines have been demonstrated to be strongly associated with fibrosis formation in burn patients with hypertrophic scars. 97 In addition, the Th2 axis and/or JAK (janus kinase)/STAT (signal transducer and activator of transcription) signaling are associated with keloids. 61,98 Th2 cytokines, including IL-4 and IL-13, have been verified to enhance M2 macrophage polarization and fibroblast activation, which further promotes tissue fibrosis. 99 IL-17 can induce the protein expression of stromal cell-derived factor-1, TGF-β, collagen, and α-SMA in keloid fibroblasts through STAT3 signal transduction, hinting that Th17 cells, a subtype of CD4+ T cells, could play a role in fibroblast activation of pathological scar via secreting IL-17. 100
Scar formation in children with Bacillus Calmette-Guérin (BCG) vaccine injection and tuberculosis is correlated with Foxp3 gene expression and regulatory T cells. 101 Further, cytotoxic CD8+ T cells are found to be downregulated in keloids. 102 Both CD4+ and CD8+ T cells play contrast roles in scar formation; however, the mutually interactive mechanism should be further demonstrated in in vitro or in vivo studies.
Treatment
Conventional treatment modalities include mono- or combination therapies: local corticosteroid injections, topical corticosteroid agents, other topical agents with moisturizing effect, rest/fixation/compression therapy, surgery, and radiotherapy. 103 The mechanisms behind these strategies concentrate on the anti-inflammatory effects by switching off the pro-inflammatory signaling pathways, softening the wound matrix, and inhibiting angiogenesis.
Oxandrolone, which is usually used to treat wasting diseases, was reported to have an anti-fibrotic effect on a rabbit ear hypertrophic scar model by decreasing inflammation, vascularization, and fibroblast activation. 104 Silicone gel dressings and onion extract ointment interrupt scar formation by stabilizing mast cells. 105,106 Aspirin, a commonly used nonsteroidal anti-inflammatory drug, obstructs the TNF-α-induced activation of nuclear factor kappa B and the cell proliferation of keloid fibroblasts. 107 Anti-fibrotic and anti-inflammatory effects are also found in scars with the administration of anti-cancer drugs such as paclitaxel and 5-fluorouracil. 108,109
However, despite the plethora of potential treatments available, no single treatment has achieved better remission or prevented the recurrence of keloids or hypertrophic scars. Recently, the combination of surgery followed by radiation and corticosteroid tape/plaster use has been reported to achieve complete remission of inflammation and angiogenesis. 110
New strategies are springing up due to a deeper understanding of scar formation at the molecular level. For example, drugs targeting the TGF-β signaling pathway, in which TGF-β1 and TGF-β2 have been demonstrated to associate strongly with fibrosis, are in the preclinical phase or clinical trials for various fibrotic diseases. 111,112 As a potent immunosuppressive cytokine, IL-10 has the potential to recapitulate fetal scarless regenerative healing capacity by dampening the inflammatory wound microenvironment, TGF-β/SMAD signal pathway, and increasing ECM breakdown. 113
The successful regenerative healing process promoted by IL-10 overexpression has been demonstrated in many animal models. 114,115 Therefore, clinical trials investigating human recombinant IL-10 have been developed and a recent report have shown statistically significant improvement in macroscopic scar appearance and less red scar. 116
Moreover, the level of tumor necrosis factor alpha-induced protein 6 (TSG-6), which is a secreted 35-kDa glycoprotein, is much lower in keloids than in normal skin. Treatment with recombinant TSG-6 was found to reduce scar formation in rat ear and murine wound models by targeting the TGF-β1 response, myeloperoxidase, and inflammatory cytokines, including IL-1β, IL-6, and TNF-α, 117,118 suggesting that TSG-6 holds great potential as a biologic for scar treatment. Epidermal-released high-mobility group box 1 (HMGB1) protein activates dermal fibroblasts through receptor for advanced glycation endproducts and Toll-like receptor-2/4 receptors. Treatment with the HMGB1 inhibitor BoxA significantly mitigates scar formation in rabbit ears. 119
SUMMARY
Pathological scars profoundly affect patients both physically and psychologically. Therefore, every attempt to achieve a total cure for scars is of great importance. Scarring usually arises from aberrant wound healing after skin injury. The dysregulated inflammation leads to abnormal wound healing and may further induce pathological scarring. Molecules released by each immune cell are discussed in this article, and their targeted functions are summarized in Fig. 3.
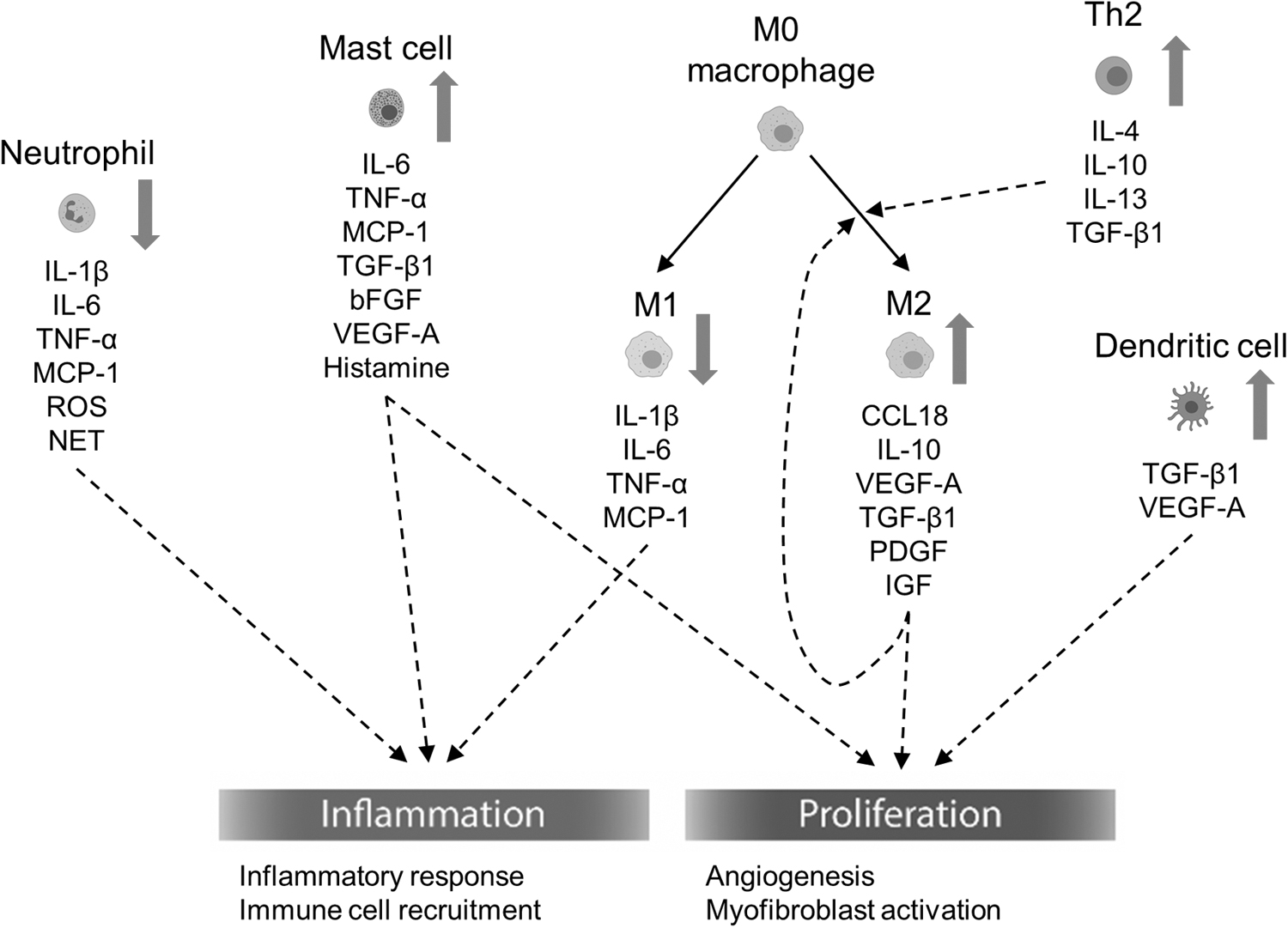
The summary of immune cells, cytokines, and growth factors contributing to wound healing and pathological scarring. The dash line indicates the effects of the immune cells, cytokines, and growth factors on inflammation and proliferation. Red arrows next to each cell type indicate the increase in cell proportion of pathological scarring respectively.
Briefly, the cell numbers or functions of dendritic cells, mast cells, M2 macrophages, and Th2 cells were increased, whereas those of neutrophils and M1 macrophages were decreased in pathological scars. Cytokines, including IL-1β, TNF-α, and MCP-1, derived majorly from neutrophils and M1 macrophages can regulate inflammatory response and immune cell recruitment in the inflammatory phase. Interestingly, the secretion of IL-6 from neutrophils, M1 macrophages, and mast cell plays a dual role in inflammatory response and myofibroblast activation.
M2 macrophages polarization is mediated by IL-4, IL-10, and IL-13 secreted by Th2 cells. TGF-β1 and VEGF-A released by mast cells, M2 macrophages, and dendritic cells facilitate myofibroblast activation and angiogenesis respectively. Effective screening for anomalous inflammatory factors and cells is, therefore, crucial to personalized therapies against pathological scarring. The effects of each immune cell subset distinguished by specific markers on wound healing, hypertrophic scars, and keloids are summarized in Table 1. The increased understanding of the complicated immune system may shed light on curing pathological scars.
The role of immune cells in wound healing and pathological scarring
TAKE-HOME MESSAGES
Innate and adaptive immune cells play distinct roles in cutaneous wound healing and pathological scarring.
Pro-fibrotic immune cells, such as M2 macrophages, dendritic cells, mast cells, and Th2 cells, promote the transition of fibroblasts to myofibroblasts, which further contribute to the formation of pathological scars.
The increased knowledge of immune system in wound healing and fibrosis provides a new insight into the management of pathological scars, such as hypertrophic scars and keloids.
Footnotes
AUTHORs' CONTRIBUTIONS
All authors have reviewed this article and agreed to publish it.
ABOUT THE AUTHORS
ACKNOWLEDGMENTS AND FUNDING SOURCES
This work was supported by a grant from the Ministry of Science and Technology, Executive Yuan, Taiwan. (MOST 109-2326-B-006-004-MY3 to Chao-Kai Hsu). The authors thank Chien-Kuo Wang and Tzu-Ying Lu for the services of drawing figures in the Department of Audiovisuals, National Cheng Kung University Hospital, Learning Resource Center, College of Medicine, National Cheng Kung University.
AUTHOR DISCLOSURES AND GHOSTWRITING
The authors have no conflicts of interest to declare. In this article, the content was written by the listed authors. There are no ghostwriters.