
Abstract
Objective:
Despite the significant function of lymphatics in wound healing, and frequent clinical use of Negative Pressure Wound Therapy (NPWT), the effect of mechanical force application on lymphangiogenesis remains to be elucidated. We utilize a murine incisional wound healing model to assess the mechanisms of lymphangiogenesis following NPWT.
Approach:
Dorsal incisional skin wounds were created on diabetic mice (genetically obese leptin receptor-deficient mice [db/db]; n = 30) and covered with an occlusive dressing (Control, n = 15) or NPWT (−125 mmHg, continuous, 24 h for 7 days; NPWT, n = 15). The wounds were macroscopically assessed for 28 days. Tissue was harvested on day 10 for analysis. Qualitative functional analysis of lymphatic drainage was performed on day 28 using Evans Blue staining (n = 2).
Results:
NPWT increased lymphatic vessel density (40 ± 20 vs. 12 ± 6 podoplanin [PDPN]+ and 25 ± 9 vs. 14 ± 8 lymphatic vessel endothelial receptor 1 [LYVE-1]+) and vessel diameter (28 ± 9 vs. 12 ± 2 μm). Western blotting verified the upregulation of LYVE-1 with NPWT. Leukocyte presence was higher with NPWT (22% ± 3.7% vs. 9.1% ± 4.1% lymphocyte common antigen [CD45]+) and the leukocytes were predominately B cells clustered within vessels (8.8% ± 2.5% vs. 18% ± 3.6% B-lymphocyte antigen CD20 [CD20]+). Macrophage presence was lower in the NPWT group. Lymphatic drainage was increased in the NPWT group, which exhibited greater Evans Blue positivity.
Innovation:
The lymphangiogenic effects take place independent of macrophage infiltration, appearing to correlate with B cell presence.
Conclusion:
NPWT promotes lymphangiogenesis in incisional wounds, significantly increasing the lymph vessel density and diameter. This study highlights the potential of NPWT to stimulate lymphatic drainage and wound healing of surgical incisions.
Adriana C. Panayi, MD

INTRODUCTION
In uncomplicated wound healing, the lymphatic network efficiently drains the protein-rich fluid from the interstitial space, maintaining fluid balance and limiting edema; up to 90% of the interstitial fluid is recycled by the lymphatic, and not by the venous system. 1 Furthermore, the lymphatic network facilitates the transport and delivery of cells mediating the immune response at the wound site. 2 Despite the lymphatic network's crucial functions—and in contrast to the vascular system whose significance in proper healing is widely accepted and described—research on lymphatic regeneration in wound healing has only recently gained momentum.
Similarly, Negative Pressure Wound Therapy (NPWT), which Argenta and Morykwas introduced in 1997, 3 has been traditionally used to treat open wounds, with use in surgical incisions only gaining attention in the past few years. In open wounds, a porous foam sponge is applied directly into the cavity and connected to a vacuum source, allowing negative pressure to be exerted on the wound. Research has identified four general mechanisms through which NPWT exerts its functions: macrodeformational force application, 4 microdeformational force application, 5 fluid removal, 6 and modification of the wound microenvironment. 7 These four main mechanisms rely on multiple secondary mechanisms, including promotion of angiogenesis 8 and neurogenesis, 9 and modification of the bioburden 10 and the inflammatory process. 11
Given the aforementioned benefits of NPWT in open acute or chronic wounds, it was theorized that the principles underlying the success of the therapy may be applicable to closed incisional wounds. 12
It is traditionally accepted that diabetic patients experience compromised wound healing, with research suggesting an association between diabetes and increased risk of postsurgical wound infections, particularly in patients with diabetes-induced peripheral arterial disease undergoing amputation. 13 An investigation of the mechanisms of NPWT in incisional wounds is, therefore, important. Diabetic mice have previously been shown to display poorer wound healing compared to normoglycemic mice in an incisional model. 14
Increased understanding of the mechanisms of action of incisional NPWT is necessary for increasing and improving the use of therapy in incisions. Previously described pathways through which incisional NPWT is believed to act include prevention of infection of the incisional site and more effective wound edge alignment and closure. Furthermore, incisional NPWT aids fluid removal and contamination elimination from the surgical site. As previously stated, other pathways, particularly those at the cellular and molecular level, such as angiogenesis and lymphangiogenesis are much less understood and should be the subject of future research.
In a recent study, our group showed that microdeformational forces applied to excisional murine wounds through continuous NPWT significantly increased lymphangiogenesis at the microscopic level, improving lymphatic drainage and translating to improved quality and rate of wound healing (In Press). We showed both an increase in the density and dilatation of the lymphatic vessels and identified an increase in the lymphatic specific receptor lymphatic vessel endothelial receptor 1 (LYVE-1). We, therefore, underlined the role of lymphatic vessels in wound healing and highlighted the promising potential of NPWT on lymphatic stimulation.
More relevant to incisional healing, in a 2011 study, Kilpadi and Cunningham applied closed incisional NPWT to a porcine model and showed that NPWT was associated with significantly decreased occurrence of hematoma and seroma, which they assigned to increased lymph clearance. 15 By using nanospheres as a proxy for fluid, a higher systemic presence of the nanospheres was seen when NPWT was applied, showing that NPWT promotes lymphatic engagement and circulation. Although the results from the Kilpadi and Cunningham study should have provided the impetus for further investigation of the effect of NPWT on the regeneration of the lymphatic system, research has been scarce.
Studies on the effects of NPWT in incisional wounds have, therefore, focused on the vascular system and wound infection prevention, with the lymphatic system receiving limited attention. Given the significant role of lymphatics in wound healing, investigation of the local lymphatic network following NPWT in incisional healing is imperative.
CLINICAL PROBLEM ADDRESSED
Clinically, excess swelling and edema can impede wound healing. Prompt and uncomplicated incisional wound closure are key to successful tissue healing following surgery, with improper closure leading to several sequelae, including infection and dehiscence, with subsequent poor esthetic and functional outcomes. Delayed wound healing has been hypothesized to be partly due to reduced lymphatic regeneration, dysfunctional drainage, and compromised compensatory mechanisms that maintain fluid homeostasis, processes resulting in persistent edema and poor recycling of inflammatory cells. 16,17
NPWT is clinically employed in the treatment of various acute and chronic wounds, surgical incisions, lower extremity wounds, and pressure ulcers, as well as traumatic and burn wounds. 3,18 –22 The molecular mechanism underlying the functions of NPWT in wound healing are complex; further exploration of these mechanisms can provide a better understanding of the therapy, potentially suggesting new therapeutic avenues, particularly with regard to lymphatic disorders.
MATERIALS AND METHODS
Animals
Diabetic and obese male mice (genetically obese leptin receptor-deficient mice [db/db]; 10 weeks; n = 30) were purchased from Jackson Laboratories (Bar Harbor, ME; Strain: BKS.Cg-Dock7m +/+Leprdb J) and acclimatized in the Brigham and Women's Hospital animal facility for 1 week before surgery. All procedures were performed in accordance with a protocol approved by the Harvard Medical School Institutional Animal Use and Care Committee (Protocol No. 2018N000117). Obese and diabetic mice were used for two reasons: (1) prior studies 23 –25 have shown that diabetic mice display delayed and inefficient wound healing and (2) obese mice have a larger dorsal surface area and are slower and much less active than wild-type mice. Given that continuous NPWT requires uninterrupted vacuum application, the larger surface area of the dorsum of obese mice, as well as their limited activity were advantageous for limited interruption to the NPWT setup.
Surgical procedure and postsurgical monitoring
Surgery was performed when the mice were 11 weeks of age. The mice underwent hair removal (Nair®) of the entire dorsum and were anesthetized in an induction chamber (2–4% isoflurane vapor). A 2 cm vertical incisional wound was created on their dorsum and the mice were separated into two treatment groups (Fig. 1). The mice in the Control group (n = 15) were covered with an occlusive dressing (Tegaderm™; 3M™, St. Paul, MN) throughout the 28-day period (Fig. 1). The mice in the NPWT group (n = 15) were treated with a setup that allowed continuous suction of 125 mmHg to be applied for 24 h/day for 7 days. Our group has previously described this setup (In Press).

Experimental setup. An incisional wound was created on the dorsum of 30 diabetic mice, which were split into 2 groups, treated either with continuous vacuum application for 7 days (NPWT group) or with an occlusive dressing (control group). The NPWT setup depicts the different layers required to secure vacuum. The timeline outlines the days of intervention: animals were photographed on days 0 (n = 15 per group), 7 (n = 15 per group), 10 (n = 15 per group), 14 (n = 5 per group), 21 (n = 3 per group), and 28 (n = 2 per group), and sacrificed on days 10 (n = 10 per group), 14 (n = 2 per group), 21 (n = 1 per group), and 28 (n = 2 per group). Evans Blue assessment was performed on day 28. NPWT, Negative Pressure Wound Therapy.
Briefly, a four-layer dressing system was used to create a tight seal for NPWT application: (1) a modified DuoDERM® dressing (Conva Tec, Reading, United Kingdom) with an adhesive window was placed over the disinfected skin; (2) a 2 × 1 × 1 cm3 of polyurethane foam (GranuFoam; Kinetic Concepts, Inc., San Antonio, TX) was placed within the window and over the wound; (3) an adhesive film (V.A.C.® Therapy System; Kinetic Concepts, Inc.) was placed over layer 1 and 2 creating a tight seal; and (4) an occlusive dressing (Tegaderm; 3M) was placed over all layers and was sutured in place (4–0 Prolene suture; Ethicon, Inc., Somerville, NJ) at four corners on the mice dorsa (Fig. 1). The tube inserted in the foam was connected to the vacuum source (V.A.C. Therapy System; Kinetic Concepts, Inc.), which exerted continuous suction (125 mmHg, 24 h/day for 7 days).
The suction apparatus, including foam and dressings, was replaced, under anesthesia, with clean materials on day 3 postsurgery. On day 7 of suction application, the apparatus was removed, and the wounds were covered with occlusive dressing alone. On macroscopic assessment and photography days, the suction apparatus (NPWT group) and occlusive dressing (Control group) were temporarily removed and replaced after imaging. When harvesting, the wound and 0.5 cm of skin surrounding the wound were collected, split into equal halves, and either cryopreserved in liquid nitrogen to be stored at −80°C for molecular analysis or fixed in 10% formalin followed by 70% ethanol for histological analysis. The number of mice per group, which were sacrificed and tissue harvested, was as follows: 10 mice per group on day 10, 2 mice per group on day 14, 1 mouse per group on day 21, and 2 mice per group on day 28.
Evans Blue dye tracing
To allow visualization of the lymphatic drainage network, on the final day of assessment (day 28), Evans Blue dye (25 μL, 4%; Sigma-Aldrich, San Francisco, CA) diluted in phosphate-buffered saline (1 μL/gbw) was subcutaneously injected in the murine wounds using a 30 G needle (n = 2 per group). Images were taken at 5-min intervals for a total of 30 min after injection and were analyzed in ImageJ® (version 1.52a; Media Cybernetics, Bethesda, MA). The dye-positive area at each time point was measured, under double-blinded conditions, by outlining the border.
Histology
Paraffin-embedded sections were stained with Hematoxylin and Eosin (HE) 26 and Picro Sirius (PS) Red Stain (Abcam, Cambridge, United Kingdom) according to the manufacturer's protocol. HE-stained sections were used to assess wound bed thickness, and PS-stained sections were used to assess collagen deposition. PS staining is a robust method for selectively highlighting collagen networks in red. 27 Collagen deposition was measured as a percentage of total red pixels to total pixel area of the wound bed using the image thresholding plugin in ImageJ software (v1.52a; Media Cybernetics, Rockville, MD).
For antibody probing, the sections were de-paraffinized, rehydrated, blocked with 1% bovine serum albumin (BSA; 1 h), and incubated with the primary antibodies (4°C, overnight) against LYVE-1 (1:200; NB100-725SS; Novus, Littleton, CO), lymphocyte common antigen (CD45; 1:200; NB100-77417; Novus), and Ki-67 (1:100; ab15580; Abcam). The following day, the sections were washed and incubated with the following secondary antibodies (1 h, in the dark): Alexa Fluor 594 (1:400; A-11007; Invitrogen, Waltham, MA) for LYVE-1 and Ki-67 and Alexa Fluor 647 (1:500; ab180117; Abcam) for CD45.
For double immunofluorescence, the slides were co-incubated (37°C, 2 h) with the primary antibodies against Podoplanin (PDPN)/gp36 antibody (1:300; ab11936; Abcam) and cluster of differentiation 31 (CD31; 1:500; NB600-1475; Novus). Secondary antibodies were Alexa Fluor 647 (1:500; ab180117; Abcam) and Alexa Fluor 568 (1:500; ab175476; Abcam), respectively.
All slides were washed and mounted with 4′,6-diamidino-2-phenylindole (DAPI) solution with antifade (MBD0020; Millipore Sigma, Burlington, MA). Images were captured with an Olympus model BX53 microscope (UCMAD3; T7, Tokyo, Japan). The stained sections were analyzed with ImageJ (version 1.52a; Media Cybernetics, Bethesda, MA) by two independent assessors under double-blinded conditions. The thresholding plugin was employed and the sliders were manually adjusted to ensure that only positively stained areas remained. 28
To measure colocalization, two thresholded images were merged using the built-in tool “Image Calculator.” All quantitative histological analysis was performed using three high-power field photos of the wound bed or border of samples from day 10 postsurgery (n = 10 per group). LYVE-1 and PDPN probing was used to assess the density and average diameter of the lymphatic vessels, CD45 to assess leukocyte presence, Ki-67 to assess cellular proliferation, and CD31 to assess the microvessel density as previously described by our group. 29 –32 PDPN and CD31 analysis were performed both manually by counting positively stained as well as automatically by measuring the immunofluorescent intensity with ImageJ.
Western blotting
The cryopreserved tissue was added to a Tissue Protein Extraction Reagent (78510; Thermo Scientific, Waltham, MA) containing protease inhibitors (11836170001; Millipore Sigma). The concentrations of supernatants were quantified and normalized using Coomassie Plus (Bradford) Assay Reagent (23238; Thermo Scientific). The samples were heated (90°C, 6 min) and loaded onto 4–12% Bis-Tris NuPAGE gels (NP0321PK2; Thermo Scientific) and the proteins from the gels were transferred to polyvinylidene difluoride (PVDF) membranes (IPVH304F0; Millipore Sigma), followed by blocking with 5% BSA in Tris-buffered saline with 0.1% Tween® 20 Detergent (TBS-T; 0.1% Tween 20; room temperature, 1.5 h).
The membranes were incubated with anti-LYVE-1 (4°C, overnight; 1:1,000, NB100-725SS; Novus) followed by horseradish peroxidase-conjugated secondary antibodies (room temperature, 45 min). Finally, the membranes were washed with TBS-T, and the blots developed with enhanced chemiluminescence. Glyceraldehyde-3-phosphate dehydrogenase (GAPDH) was used as the internal reference. Densitometric analysis was performed in ImageJ software (v1.52a; Media Cybernetics, Bethesda, MA).
Terminal deoxynucleotidyl transferase-mediated dUTP nick-end labeling (TUNEL)
TUNEL assay to detect apoptosis was performed according to the manufacturer's instructions (Thermo Fisher Scientific, Waltham, MA). Apoptotic cells were visualized using an Olympus model BX53 light microscope (Olympus UCMAD3; T7) and counted using ImageJ (version 1.52a).
Wound surface macroscopic assessment
The incisions were photographed with an SLR camera (Nikon D3100) and the photographs were assessed for evidence of infection, bleeding, necrosis, or ulceration.
Statistical analysis
Differences between the two groups were assessed using t-tests and data are expressed as mean ± standard deviation. Statistical significance was set at p-value <0.05. All statistical analyses were performed in GraphPad Prism (v8.00; MacOS GraphPad Software, La Jolla, CA).
Data and resource sharing and availability
Details on any resource employed for this study, such as the murine model, drugs, antibodies, software, and hardware, are provided throughout the article. Datasets generated and analyzed to provide the findings in this study are available from the corresponding author upon reasonable request.
RESULTS
NPWT promoted lymphangiogenesis
The NPWT group had a greater density of lymphatic vessels than the Control group, as assessed with two lymphatic specific markers, PDPN and LYVE-1. The number of LYVE-1+ vessels was significantly higher in the NPWT group (25 ± 25 LYVE-1+/high power field [HPF]) compared to the Control group (14 ± 7.9 LYVE-1+/HPF; p = 0.01; Fig. 2a, b). A similar pattern was identified when analyzing the PDPN-stained sections, with the number of PDPN+ vessels being higher in the NPWT group (40 ± 20 PDPN+/HPF) compared to the Control group (12 ± 5.9 PDPN+/HPF; p = 0.0008; Fig. 2a, f).

Lymphatic and blood vessel analysis.
PDPN immunofluorescence intensity was also higher in the NPWT group (1.7 ± 0.7 Relative Fluorescence Intensity [RFI]) compared to the Control group (1 ± 0.8 RFI; p = 0.04; Fig. 2a, g). Lymphatic vessel diameter, assessed by measuring the diameter of LYVE-1 positive cells, was higher in the NPWT group (28 ± 9 μm) compared to the control group (12 ± 2 μm; p < 0.0001; Fig. 2a, c). Upregulation of LYVE-1 was confirmed with Western blotting, with the ratio of LYVE-1 to GAPDH expression significantly higher in the NPWT group (5.9 ± 3.4) than in the Control group (1 ± 1; p = 0.03; Fig. 2d, e).
NPWT promoted angiogenesis
The NPWT group had a significantly greater blood vessel density, as assessed by counting the number of CD31+ vessels (31 ± 1 CD31+/HPF) compared to the Control group (16 ± 5 CD31+/HPF; p = 0.002; Fig. 2a, h). CD31 immunofluorescence intensity was also higher in the NPWT group (1.4 ± 0.4 RFI) compared to the Control group (1 ± 0.4 RFI; p = 0.04; Fig. 2a, i).
NPWT modified the cellular composition of the wound bed
The wound bed thickness in the NPWT mice (123 ± 45 μm) was significantly lower than that in the Control mice (253 ± 98 μm, p = 0.002; Fig. 3a, b). Collagen deposition was significantly higher in the Control mice (NPWT: 22% ± 2.1% vs. Control: 33% ± 2.8%; p < 0.0001; Fig. 3a, c).
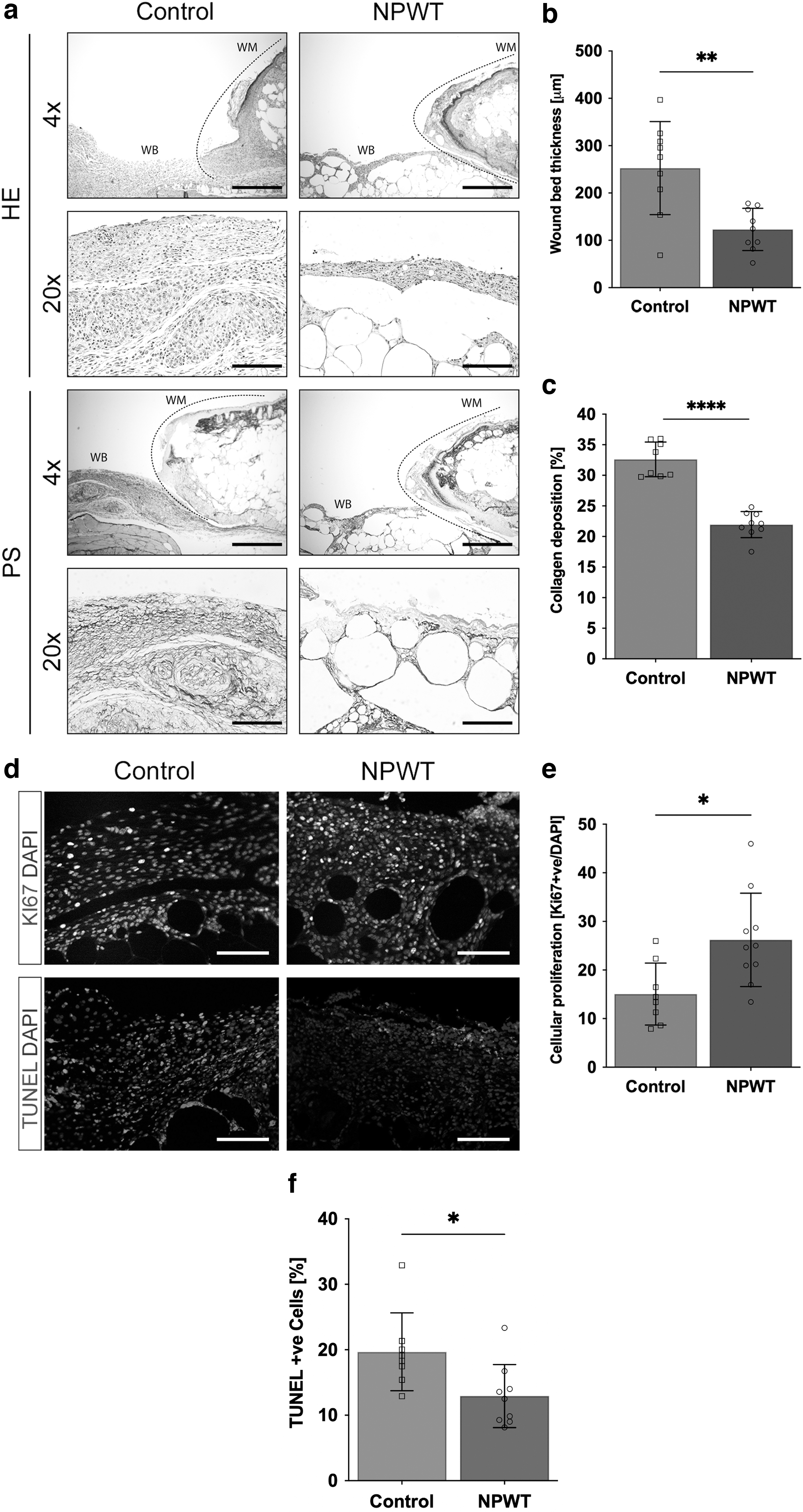
Histological analysis of the wound bed, day 10.
NPWT affected cellular turnover
Cellular proliferation was significantly higher in the NPWT group (26% ± 9.6% nuclear protein Ki67 [Ki67]+/HPF) compared to the Control group (15% ± 6.4% Ki67+/HPF; p = 0.01; Fig. 3d, e). Cellular apoptosis was significantly lower in the NPWT group (13% ± 4.8% TUNEL+/HPF) compared to the Control group (20% ± 6% TUNEL+/HPF; p = 0.02; Fig. 3d, f).
NPWT modulated immune cell presence
Leukocyte presence was significantly higher in the NPWT group (22% ± 3.7% CD45+/HPF) compared to the Control group (9.1% ± 4.1% CD45+/HPF; p < 0.0001; Fig. 4a, b), with the leukocytes appearing to be localized as clusters within vessels. Probing for B-lymphocyte antigen CD20 (CD20) identified the leukocytes to be B cells (Control group: 8.8 ± 2.5 vs. NPWT group: 18% ± 3.6% CD20+/HPF; p = 0.0001; Fig. 4a, c).
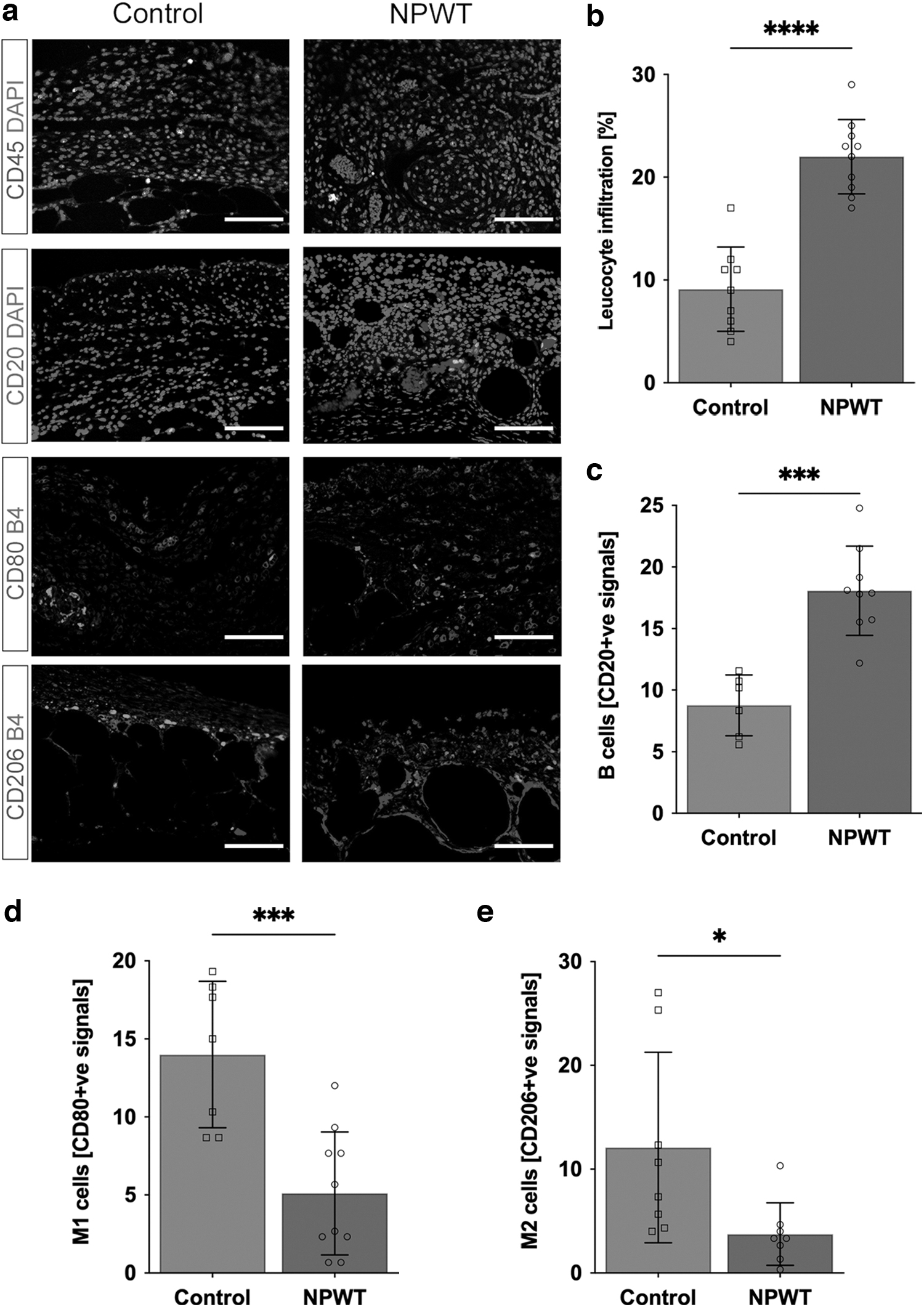
Effects on the immune cell composition of the wound bed
The number of M1 macrophages, assessed by co-staining Isolectin B4 and cluster of differentiation 80 (CD80), was significantly lower in the NPWT group (NPWT: 5 ± 3.9 cells vs. Control: 14 ± 4.7 cells; p = 0.0007; Fig. 4a, d). The number of M2 macrophages, assessed by co-staining Isolectin B4 and CD206, was also significantly lower in the NPWT group (NPWT: 4 ± 3 cells vs. Control: 12 ± 9.2 cells; p = 0.03; Fig. 4a, e). Therefore, the difference in M1 macrophage presence was greater and more significant than the difference in M2 macrophages. This can be estimated as an M1/M2 ratio of 1.3 and 1.2 for the NPWT and Control groups, respectively. Therefore, although there is overall decrease in macrophage presence, both M1 and M2 are lower and the ratio remains comparable between the two groups. Higher magnifications of the immunofluorescent sections can be seen in Supplementary Fig. S1.
NPWT improved lymphatic drainage
A qualitative assessment of lymphatic perfusion was performed. Specifically, assessment of lymphatic drainage based on the expansion of Evan's Blue positivity across a 30-min period identified differences between the two groups (n = 2 per group), with the NPWT group having a larger area of the dye-positive area at all time points (0, 5, 10, 15, 20, 25, and 30 min; Fig. 5a–c). At the endpoint (30 min), the dye-positive area in the NPWT group and Control group was 5.3 and 3 cm2, respectively (Fig. 5c). Given the lower number of animals in each group, no quantitative result was obtained and no statistical significance could be concluded.
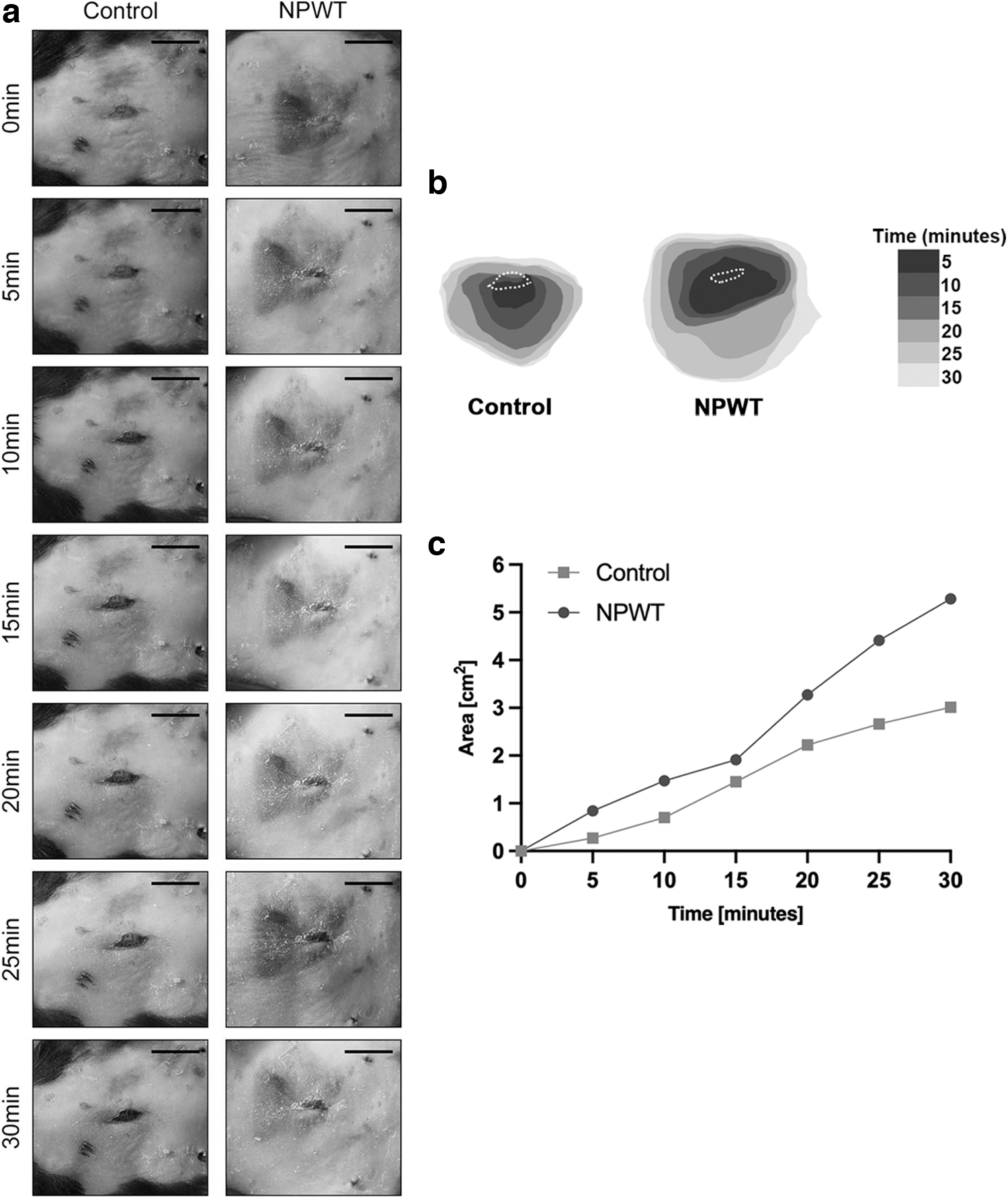
Lymphatic drainage.
NPWT was not associated with side-effects
Prior research in patients has reported possible minor side effects of NPWT use, including skin irritation, infection, and the risk of bleeding in patients using anticoagulants. 33 Although the goal of our study was not to quantitate such complications, none of the mice receiving NPWT displayed any evidence of skin irritation or bleeding (Supplementary Fig. S2).
DISCUSSION
We applied continuous NPWT to a murine incisional wound model to demonstrate that micromechanical forces improve the lymphatic network's structural regeneration, promoting lymphangiogenesis. An upregulation of LYVE-1 expression at the transcriptional level, and an increase in the density and diameter of the lymphatic vessels, contributed to an overall improvement of the wound milieu and promoted healing. The mode through which NPWT is able to stimulate local lymphangiogenesis, optimizing the network's perfusion and drainage, is likely multifactorial (Fig. 6).

Proposed mechanism underlying NPWT-induced lymphangiogenesis. Mechanosensors present on the lymphatic endothelial cells allow mechanical detection with the signals translated into molecular signals, which exert multifactorial functions on the lymphatic system. Increased osmotic tension aids fluid removal and increases lymphatic drainage. At the same time, upregulation of the LYVE-1 receptor optimizes macrophage recycling, limiting macrophage-induced inflammation. An increase in B cell presence and release of growth factors such as VEGF-A creates a positive feedback loop to induce further lymphatic growth. VEGF, vascular endothelial growth factor.
As visualized through Evan's Blue positivity assessment, lymphatic drainage was improved using NPWT. This agrees with prior research by Kilpadi and Cunningham in which they utilized a porcine wound model to assess clearance of nanosphere markers to the lymph nodes as an indicator of lymphatic drainage, and found that NPWT promoted clearance. 15 The authors also used ultrasound analysis to show that the increased lymphatic drainage was associated with a decrease in seroma and hematoma formation. 15 As the data of the Evans Blue analysis are based on two mice per group, statistical significance could not be established. The results are, however, in agreement with our prior work on excisional wounding and it is our intention to further investigate these early data in future work.
The lymphatic vessels form a conduit network that permeates leukocyte recycling, delivering B and T cells, neutrophils, and macrophages to the wound, while clearing the leukocytes during the stage of inflammation resolution. 34 Therefore, immunomodulation is an important mechanism through which NPWT appears to induce lymphangiogenesis. We found that the NPWT group exhibited a higher presence of leukocytes, with the CD45+ cells appearing to cluster within vessels. Further study identified these cells as B cells, with the NPWT group having a significantly higher presence of B cells. We show a decreased macrophage presence, both in terms of M1 and M2, compared to the control. The difference in M1, a proinflammatory macrophage phenotype, was greater than the difference in M2, a macrophage anti-inflammatory phenotype.
The estimated M1/M2 ratio was, however, comparable between the two groups. Therefore, our findings support an overall decrease in macrophage infiltration. These findings support prior research by Song et al who suggested that NPWT promotes wound healing in a diabetic murine model by suppressing macrophage infiltration. 35 The actual underlying mechanism of this phenomenon will serve as the basis of our subsequent experiments, which will focus on immunomodulation.
Similarly, Eisenhardt et al investigated the effects of NPWT in patients undergoing free flap tissue and found a significant decrease in inflammatory cell infiltration in the postischemic biopsy samples of patients treated with NPWT compared to the non-NPWT-treated muscle flaps. The authors found decreased macrophage infiltration and lower levels of inflammatory marker proteins interleukin 1β (IL-1β) and tumor necrosis factor alpha (TNFα). 36 The group also found a significant decrease in the number of apoptotic cells in the group treated with NPWT, findings that we verify in this study. 36
Increased macrophage recycling may be partly due to the upregulation of LYVE-1, a leukocyte docking receptor found specifically on lymphatic vessels that mediate macrophage, T lymphocyte, and dendritic cell trafficking. 34 The mechanism through which the LYVE-1 receptor functions relies on hyaluronan (HA), a cell surface glycosaminoglycan shown to be synthesized and assembled on macrophages' surfaces, enabling their uptake. 37 Prompt macrophage recycling is key to timely resolution of the inflammation and return to homeostasis. 38 Similarly, prior research has identified a decrease in inflammatory marker levels with the use of NPWT compared to sterile dressings in incisional wounds. 39
Overall, we hypothesize that the increased lymphatic drainage, shown in prior studies and this study with Evan's blue dye, enabled more adequate B cell presence and macrophage recycling. Increased B cell presence may itself be modulating lymphangiogenesis, as it has been previously identified to be a source of vascular endothelial growth factor (VEGF)-A, a mediator of lymphangiogenesis. 40 NPWT has previously been shown by our group to result in increased VEGF-A. 41
Shrestha et al created transgenic mice that specifically expressed human VEGF-A in B cells, and found that the VEGF-A released could induce lymphangiogenesis with the expansion of lymph nodes and development of high endothelial venules. 42 Furthermore, VEGF-A has been shown to promote antigen clearance and resolution of inflammation. 43 This mechanism, the origin and location of the different leukocytes, as well as more generally the immunomodulatory effects exerted by NPWT is an interesting research question that we aim to explore in further detail in future experiments.
Overall, this study is the first to outline the structural changes that occur in the lymphatic network when NPWT is applied to diabetic murine incisional wounds. Our findings show that NPWT promotes LYVE-1 upregulation, lymphatic vessel proliferation and dilation, and lymphatic drainage. Immunomodulation, through increased macrophage recycling and B cell presence, further regulates lymphangiogenesis.
We confirm the results from our previous study on NPWT application to excisional open wounds, further lending support of the potential of NPWT to stimulate lymphatic regeneration in different types of wounds. With regard to incisional wounds in particular, our study may help explain the effects of NPWT in surgical wounds: NPWT increases lymphatic clearance, helping to limit the formation of hematoma and seroma. At the same time, increased lymphatic and vascular perfusion has been suggested to limit surgical site infection, 44 although the mechanisms have not been elucidated, presenting an interesting future research question.
Limitations
Although our data support that NPWT was able to improve the overall quality of wound healing, as exemplified by classic features of healthy wound healing, for example, increased lymphangiogenesis and angiogenesis, increased cellular proliferation and decreased cellular apoptosis, and limited inflammation, assessment of the actual wound closure was challenging. Incisional wound closure in mice is difficult to accurately assess, especially due to alterations caused by the dressing changes and the very pliable mice skin.
However, as we have identified in previous experiments, quality of wound healing, at least in mice, does not always necessarily correlate with speed of wound closure. Furthermore, functional analysis using Evan's Blue staining was based on two mice per group. Given the low number of mice, it is not possible, at this stage, to provide concrete conclusions on the effect of NPWT on the function of the lymphatics. A further limitation, and one we will be remedying in future experiments, is the lack of additional time points, earlier on during the process of wound healing (e.g., day 5), as well as more long term, for example day 14 or 28 postwound closure.
Assessment of the mechanism, origin, and location of different immune cells, using cell markers, is an interesting research question, which we aim to explore in future experiments to fully characterize the immune response. Although we were able to show notable differences between the two groups, no statistical significance could be conferred. How the structural modifications induced by NPWT on the lymphatic system translate to functional effects, for example, through the use of Evan's blue staining, lymphoscintigraphy, and other imaging techniques, will serve as the premise for our future experiment.
INNOVATION
The effect of NPWT on the lymphatic network in incisional healing, particularly concerning local regeneration, remains to be elucidated. To address this gap in the literature, we utilized a diabetic murine model treated with continuous NPWT to study lymphangiogenesis after incisional wounding. The effects of NPWT on lymphatic regeneration are assessed at the molecular, structural, and functional levels. Ultimately, we sought to assess how modifications in the morphology and density of the lymphatic network affect the quality of wound healing.
KEY FINDINGS
The mechanisms of action of NPWT in murine diabetic incisional wounding are elucidated.
NPWT increases the lymphatic vessel density and dilation of incisional wounds.
NPWT limits inflammatory infiltration, decreasing the overall presence of macrophages, and promotes leukocytosis, with the cells identified to be predominately B cells.
Contrary to prior research, lymphangiogenesis following NPWT occurs independent to macrophage infiltration, correlating instead with B cell presence.
Footnotes
ACKNOWLEDGMENT AND FUNDING SOURCES
This work is funded, in part, by NIH 5 T35 HL11084 fellowship award to L.F., and a Gillian Reny Stepping Strong Center for Trauma Innovation award and 3M+KCI award to D.P.O.
AUTHORs' CONTRIBUTIONS
A.C.P. and D.P.O. conceptualized and designed the experiment. M.W., D.Y.M., and B.N. performed the animal experiments. M.W., D.Y.M., Z.Y., Z.C., S.K., L.F., and B.N. performed the staining, and molecular and histology analysis. A.C.P. created all images. M.W., D.Y.M., Z.Y., Z.C., S.K., O.D., and B.N. analyzed the animal data. V.H. performed the statistical analysis and created the graphs. A.C.P. wrote the article. M.W., D.Y.M., S.K., V.H., D.P.O., and A.C.P. edited the article. All authors reviewed the article.
AUTHOR DISCLOSURE AND GHOSTWRITING
D.P.O. received research funding through sponsored research agreement to Brigham and Women's Hospital from 3M+KCI. The content of this study was exclusively contributed and written by the authors listed, and no ghost writer was involved in the writing of this article.
ABOUT THE AUTHORS
SUPPLEMENTARY MATERIAL
Supplementary Figure S1
Supplementary Figure S2
Abbreviations and Acronyms
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.