
Abstract
Objective:
Impairments in the differentiation and migratory capacity of epidermal stem cells (ESCs) are pivotal factors contributing to delayed wound healing. High mobility group box1 (HMGB1) has recently emerged as a potential target for tissue repair. Therefore, we aimed to investigate the role and molecular mechanisms of HMGB1 in ESCs during the wound-healing process.
Approach:
Initially, we examined the expression of HMGB1 and the differentiation of ESCs in normal skin, normal wounds and chronic wounds. Then, we assessed the ESC migration and differentiation, and the key markers in the Wnt/Notch signaling pathways, after treatment of HMGB1 and inhibitor, and the knockdown of toll-like receptor 4 (TLR4), using scratch assay, qPCR, western blotting, and immunofluorescence. Finally, we conducted mice models to analyze the healing rates and quality in vivo.
Results:
HMGB1 was decreased across all epidermal layers, and the differentiation of ESCs was hindered in diabetic foot ulcer. In vitro, HMGB1 enhanced both the migration and differentiation of ESCs while stimulating the expression of the Wnt/Notch pathway within ESCs. However, the downregulation of TLR4 negated these effects. Finally, our in vivo experiments provided evidence that HMGB1 facilitates wound healing and epidermis differentiation via TLR4 and Wnt/Notch signaling pathways.
Innovation:
This study innovatively introduces HMGB1 as a novel target for skin wound healing and elucidates its mechanisms of action.
Conclusions:
HMGB1 accelerated wound healing by promoting the differentiation of epidermal stem cells through the “HMGB1-TLR4-Wnt/Notch” axis, which reveals a new potential mechanism and target to expedite wound healing.
INTRODUCTION
The skin, as a vital organ of the human body, directly interfaces with the external environment and is susceptible to damage from various detrimental factors, leading to a disruption of its integrity and the formation of wounds. 1 Large skin defects or various pathological conditions may all lead to delayed or non-healing skin wounds, which severely impact the physical and mental health of patients and diminishes their quality of life, resulting in a significant economic burden. 2 Therefore, it remains critically important to explore and discover new therapeutic targets and to achieve precise treatment approaches.
Wound healing is a complex but well-orchestrated biological event, with interplay between different tissue structures and a large number of resident and infiltrating cell types, including epidermal stem cells (ESCs). 3 ESCs, classified as adult stem cells, are primarily located in the basal epidermis. They possess the remarkable capacity for unbounded proliferation and differentiation into various lineages. The proliferation, differentiation, and migration of ESCs into keratinocytes are critical steps in the epidermalization process, which plays a pivotal role in wound healing. 4,5 However, the exact mechanisms underlying ESC activation during wound healing have not been thoroughly investigated.
High mobility group box 1 protein (HMGB1) is a high-mobility group protein containing the HMG-box domain, encoded by the HMGB1 gene, 6 and plays an important role in immune response, 7 tumor occurrence and development, 8 and tissue regeneration. The role of HMGB1 in the process of wound healing remains a subject of debate. Previous research indicated that the expression of HMGB1 in the blood of diabetics is elevated, contributing to immune system inflammation. 9 However, the local events within the tissue and those occurring in the bloodstream are not precisely identical. When focusing on local tissues, more studies suggest that HMGB1 may facilitate cell differentiation and migration, thereby promoting tissue repair. Topical application of HMGB1 in STZ-induced diabetic mice has been shown to increase skin’s small vessel density at the wound site, promote granulation tissue formation, and expedite wound healing. 10 In addition, extracellular HMGB1 has been implicated in the wound-healing process of the myocardium 11 and bronchial epithelium. 12 Furthermore, HMGB1-modified mesenchymal stem cells may mitigate radiation-induced vascular injury due to their high mobility and promotion of endothelial differentiation. 13 The HMGB1/RAGE axis also safeguards keratinocytes from apoptosis by encouraging cell autophagy post-injury 14 and promotes the healing of maxillary wound surfaces. 15 All these researches suggest a potential role for HMGB1 in the process of tissue repair.
Recent studies have highlighted the involvement of toll-like receptor 4 (TLR4) in the action of HMGB1, mediating the regulation of inflammation and the remodeling of tissue repair. 16 –18 In addition, TLR4 signaling also plays a crucial role in maintaining the niche of ESCs, 19 driving their proliferation and facilitating wound healing. 20 Mice lacking TLR4 exhibit significantly delayed wound healing, with impaired keratinocyte migration. 21,22 Notably, it has been reported that TLR4 can interact with Wnt/β-catenin 23 and Notch pathway, 24 which also play vital roles in wound healing and ESCs function. 25
Based on the above potentials, we assumed that HMGB1 may accelerate wound healing by promoting the differentiation of ESCs via the “HMGB1-TLR4-Wnt/Notch” axis. To prove this hypothesis, we examined the role of HMGB1 and its downstream signaling molecules in wound repair in ESCs and C57 wound healing models. This study seeks to shed light on the intricate mechanisms underlying the potential therapeutic benefits of HMGB1 in promoting efficient wound healing.
INNOVATION
The progression of a wound into a refractory state poses a persistent challenge in clinical practice. This study explored the impact and mechanism of HMGB1 on acute wounds and ESCs, uncovering the stimulatory effect of the “HMGB1-TLR4-WNT/Notch” signaling axis on the biological function of ESCs. This initial revelation underscores HMGB1’s potential in enhancing wound healing. It lays the groundwork for further clinical investigations into HMGB1’s role in wound healing and opens up new avenues for drug treatment and the research and development of therapies for clinically refractory wounds.
CLINICAL PROBLEM ADDRESSED
The prolonged or non-healing of wounds significantly affects patients’ physical and mental health and imposes a substantial economic burden on their families and society. Impairments in the differentiation process of ESCs and a diminution in their migratory capacity are pivotal factors contributing to delayed wound healing and the development of chronic ulcers. 2,4,5 HMGB1 has garnered attention as a promising target for tissue repair in recent years. 12,26 In this study, we showed that HMGB1 was decreased across all epidermal layers, and the differentiation of ESCs was hindered in chronic wounds. In addition, HMGB1 could accelerate wound healing by promoting the differentiation of ESCs through the “HMGB1-TLR4-Wnt/Notch” axis in vitro and in vivo. This evidence illuminates a new approach and therapeutic target for enhancing wound healing.
MATERIALS AND METHODS
Electronic laboratory notebook platform was not used.
Skin biopsies
After Local Ethical Committee approval and signed informed consent (IRB approval No. [2020] 292), skin biopsies were obtained from normoglycemic controls, including three cases of normal skin (NS) from non-diabetic volunteers: two were chest skin tissues from breast reduction surgery, and one was abdominal skin tissue from abdominoplasty, and three cases of acute wounds (two at D7 and one at D5 after normal full-layer skin defect, normal wound [NW]). In addition, specimens from three cases of diabetic foot ulcer (DFU) on the dorsum of the foot, one case of post-chemotherapy ulcer, one case of pressure sore, and one case of post-traumatic hematoma ulcer (representing chronic wounds, CW) were collected. The specimens comprised both wound base and margin tissues within a 0.5 cm distance from the margin (Fig. 1A).
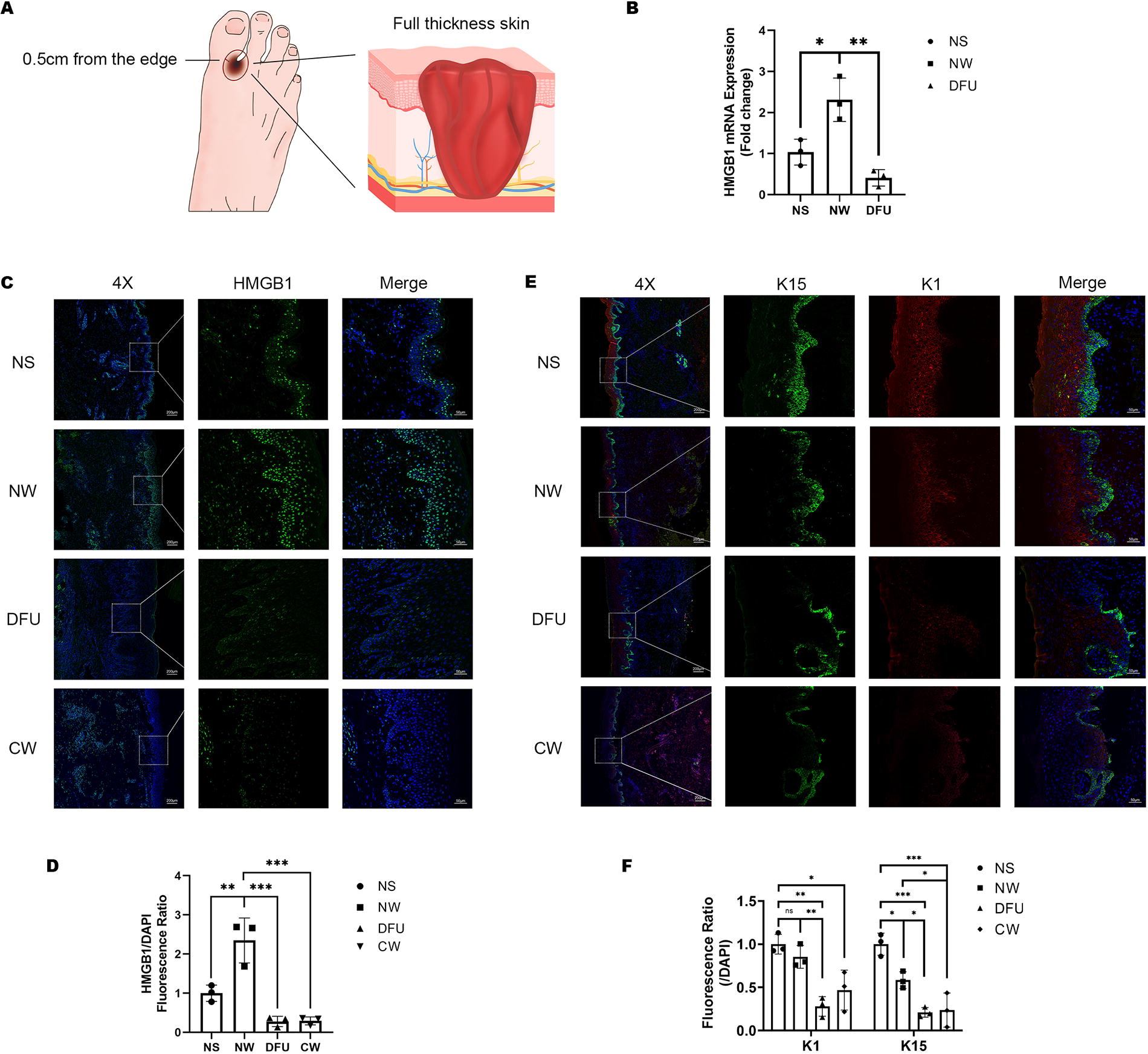
Increased expression of HMGB1 in acute and chronic wounds.
Cell isolation and culture
Human skin tissue samples were obtained from The First Affiliated Hospital of Sun Yat-Sen University, and ethical approval and informed consent were obtained. ESCs were isolated from human foreskins as described. 27 Briefly, subcutaneous tissues and fat were removed before the skin was disinfected and incubated overnight with DispaseII (3 mg/mL; D4693-1G, Sigma-Aldrich) in keratinocyte serum-free medium (Epilife, MEPI500CA, Gibco) with HKGS (S0015, Gibco) at 4°C to separate the epidermis from the dermis. After that, the epidermis separated from the foreskins was digested with prewarmed Trypsin/EDTA (0.05%, Gibco) at 37°C for 10 min to obtain ESCs, which were resuspended in Epilife and seeded at a density of 105 cells/cm2 in 100 μg/mL type IV collagen-coated culture dishes. Cells adhering within 10 min were selected according to the rapid attachment method. 28 Cells at the 3–6 passages were used for this study.
Flow cytometric analysis
To identify ESCs, about 106 cells were collected, prepared as single-cell suspensions, and incubated with 2.5 μg Fc block in 100 μL PBS. Surface marker anti-CD49f-FITC (1:100, 313605, Biolegend, USA) was used to stain cells at room temperature for 30 min. For intracellular staining, cells were permeabilized and fixed with the intracellular staining kit (554714, BD Biosciences) and then washed and stained with antibodies anti-cytokeratin 15 (1:100, ab52816, Abcam) at room temperature for 30 min. After washing, cells were incubated with secondary antibodies (1:500, 4408S, CST) and washed twice before resuspending. The stained cells were analyzed using a CytoFlex flow cytometer (Beckman, USA). Data were processed by FlowJo Software v10.8.1.
Cell viability assays
Cells were plated in 24 wells at a density of 105 cells/cm2 in a 37°C and 5% CO2 incubator. After 24 h, living and dead cells were detected using the Calcein/PI Cell Viability/Cytotoxicity Assay Kit (C2015S, Beyotime, China) following the manufacturer’s instructions. Then the cells were observed under an inverted fluorescence microscope (OLYMPUS IX83, Japan).
Western blot analysis
Protein samples were lysed using RIPA lysis buffer supplemented with protease inhibitors (Calbiochem, USA). Thirty micrograms of proteins were loaded and separated by 10% SDS-PAGE and then transferred to polyvinylidene difluoride membranes (PVDF, Millipore, USA). The membrane was blocked with 5% BSA and then incubated with one of the primary antibodies: anti-cytokeratin1 (1:500, PA5-113746, ThermoFisher), anti-cytokeratin 15 (1:10000, ab52816, Abcam), anti-TLR4 (1:1000, 19811–1-AP, Proteintech), anti-Wnt1 (1:1000, 27935–1-AP, Proteintech), anti-β-catenin (1:5000, 51067–2-AP, Proteintech), anti-Notch1 (1:1000, ab52627, Abcam), anti-Hes1 (1:1000, A0925, Abclonal), anti-c-myc (1:1000, D84C12, Cell Signaling Technology), anti-GAPDH (1:2000, 10494–1-AP, Proteintech) and anti-α-tubulin (1:5000, GB11200, Servicebio) overnight at 4°C. Afterward, the PVDF membranes were incubated with HRP-conjugated secondary antibodies at room temperature for 1 h. The immunoreactive bands were visualized with the enhanced chemiluminescence kit (Sigma-Aldrich) and analyzed by densitometry using ImageJ software.
RNA extraction and real-time PCR assay
Total RNA from cells was extracted using Trizol reagent (Thermo Fisher) and reverse-transcribed into cDNA using the PrimeScript RT reagent kit (Takara) according to the manufacturer’s protocol. Quantitative real-time PCR (qPCR) reaction was performed with the Roche 480 system (Roche) using the LightCycler 480 SYBR Green I Master Mix (Roche). Primers used for this study can be found in Table 1. The relative mRNA expression levels of target genes were calculated using the 2−ΔΔCt method.
qPCR primers used for this study
Scratch assay
The number of 2 × 105 cells were seeded into a six-well plate and the scratch assay was conducted when the cells grew to 100% of the six-well plate area. The day before the cell scratch assay, the cells were treated with 1 μg/mL of mitomycin C for approximately 8–10 h to inhibit cell proliferation. A yellow sterile pipette tip was then used to scrape the cellular monolayer. The associated pictures were taken at 0, 24, and 48 h after incubation.
Cell transfection
A total of 1 × 105 cells were seeded into each well of a six-well plate, and the medium in the plate was gently agitated to ensure even distribution of the cells. Transient transfection was performed when the cells grew to 40–50% of the six-well plate area. Before transfection, the old medium in the six-well plate was replaced with a fresh complete medium. A total of 150 μL Opti-MEM (Gibco) and 3 μL Lipofectamine 3000 (Invitrogen) were taken from each well and gently mixed in EP tube A. A total of 150 μL Opti-MEM and an appropriate amount of siRNA (final concentration of siRNA was 50 nM) were taken from each well and gently mixed in EP tube B. Gently blowing the liquid in tube A, it was slowly added to tube B, letting it stand at room temperature for 5 min, waiting for the liposomes to fuse with siRNA. Finally, the mixed liquid was slowly dropped into each well of the six-well plate, and the cells were placed back in 37°C, 5% CO2 incubator for further culture. The culture medium was changed after 8 h, and the next experiment was carried out after 48–72 h culture.
In vitro immunofluorescent staining
A total of 104 ESCs were resuspended in 500 μl Eplife and added into one well of an eight-well removable glass chamber slide (Nunc Lab-TekIIChamber Slide System, Thermo Fisher Scientific). After each group’s treatment for 48 h, cells were fixed with 4% formaldehyde, permeated with 0.1% Triton X-100 (Solarbio, China) for 15 min, and blocked with 2% BSA for 1 h. Next, the cells were stained with primary antibodies, including anti-cytokeratin1 (1:100, 16848–1-AP, Proteintech), anti-cytokeratin 15 (1:100, ab52816, Abcam), anti-TLR4 (1:100, A5258, Abclonal), anti-Wnt1 (1:100, 27935–1-AP, Proteintech), anti-β-catenin (1:500, 51067–2-AP, Proteintech), anti-Notch1 (1:100, ab52627, Abcam), anti-Hes1 (1:100, A09257, Abclonal), and anti-c-myc (1:400, D84C12, Cell Signaling Technology) overnight at 4°C. Cells were washed and incubated with secondary fluorescence antibodies (1:2000, 4408S, 4409S, 4412S, 4413S, Cell signaling Technology) for 1 h and DAPI for 10 min at room temperature. Then the cells were observed under an inverted fluorescence microscope (OLYMPUS IX83, Japan). Quantitative analysis of fluorescence intensity was conducted on a single field of view from three independent experiments.
Histological examination
Skin tissue was fixed overnight in 4% paraformaldehyde at room temperature and embedded in paraffin. Afterward, 5-μm-thick slices were cut out. Some of them were stained with hematoxylin-eosin (H&E) and Masson and analyzed using a microscope (Kfbio, China). Others were blocked with goat serum and then stained with primary antibodies, including anti-HMGB1 (1:1000, ab18256, Abcam), anti-cytokeratin1, anti-cytokeratin 15, anti-TLR4, anti-Wnt1, anti-β-catenin (1:100, A0316, Abclonal), anti-Notch1, anti-Hes1 and anti-c-myc overnight at 4°C. Sections were washed and incubated with HRP-conjugated secondary antibodies (1:2000, ab6728, ab6721, Abcam) for 1 h, fluorochrome (15X, ASOP520, ASOP570, Asbio) for 10 min, and DAPI for 10 min at room temperature. Slides were scanned afterward (Pannoramic MIDI, 3DHISTECH, Hungary) and analyzed using CaseViewer 2.4. The integrated fluorescence intensity index was calculated.
Wound model
Male C57 mice were obtained from Guangzhou Ruige Biological Technology Corporation. All animal-related procedures were conducted in strict accordance with the approved institutional animal care and maintenance protocols. Experimental protocols were approved by the Laboratory Animal Welfare and Ethics Committee of The First Affiliated Hospital of Sun Yat-sen University, (ethics approval No. [2020] 010). Full-thickness skin wounds that were 1 cm in diameter were established on the dorsum of mice (6–8 weeks). A donut-shaped splint was used to prevent contractions. 29 The wound was covered with a transparent semi-occlusive dressing (Tegaderm, 3M, Saint Paul, MN, USA) to prevent desiccation. The above wounds were randomly divided into six groups. Saline with HMGB1 (400 μg per mouse in 20 μl of saline, HMGBiotech), saline with HMGB1-BOXA (400 μg per mouse in 20 μl of saline, HMGBiotech), PBS with TAK-242 (3 mg/kg, MCE), TAK-242 and HMGB1, and PBS and saline were injected around the wound following the four-point injection, respectively, on the day of wound creation. Animals were photographed on days 0, 3, 5, 6, 7, 10, and 14 after injury. For each sample, the rate of the healing process was measured as a ratio of the area at each time point divided by the area at time 0 (that is, immediately after the wound). Mice were sacrificed at 3 days, 5 days, 7 days, 10 days, and 14 days after injection.
Transepidermal water loss
Transepidermal water loss (TEWL) was measured using a noninvasive Tewameter® TM Hex (Courage + Khazaka Electronic GmbH, Germany). The TEWL was measured by gently placing the probe head on the skin and pressing the probe button. The TEWL measurements were made 10 s after the probe was applied and when the TEWL level had stabilized. The measurements lasted for 30 s, and the average of the last 20 s was taken as the result. The values are recorded in g/m2/h.
Data analysis
All data are presented as the mean ± SD. Comparisons of changes in levels of signaling component expression between control and experimental groups at the same time point were conducted by using the Student’s t-test. The differences between the groups at different time points were compared by one-way analysis of variance followed by the Bonferroni test. Statistical analyses were performed with GraphPad Prism 8.3.0 software (GraphPad Software, USA); p < 0.05 was considered statistically significant
RESULTS
Expression of HMGB1 in acute and chronic wounds
To investigate the expression of HMGB1 in acute and chronic wounds, we procured specimens that included three cases of NS tissues, three cases of acute wound tissues obtained within 7 days of normal full-thickness skin defects (NW), three cases of granulation tissues from chronic DFUs on the dorsum of the foot. The specimens comprised both base and margin tissues within a 0.5 cm distance from the margin (Fig. 1A). We found that compared with normal skin tissues, HMGB1 mRNA was significantly increased in normal acute wound tissues, while it was significantly decreased in diabetic chronic wound tissues (Fig. 1B). Hence, we hypothesized that HMGB1 might exert a significant influence on wound healing. To delve deeper into this hypothesis, we collected samples from three chronic wounds (refer to the Methods section for specifics) and proceeded with multiplex immunohistochemistry experiments. Results showed that compared with NS tissue, the expression of HMGB1 was significantly increased in normal acute wound tissue, especially in the differentiated layers of the epidermis. On the other hand, the expression of HMGB1 was significantly decreased in all layers of the epidermis in chronic wounds (Fig. 1C–D). In terms of the differentiation of ESCs, it can be observed that in NS, K15 was mainly enriched in the basal layer, indicating good stemness of ESCs. K1, on the other hand, was mainly expressed in the differentiated layers of the epidermis, indicating good differentiation of ESCs. Compared with NS tissue samples, the expression of K15 in the basal layer decreased in both acute wounds and chronic wounds, suggesting the trauma stimulated the differentiation of ESCs. However, the expression of K1 in acute wounds was significantly higher than in chronic wounds, revealing the impaired differentiation process of ESCs in chronic wounds (Fig. 1E–F).
HMGB1 promoted the migration and differentiation of ESCs
The primary ESCs were isolated from human skin tissues, displaying high proliferative capacity, and forming colonies. Flow cytometry analysis revealed that ESCs were positive for integrin cd49f (98.1%) and Krt15 (94.3%) (Supplementary Fig. S1). HMGB1 and its inhibitors (including HMGB1-BOXA and glycyrrhizic acid) showed no significant effect on the proliferation of ESCs (Supplementary Fig. S2). However, HMGB1 effectively promotes the migration of ESCs, with the optimal effect observed at a concentration of 100 ng/mL while HMGB1-BOXA (hereinafter referred to as “Inhibitor”) significantly inhibits the migration of ESCs (Fig. 2A–B). Further, HMGB1 at a concentration of 100 ng/mL upregulates the expression of K1, loricrin, and involucrin (makers of keratinocytes at different stages of differentiation) in ESCs, while downregulating the expression of K15 (stemness marker of ESCs), indicating the promoting effect of HMGB1 on the differentiation of ESCs. Conversely, its inhibitor significantly downregulates the expression of differentiation markers, while upregulating the expression of K15 (Fig. 2C). These findings were corroborated with western blot (Fig. 2D–E) and immunofluorescence (Fig. 2F–I).

HMGB1 promoted the migration and differentiation of ESCs.
HMGB1 activated ESCs through TLR4
HMGB1 can bind to TLR4 and exert biological functions such as promoting the release of immune factors. 7 According to the prediction from the Search Tool for the Retrieval of Interacting Genes (“https://string-db.org/”), 30 HMGB1 may interact with the Wnt/Notch pathway through TLR4 (Supplementary Fig. S3). Thus, we detected the expression of TLR4 after the addition of HMGB1 and found that HMGB1 could indeed promote the expression of TLR4 (Fig. 3A–D). Therefore, we used siRNA to knock down TLR4 on ESCs (siRNA-1 showed the best efficacy, Supplementary Fig. S4, Fig. 3F–G), resulting in a slight inhibition of cell migration, and the effect of promoting ESC migration by HMGB1 disappeared when reintroduced (Fig. 3H–I). Similarly, knocking down TLR4 resulted in weakened differentiation of ESCs, and when HMGB1 was reintroduced, we found that its effect in promoting K1 expression and reducing K15 expression in ESCs was abolished (Fig. 3J–M). Results suggest that HMGB1 induces ESCs migration and differentiation by targeting the TLR4 receptor.
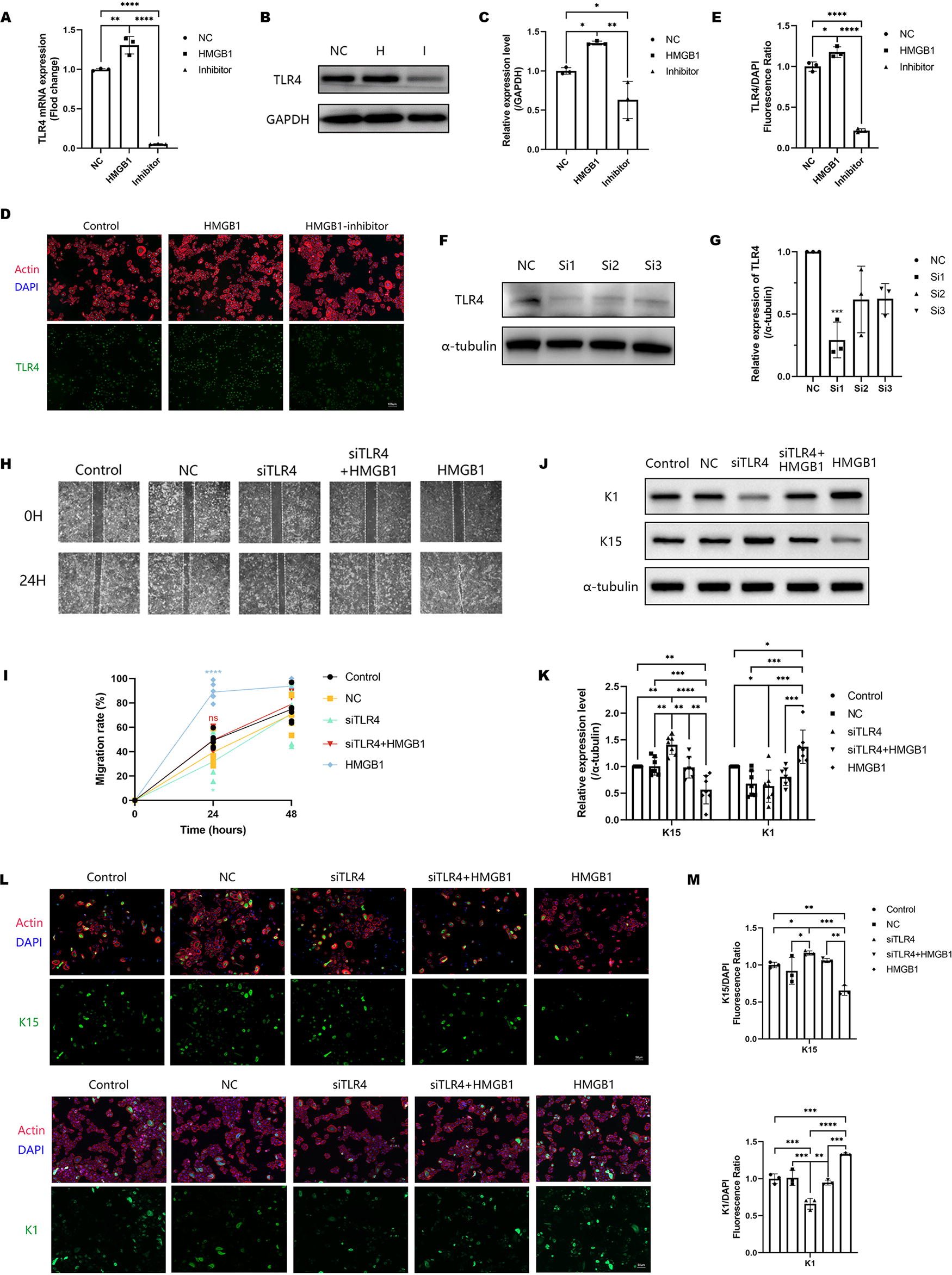
HMGB1 promoted ESCs differentiation through activating TLR4.
HMGB1 activated the Wnt/
β
-catenin and notch/Hes signaling pathways through TLR4
Our previous study revealed that Wnt/β-catenin and Notch/Hes pathways may promote wound healing by stimulating ESCs. 31 It has been reported that TLR4 can interact with Wnt/β-catenin 24 and Notch pathway. 32 Hence, to elucidate whether Wnt/β-catenin and Notch/Hes pathways were involved in the function of HMGB1, we detected the expression and localization of key proteins involved in the Notch/Hes signaling pathway, including Notch1 and Hes1, as well as key proteins involved in the Wnt/β-catenin signaling pathway, such as Wnt1, β-catenin, and c-myc, after the treatment of HMGB1. We found that HMGB1 significantly promoted the expression of Notch1, Hes1, Wnt1, β-catenin, and c-myc in ESCs (Fig. 4A–E), with β-catenin exhibiting pronounced nuclear translocation and enrichment (Fig. 4F–G), indicating the effective activation of both the Notch/Hes and Wnt/β-catenin signaling pathways by HMGB1. After knocking out TLR4, the stimulatory effect of HMGB1 on key proteins disappeared (Fig. 4H–K), and the nuclear translocation of β-catenin decreased (Fig. 4L–M), which indicates that HMGB1 indeed activates the Notch/Hes and Wnt/β-catenin signaling pathways through TLR4.

HMGB1 activated the Wnt/β-catenin and Notch/Hes signaling pathways through TLR4.
HMGB1 accelerated wound healing through TLR4
To further elucidate the function of HMGB1 in wound healing in vivo, C57 mice were divided into five groups, respectively were PBS with saline (NC), PBS with HMGB1 (400 ng per mouse), PBS with HMGB1-BOXA (400 ng per mouse), saline with TLR4 inhibitor TAK-242 (3 mg/kg), and a combination of TAK-242 with HMGB1. The reagents above were administered through a single injection around the wound site using a four-point injection method on the same day the wounds were created. Comparing the results to the saline-treated group, it was observed that the group treated with HMGB1 exhibited faster wound healing, with significant improvement observed as early as three days after wound creation. Conversely, wound healing was significantly delayed in the groups treated with HMGB1-BOXA and TAK-242, and this delay persisted for up to seven days after treatment (Fig. 5A–B). TEWL serves as an in vivo indicator of skin barrier function, providing a means to evaluate the normal differentiation of keratinocytes. 33 Consequently, the TEWL assay was conducted on day 14 post-injury to quantitatively assess the restoration of skin barrier function after wound healing, revealing that HMGB1 effectively facilitated wound barrier repair, a function that was nullified following TLR4 inhibition (Fig. 5C). To further evaluate the quality of wound healing, we observed the re-epithelialization by H&E staining. Detailed histopathological analysis showed that HMGB1 significantly enhanced the epidermal tongue crawling of the wound surface (Fig. 5D–E). While no significant differences were noted in the thickness of the newly formed epidermis (Supplementary Fig. S5), HMGB1 exhibited more pronounced effects on epidermal stratification, leading to a clearer and more regular cell layer. In addition, notable dyskeratosis was observed in the groups treated with the BOXA inhibitor and TLR4 inhibitor (Fig. 5D), suggesting inhibition of keratinocyte differentiation and stratification. In addition, the inhibition of TLR4 appeared to weaken the effect of HMGB1 in promoting epidermal differentiation. Further immunofluorescence staining analysis revealed that HMGB1 significantly stimulated the expression of the Wnt/Notch pathway and promoted the differentiation of ESC in vivo, which could be partially reversed by TLR4 inhibitor (Fig. 5F–G).
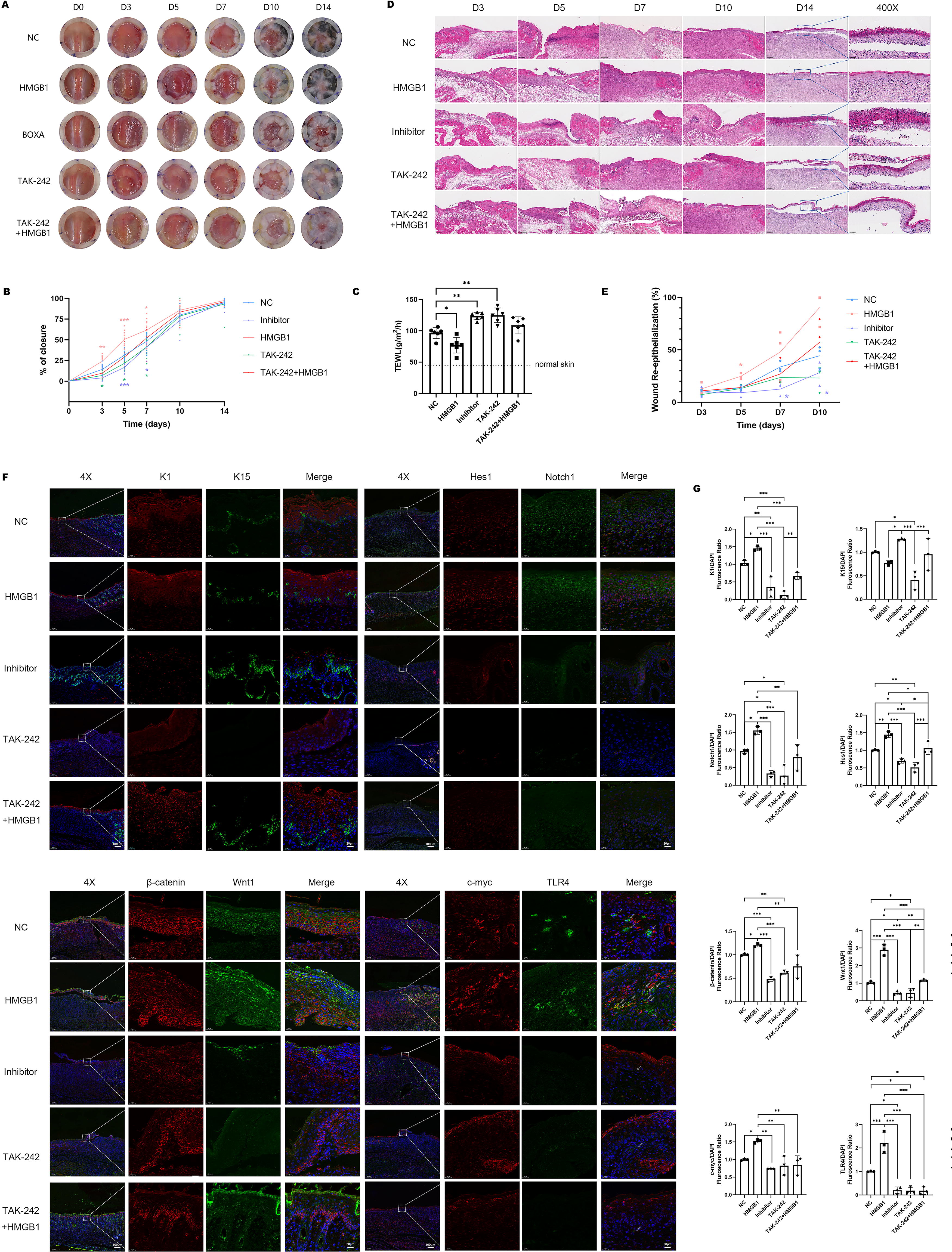
HMGB1 accelerated skin wound healing through activating TLR4.
DISCUSSION
The healing of wounds is a complex and orderly biological process that involves cell migration, wound contraction, formation of granulation tissue, angiogenesis, re-epithelialization, and matrix remodeling. Any impairment or dysfunction in these crucial processes can lead to delayed wound healing and the development of chronic ulcers. Studies in the field of skin development biology have shown that ESCs are crucial sources for skin and its appendages in terms of development, repair, and remodeling, playing an important role in maintaining epidermal renewal and promoting wound healing. 34 ESCs typically originate from the basal layer, undergo differentiation into mature cells, and gradually form layers to thicken the epidermis. 35 In this study, we found that traumatic stimuli effectively stimulate the differentiation of ESCs. However, in chronic ulcers, the differentiation process of ESCs was hindered. This may be related to the lack of HMGB1 in chronic wounds. Unlike in blood, HMGB1 expression is markedly diminished in diabetic wounds and various types of chronic ulcers. Immunofluorescence localization results indicate a significant decrease in HMGB1 expression in epidermal keratinocytes surrounding wounds, possibly contributing to slow wound contraction, delayed re-epithelialization, and thin and fragile regenerated skin.
Extracellular HMGB1, as an inflammatory stimulus factor, can participate in promoting cellular differentiation, migration, and the process of repair. 15 However, the precise influence of HMGB1 on the migration and differentiation of ESCs remains unclear. Therefore, in this experiment, we employed the well-established rapid attachment method and successfully isolated ESCs from the normal foreskin. These cells exhibited good viability, a regular round shape, clustered arrangement resembling “cobblestones,” and high expression of ESC markers, such as cd49f and K15. Furthermore, we introduced varying concentrations of HMGB1 into the culture of ESCs and performed proliferation, migration, and differentiation assays. It was observed that while HMGB1 did not exert a significant impact on the proliferation of ESCs, it effectively promoted their migration and differentiation. Regarding the expression and localization of HMGB1 in the connective tissue, we speculate that in normal acute wounds, keratinocytes produce more HMGB1 due to the stimulus from the injury. This stimulates the migration and differentiation of basal layer ESCs, thereby promoting re-epithelialization, which is a critical step in wound healing (Fig. 6A). Conversely, the reduced production of HMGB1 by keratinocytes may be a key factor contributing to the prolonged non-healing process in chronic wounds.
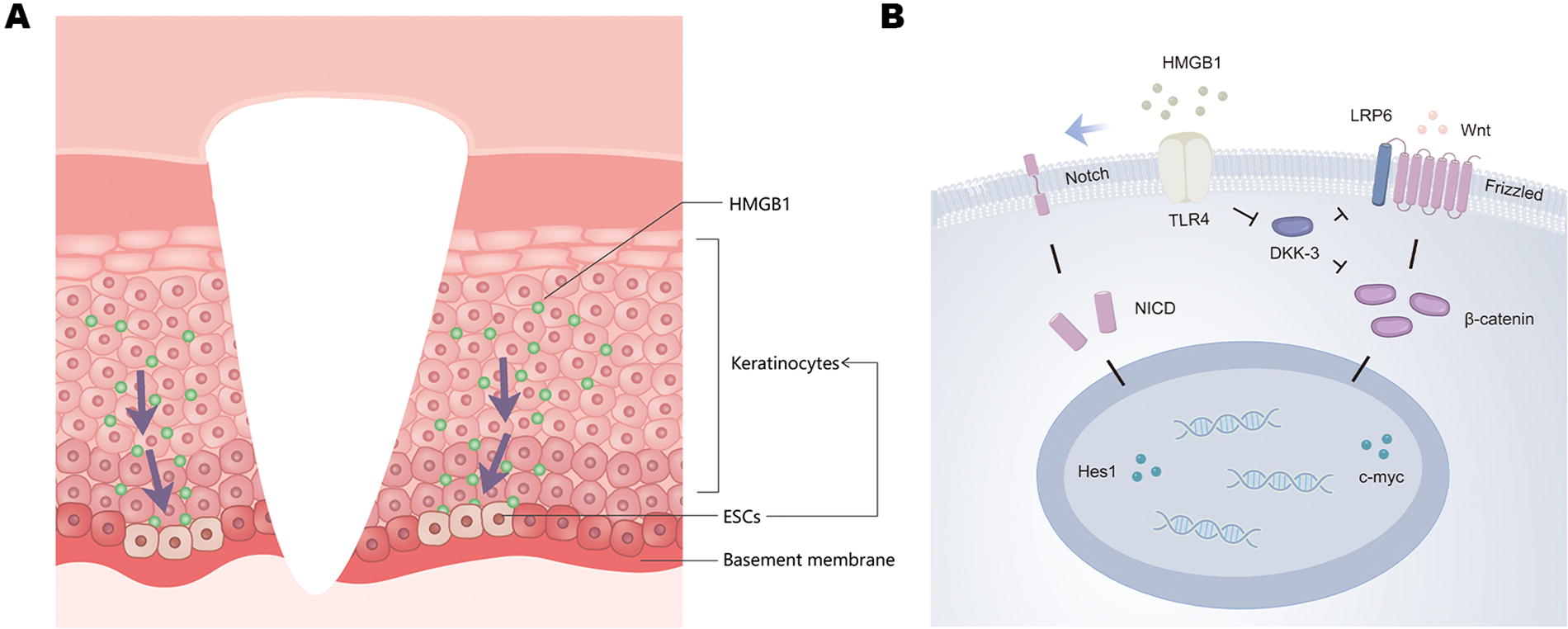
Schematic diagram illustrates the production of HMGB1 and the stimulation of ESCs
TLR4 serves as one of the primary receptors involved in the actions of HMGB1. Building upon this knowledge, we conducted further investigations into the mechanisms by which HMGB1 activates ESCs. Through experimental studies, we made a significant discovery: the stimulating effect of HMGB1 on ESCs exhibited a marked reduction when we selectively suppressed TLR4 receptors within these cells. This observation indicates that HMGB1 activates the migration and differentiation capabilities of ESCs by directly targeting the TLR4 receptors situated on the cell surface. Furthermore, the exposure of HMGB1 to ESCs positively regulated the expression of Notch1, Hes1, Wnt1, β-catenin, and c-myc, subsequently inducing the translocation of β-catenin to the cell nucleus. However, when TLR4 expression was suppressed in ESCs, this effect was no longer observed. Existing literature suggests a correlation between high TLR4 expression and the activation of the Wnt and Notch signaling pathways. 36,37 Studies have demonstrated the spatial co-localization of TLR4 and Dickkopf WNT signaling pathway inhibitor 3 (DKK-3), a crucial inhibitor of the Wnt pathway. DKK-3 exerts inhibitory effects on β-catenin by either suppressing LRP6 38 or forming a complex with β-TRCP, 39,40 thereby promoting intracellular degradation of β-catenin and negatively regulating the Wnt/β-catenin pathway. The absence of TLR4 can lead to increased DKK-3 expression, consequently inhibiting the response of the Wnt pathway. 41 However, research on how TLR4 activates the Notch/Hes signaling pathway remains limited, necessitating further comprehensive investigations.
Wnt and Notch pathways play important roles in regulating the proliferation and differentiation of ESCs and promoting wound-healing processes. The Notch pathway plays a complementary role by promoting the differentiation of stem cells and facilitating the efficient repair of wounds. 42 The Wnt/β-catenin pathway is also involved in maintaining the characteristics of ESCs and controlling their lineage differentiation. Activation of β-catenin signaling occurs upon skin injury, thereby altering the fate of ESC differentiation. 43 –45 Consequently, we postulate that following stimulation of TLR4 by HMGB1, it is plausible to assume that the activation of the Notch/Hes and Wnt/β-catenin pathways act as a mechanism to activate ESCs (Fig. 6B). However, there is still considerable debate as to whether the β-catenin/c-myc signaling pathway plays an active role in skin repair, especially for keratinocytes. 46 –51 This could be associated with the diverse post-translational modifications of the c-myc. 52 Besides, a recent single-cell sequencing study proposes that keratinocytes can modify, mediate and regulate the differentiation state of both their own and neighboring cells via the Wnt/β-catenin pathway in response to external stimuli. The induction of this effect is contact-dependent, implying that varying experimental outcomes might be linked to different conditions of intercellular contact. 53 Moreover, the differential interactions between the β-catenin and Notch pathways 53 –56 may also explain the divergent results seen in various studies, necessitating further research for clarification.
Through in vivo experiments, it has been observed that HMGB1 plays a significant role in promoting the process of wound healing. It contributes to epidermal proliferation and supports normal differentiation, thereby demonstrating its potential to enhance the overall quality of the healing process. In addition, HMGB1 may exert effects on other cells within the wound bed, including promoting fibroblast proliferation and affecting collagen synthesis, 57,58 stimulating endothelial cell proliferation, 10 and mediating wound immunity. 59 Excessive levels of HMGB1 may trigger an exaggerated inflammatory response, prolonging the inflammatory phase and resulting in the formation of hypertrophic scars. 57 NGS technology enables high-throughput capture of a wide range of genetic variations, gene expressions, and epigenetic modifications. Integrating NGS into future research can yield more comprehensive and unbiased assessments of the genome and transcriptome, aiding in a deeper understanding of the mechanisms mediated by HMGB1 and facilitating further exploration of the optimal concentration and timing for HMGB1 utilization.
CONCLUSIONS
Our study elucidated the role of HMGB1 in the process of wound healing, both in vivo and in vitro. Extracellular HMGB1 induced TLR4, activated the canonical Wnt and Notch signaling pathway, and increased the expression and nuclear translocation of β-catenin, ultimately enhancing the migration and differentiation of ESCs. In summary, our study sheds light on a novel potential approach and therapeutic target to increase wound healing.
KEY FINDINGS
HMGB1 was significantly elevated in normal acute wounds, while it is markedly reduced in chronic wounds.
In vitro, HMGB1 enhanced both the migration and differentiation of ESCs via the “HMGB1-TLR4-Wnt/Notch” axis.
In vivo, HMGB1 facilitated wound healing and epidermis differentiation via the “HMGB1-TLR4-Wnt/Notch” axis.
Footnotes
ACKNOWLEDGMENTS AND FUNDING SOURCES
This work was mainly supported by the
AUTHOR DISCLOSURE AND GHOSTWRITING
No competing financial interests exist. The content of this article was expressly written by the authors listed. No ghostwriters were used to write this article.
ABOUT THE AUTHORS
SUPPLEMENTARY MATERIAL
Supplementary Figure S1
Supplementary Figure S2
Supplementary Figure S3
Supplementary Figure S4
Supplementary Figure S5
Abbreviations and Acronyms
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.