
Abstract
Significance:
The incidence of diabetes continues to rise throughout the world in an alarming rate. Diabetic patients often develop diabetic foot ulcers (DFUs), many of which do not heal. Non-healing DFUs are a major cause of hospitalization, amputation, and increased morbidity. Understanding the underlying mechanisms of impaired healing in DFU is crucial for its management.
Recent Advances:
This review focuses on the recent advancements on macrophages and neutrophils in diabetic wounds and DFUs. In particular, we discuss diabetes-induced dysregulations and dysfunctions of macrophages and neutrophils.
Critical Issues:
It is well established that diabetic wounds are characterized by stalled inflammation that results in impaired healing. Recent findings in the field suggest that dysregulation of macrophages and neutrophils plays a critical role in impaired healing in DFUs. The delineation of mechanisms that restore macrophage and neutrophil function in diabetic wound healing is the focus of intense investigation.
Future Directions:
The breadth of recently generated knowledge on the activity of macrophages and neutrophils in diabetic wound healing is impressive. Experimental models have delineated pathways that hold promise for the treatment of diabetic wounds and DFUs. These pathways may be useful targets for further clinical investigation.
SCOPE AND SIGNIFICANCE
By 2045, the diabetic patient population is projected to reach 784 million worldwide (https://diabetesatlas.org/. Accessed April 2024). Diabetic foot ulcers (DFUs) are expected to develop in up to 34% of diabetic patients and up to 20% of these may require amputation. 1,2 Thus, it is crucial to understand the mechanisms that impair healing in DFUs to pave the way for the development of novel therapeutic strategies aimed at tackling this debilitating condition. This review focuses on the changes observed in macrophages and neutrophils in a diabetic environment, and how these affect wound healing.
TRANSLATIONAL RELEVANCE
Diabetes is characterized by hyperglycemia, insulin resistance, and stalled inflammation. Non-resolving inflammation is at least partly due to dysregulation and dysfunction of macrophages and neutrophils in diabetic wounds and DFUs. Delineation of deregulated cellular and molecular pathways in diabetic macrophages and neutrophils has the potential to identify novel therapeutic strategies to improve diabetic wound healing.
CLINICAL RELEVANCE
Macrophages and neutrophils are major players in the wound healing process and recent studies suggest that modulation of their activities may lead to improved outcomes in DFUs. In this review, we discuss recent discoveries in macrophage and neutrophil dysfunction in experimental models of diabetic wound healing and in studies of tissues isolated from diabetic patients with DFUs.
OVERVIEW
Diabetes is a complex metabolic disorder that is characterized by chronic hyperglycemia, insulin resistance, and dysregulation of the inflammatory environment. All these are associated with increased incidence of foot ulcers, many of which become chronic and pose a serious health risk. 3 The diabetic environment has profound effects on the function of macrophages and neutrophils. 4 In this review, we discuss how their functions are altered in diabetic wound healing.
DISCUSSION
The skin, the largest organ in our body, provides a protective barrier against pathogens, regulates body temperature, prevents water loss, and more. It consists of the epidermis, dermis, and subcutaneous tissue layers, all hosting various immune cells in steady state that are crucial for maintaining homeostasis.
Following an injury, the affected tissue initiates a cascade of events that are referred to as the phases of wound healing. The inflammatory phase is characterized by infiltration of the affected site by immune cells to protect against pathogens and clear debris. The proliferative phase aims to generate cells necessary for wound closure. The final phase, remodeling, consists of maturation and reorganization of the tissue. These phases overlap significantly, and are tightly regulated. 5
Macrophages and neutrophils are immune cell subsets that play crucial roles in the wound healing process and have been extensively studied. In diabetic wound healing, dysfunctional behavior of these cells impedes proper healing and elevates the risk for wound chronicity. Below we discuss the current knowledge on these cell subsets in skin homeostasis and in normal and diabetic wound healing, focusing on the inflammatory and proliferative phases. We also discuss how diabetes disrupts the functions of these cells, especially in the context of diabetic wounds and DFUs.
Macrophages
One of the major immune cell types of the skin in steady state are the tissue-resident macrophages. Skin-resident macrophages seed the dermis during fetal development. 6 In homeostasis, a small population of skin-resident macrophages replenishes via local progenitor cells throughout life, while the majority are continually replenished from circulating monocytes. 7 Skin macrophages actively survey their environment for signs of infection, damage, or tumor growth. They promote tissue function via close contact with local skin cells and elements, such as hair follicles and sebaceous glands, vasculature, and nerve endings. 8,9 Upon injury, both local skin macrophages and macrophages derived from peripheral monocytes infiltrate the wound where they modulate healing. 5
Macrophages in wound healing
Below, we discuss the functions of macrophages in normal healing and diabetic wounds. We use nomenclature to depict inflammatory (M1) and non-inflammatory/reparative (M2) macrophages. 5 The initial dichotomy of these broad macrophage functions stems from the differential ability of these cell subsets to metabolize arginine, with M1 using inducible nitric oxide synthase (iNOS) to generate nitric oxide (NO) to promote inflammation, and M2 using arginase 1 (Arg1) to generate ornithine and urea to promote cell proliferation and tissue repair. M2 macrophages are further subdivided into subsets with distinct functions in the wound healing process, with M2a supporting cell proliferation and extracellular matrix generation, M2b regulating inflammation, M2c important for matrix remodeling and angiogenesis (the formation of new blood vessels from existing ones), and M2d exerting angiogenic and remodeling activities. 10,11 These subsets have been characterized under specific macrophage polarization conditions in vitro. It is important to underline that the polarization of macrophages within a wound is not a simple process. During healing, macrophages are exposed simultaneously to various cues, leading to co-expression of multiple markers associated with several subsets. 12,13
To phenotype wound macrophages, the markers lymphocyte antigen 6 complex (Ly6C), an extracellular antigen, and C-X3-C motif chemokine receptor 1 (CX3CR1) are commonly used. 14 These markers are analogous to CD14hiCD16lo/- and CD14loCD16hi monocytes in humans. 15 Macrophages with high levels of Ly6C and low levels of CX3CR1 (Ly6ChiCX3CR1lo) typically exhibit inflammatory properties, whereas those with low levels of Ly6C and high of levels of CX3CR1 (Ly6CloCX3CR1hi) are mostly anti-inflammatory and reparative. 14 However, recent transcriptomic studies reveal that wound macrophages exhibit significant plasticity, preforming diverse functions and simultaneously expressing multiple markers during the healing process. 12
To address the evolving changes in macrophage polarization, a recent study identified four conserved pathways of macrophage activation: a phagocytic/regulatory path, an inflammatory/cytokine producing path, an oxidative stress/antimicrobial path, and a remodeling path. 16 Although this categorization is not yet widely applied, it is valuable to consider when interpreting data on macrophage activation/polarization.
Macrophage functions in normal acute wounds promote healing
Macrophages are prominent cells at the wound site and play key roles in all stages of healing. 17 They consist of skin-resident macrophages and infiltrating macrophages derived from circulating monocytes. There are a few studies that assess the contribution of infiltrating and activated skin-resident macrophages, 16,18 but to date their distinct contributions remain unclear. Below we briefly discuss the contribution of macrophages in each healing phase of normal healing wounds, highlighting recent advances.
Inflammatory phase
Monocytes that enter the wound early after injury express the chemokine receptor 2 (CCR2), respond to cc chemokine ligand 2 (CCL2, the ligand for CCR2), and both express and respond to CCL7, promoting monocyte/macrophage recruitment to the wound. 12,14,19,20 These monocytes mature into macrophages at the wound site. This first wave of macrophages in the wound is mostly inflammatory (M1) in nature, expressing high levels Ly6C and low levels of CX3CR1. 14 These macrophages also express iNOS, interleukin 1-beta (IL-1β), and tumor necrosis factor-alpha (TNF-α), all of which promote inflammation. The active form of IL-1β, a cardinal cytokine of inflammation, is produced via the activity of NOD-like receptor pyrin domain containing 3 (NLRP3) inflammasome. The NLRP3 inflammasome converts procaspase 1 to its active form caspase 1. This activated caspase 1 then facilitates the conversion of pro-IL-1β into its mature and active form, IL-1β. Overproduction of IL-1β is detrimental to wound healing, and thus, its production is tightly regulated. Peroxisome proliferator-activated receptor gamma (PPAR-γ) inhibits NLRP3 inflammasome activation, thereby regulating the production of IL-1β (Fig. 1). 21,22

Macrophages play a pivotal role in promoting healing in normal-healing wounds. Inflammatory macrophages are recruited via CCL2 and CCL7 expressed by tissue-resident cells and by infiltrating cells themselves early during the healing process (inflammatory phase). Inflammatory macrophages express high levels of Ly6C (Ly6Chi), CCR2, iNOS, IL-1β, TNF-α, and low levels of CX3CR1 (CX3CR1lo). Macrophage NLRP3 is activated in response to various stimuli and cleaves procaspase1 to generate active caspase 1, which in turn converts pro-IL-1β to IL-1β. PPAR-γ regulates excessive inflammation by controlling NLRP3 activity. During the proliferative phase, macrophages shift to a reparative phenotype. Reparative macrophages are associated with no or low expression of Ly6C (Ly6Clo) and high expression of CX3CR1 (CX3CR1hi). In addition, these macrophages express Arg1, CD206, and CD163 expression, and promote phagocytosis and efferocytosis. Phagocytosis is also facilitated by C1q. VEGFR1+CD206+ and IL-4R+ macrophages promote angiogenesis. CD301b+CD206+ macrophages are important for myofibroblast proliferation and heterogeneity during the proliferative phase of wound healing. Black arrows denote positive regulation; blunted lines denote inhibitory effect; dotted arrows indicate increases (up) or decreases (down). Arg1, arginase 1; C1q: complement component 1q; CASP1, caspase 1; CCL: cc chemokine ligand; CCR, CC motif chemokine receptor; CX3CR1, C-X3-C motif chemokine receptor 1, IGF1, insulin growth factor 1; iNOS, inducible nitric oxide synthase, Ly6C, lymphocyte antigen 6 complex; NLRP3, NOD-like receptor pyrin domain containing 3; IL-1β, interleukin 1 beta; PDGF, platelet-derived growth factor; PPAR-γ: peroxisome proliferator-activated receptor gamma; SASP, senescence-associated secretory phenotype; TNFα, tumor necrosis factor-alpha; VEGF, vascular endothelial growth factor; VEGFR1, vascular endothelial growth factor 1.
Proliferative phase
The proliferative phase begins shortly after the initiation of the inflammatory phase. This phase aims to generate new cells and reconstruct the tissue. Macrophages transition to a reparative state; they secrete growth factors (e.g., vascular endothelial growth factor, VEGF; insulin-like growth factor, IGF-1; and platelet-derived growth factor) to support local cell proliferation, and promote the formation of granulation tissue, a preliminary vascularized extracellular matrix to allow keratinocyte migration and wound closure. 17
Reparative macrophages usually express Arg1 and high levels of CX3CR1. 12,19 Although CCR2+ macrophages (inflammatory) accumulate early during wound healing, CCR2 deletion impairs angiogenesis, suggesting that these macrophages promote the proliferative phase either directly or indirectly. 20 During this phase, macrophages also express a variety of scavenger receptors such as CD206 (C-type mannose receptor), CD163, and CD36, involved in phagocytosis (the process of engulfing and ingesting particles or tissue fragments that are bigger than 0.5 µm) and linked with anti-inflammatory and proliferative functions. 5 CD206 expressing macrophages are divided into multiple and overlapping subsets. CD206+ macrophages co-expressing CD301b are crucial for re-epithelialization, tissue proliferation, and fibroblast numbers, but do not affect angiogenesis. 23 Interestingly, CD301b+ macrophages express both inflammatory and non-inflammatory markers and regulate insulin sensitivity and glucose metabolism, in mice fed a high-fat diet. 24 CD206+ macrophage subsets co-expressing the VEGF receptor 1 (VEGFR1) promote angiogenesis, and downregulate the expression of inflammatory cytokines (Fig. 1). 25
IL-4, a central cytokine in wound healing, exerts anti-inflammatory and pro-reparative functions. 26,27 IL4-R expressing macrophages support angiogenesis in various wound healing models. 28,29 During the late stages of healing, macrophages express the complement component 1q (C1q), 12 which promotes phagocytosis and angiogenesis, 30 and to regulate NLRP3 inflammasome activity. 31 Furthermore, macrophages can convert to fibroblast-like cells, (referred to as fibrocytes), which facilitate healing. 32 This process is mediated via keratinocyte secretion of extracellular vesicles (EVs) that contain microRNA 21 (miR-21). 32 EVs are cell-derived membrane vesicles that carry bioactive molecules as a form of cell-to-cell communication.
A crucial function of macrophages in wound healing is efferocytosis, a process during which they remove apoptotic cells (particularly neutrophils) from the wound. This process is instrumental for the progression to the proliferative phase and for proper healing. 33 Interestingly, lymphatic vessel endothelial hyaluronan receptor 1 (Lyve1+) expressing macrophages were recently shown to mediate this process. 34 PPARγ activity is a key step in efferocytosis. Some of PPARγ targets include Arg1 and scavenger receptors (CD36, CD206) that aid in the recognition of apoptotic cells. 35 Macrophages are also important for the removal of senescent cells. Shortly after an injury, keratinocytes and fibroblasts enter cell cycle arrest and secrete what is known as the senescence-associated secretory phenotype (SASP). SASP results in the production of large amounts of inflammatory cytokines, as well as metalloproteinases, extracellular matrix proteins, and growth factors, thus promoting repair. The presence of senescent cells at the wound site is transient and are thought to be removed by macrophages (Fig. 1). 36 Inability to remove these senescent cells from the wound contributes to impaired healing. 36
Macrophage supplementation studies in normal healing wounds
Studies using macrophage supplementation approaches have proved to be somewhat conflicting. Wounds injected with bone marrow-derived macrophages (BMDMs) that were not polarized (M0), or polarized toward M2a (with IL-4) or M2c (with IL-10), did not promote normal acute wound healing. 37 However, CD301b+ macrophages isolated from day 5 wounds (proliferative phase) and injected into day 3 wounds (late-inflammatory phase) promoted normal wound healing, while this was not the case when total wound macrophages were used. 23 The different outcomes of the two studies suggest that in vitro generated M2 macrophages 37 do not recapitulate the healing abilities of reparative macrophages that are polarized in the wound environment. 23 However, M0 BMDMs seeded in the pullulan–collagen composite and delivered to splinted wounds also promoted wound healing. These macrophages polarized toward all subsets within the wounds. 38 This improvement in wound healing may be related to the delivery method and/or the wound healing model used (splinted wound).
Macrophage dysfunctions in diabetic wounds delay healing
Diabetic wounds, and notably DFUs, often exhibit impaired healing, which may lead to chronicity. Several animal models have been developed to study diabetic wound healing, recently reviewed by Singh et al. 39 Figure 2 depicts only the diabetic animal models that are heavily referenced in this review.

Diabetic mouse models referenced in this review.
Hyperglycemia, increased levels of advanced glycation end-products (AGEs), and persistent cellular senescence and SASPs in diabetic wounds have all been associated with elevated pro-inflammatory cytokines and oxidative stressors, 36,40,41 and deregulated macrophage subsets in diabetic wounds. 12,14,42
The wound healing phases of diabetic wounds are not as well defined as in normal healing wounds. 14,42 –44 Below we discuss how macrophage dysregulation and dysfunction adversely affect wound healing in diabetic wounds.
Inflammatory phase
Our understanding of macrophage functions during the early steps of diabetic wound healing stems mainly from studies in rodents. Time-course experiments revealed a delayed onset of the inflammatory phase in diabetic mouse wounds compared with those of normal mice. Several studies involving streptozotocin (STZ)-induced type 1 diabetic (T1D) and db/db type 2 diabetic (T2D) mice show that inflammatory macrophages appear late in diabetic wounds. 42,44 –46 Similarly, mice that were rendered diabetic due to high-fat diet also demonstrated delayed infiltration of their wounds by Ly6Chi macrophages and dampened inflammatory cytokine production. 14 Interestingly, all these studies indicate that during the later stages, diabetic wounds are persistently populated by monocytes/macrophages.
The delay in macrophage response early in a diabetic wound is attributed to elevated IL-10 expression, which reduces the production of proinflammatory cytokines at the affected site, 44 and possibly ischemia (reduced vascularization), which is a well-established co-morbidity in a diabetic wound. 47 The impact of ischemia on reduced macrophage responses in diabetic wounds during the inflammatory phase remains to be experimentally evaluated. Interestingly, topical treatments of diabetic wounds with CCL2, and antibodies against IL-10 or IL-10 receptor early after injury restored macrophage responses and promoted wound healing in diabetic wounds. 44 –46 These reports highlight the importance of circulating monocytes/macrophages in diabetic wound healing processes. They further suggest an impairment in the ability of diabetic tissue-resident cells to mount an adequate response to injury (Fig. 3).
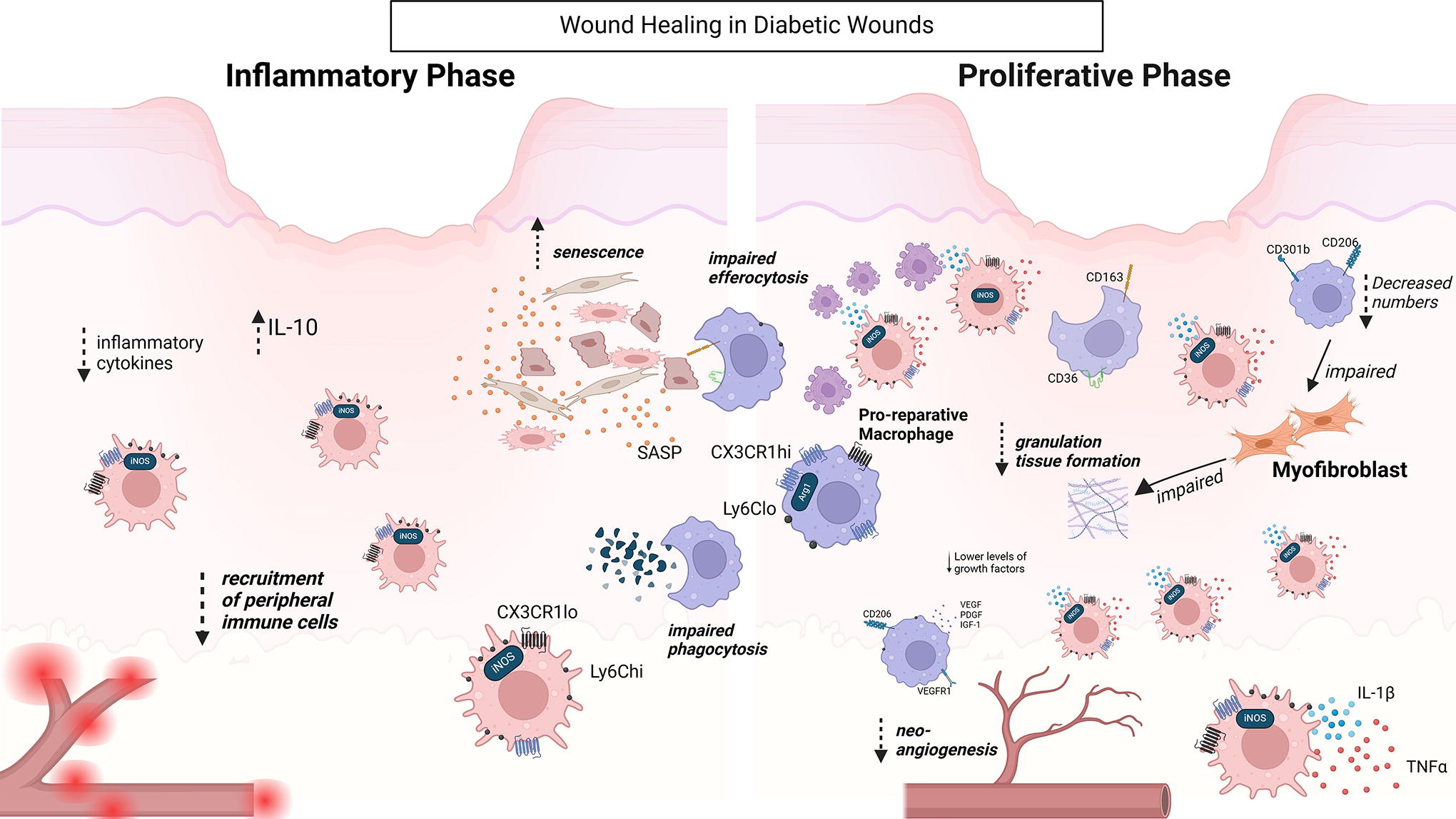
Macrophages are dysregulated in the diabetic environment, impairing wound healing. Diabetic wounds exhibit delayed inflammatory macrophage infiltration. During the later stages, both inflammatory and reparative macrophages are present but with altered functionalities leading to non-resolving inflammation. During this phase, VEGFR1 signaling is impaired in macrophages. In addition, macrophages express decreased levels of CD206 and increased levels of IL-1β. All these result in impaired angiogenesis. Decreased numbers of CD301b+CD206+ macrophages result in impaired myofibroblast proliferation and heterogeneity. Diabetic wounds have increased senescent cells, potentially due to impaired phagocytosis and efferocytosis of macrophages. Black arrows denote positive regulation; blunted lines denote inhibitory effect; dotted arrows indicate increases (up) or decreases (down) of cytokines/growth factors/gene expression/cell functions. Arg1, arginase 1; CX3CR1, C-X3-C motif chemokine receptor 1, IGF1, insulin growth factor 1; iNOS, inducible nitric oxide synthase; IL-1β, interleukin 1 beta; Ly6C, lymphocyte antigen 6 complex; SASP, senescence-associated secretory phenotype; VEGF, vascular endothelial growth factor; VEGFR1, vascular endothelial growth factor receptor 1.
In addition to impaired macrophage recruitment, the transcriptome of the subsets of diabetic macrophages differs from that of the non-diabetic counterparts, likely contributing to the inability of the diabetic wounds to heal in a timely manner. In particular, macrophages isolated from diabetic mouse wounds had decreased levels of pro-healing gene expression. 12,14,42
Proliferative phase
During the later stages of diabetic wound healing, there is a notable increase in inflammatory macrophages. Ly6Chi inflammatory macrophages persist and continue to increase over time, releasing inflammatory cytokines (Fig. 3). 14,42,48,49 Blocking these inflammatory macrophages during the later stages of healing promotes closure in diabetic wounds, 14 indicating their pivotal role in hindering diabetic wound healing.
Diabetic wound macrophages at this stage of healing increase NLRP3 inflammasome activity, resulting in increased bioactive IL-1β production. In a feedback loop, IL-1β stimulates NLRP3, thus further increasing IL-1β levels. 22 Furthermore, macrophages of diabetic mice and diabetic patients have decreased PPAR-γ, thus leading to decreased regulation of NLRP3. 21 Regulation of NLRP3 inflammasome is one of the main mechanisms by which topically applied ON101, an extract from medicinal plants, improves DFU wound healing. 50 ON101 facilitates the transition of proinflammatory macrophages toward reparative macrophages. 51
Reparative macrophage numbers are initially low in diabetic wounds, but eventually reach similar levels to those of control mice. These macrophages, however, have decreased expression of CD206 and produce lower levels of growth factors (such as transforming growth factor-β [TGF-β], IGF-1, and VEGF) at the wound site. 49 Wounds of STZ-induced diabetic mice have lower numbers of proangiogenic VEGFR1+ macrophages and inactivation of VEGFR1 signaling results in increased levels of IL-1β leading to impaired angiogenesis (Fig. 3). 25
In addition, CD301b+ macrophages, an instrumental subset for normal wound healing, are decreased in diabetic wounds. This results in decreased myofibroblast heterogeneity, and impairs myofibroblast function (Fig. 3). 52 Furthermore, the macrophage-to-fibrocyte conversion observed in the granulation tissue of healing wounds is impaired in db/db wounds due to decreased levels of miR-21 containing keratinocyte-derived EVs. This process was also shown to be impaired in non-healing DFUs: EVs isolated from wound fluids of diabetic patients with healing wounds had higher levels of miR-21 compared with those that did not heal. 32
Diabetic macrophages are characterized by impaired phagocytosis and efferocytosis, both crucial processes for a productive transition from the inflammatory to the proliferative phase. 40,53
Diabetic wounds are also associated with the increased presence of senescent cells, which may be associated with impaired phagocytosis and efferocytosis. 36,54,55 Furthermore, the SASP of senescent macrophages themselves contains high levels of CXCR2 (CXC motif cytokine receptor) ligands (CXCL1 and CXCL2; CXC motif cytokine ligand), which act on fibroblasts to promote a fibrotic phenotype and fibroblast senescence. 56
Macrophage supplementation studies in diabetic wound healing
The importance of macrophages in diabetic wound healing is also highlighted in macrophage supplementation studies. Exogenous delivery via alginate dressings of M0 macrophages, or macrophages polarized toward M1 (using lipopolysaccharide (LPS)/interferon (IFN)-γ), M2a (using IL-4 and IL-13), or M2c (using IL-10) all promoted wound healing. 13 Interestingly, all of these in vitro polarized cells eventually repolarized once in the wound and expressed markers consistent with the M2c subset. Furthermore, M2c conditioned media were sufficient to recapitulate the effect. 13 A different study showed that delivery of M0 BMDM from wild-type mice improved healing in wounds of diabetic mice. This suggests that simply increasing the numbers of normal functioning macrophages in diabetic wounds is sufficient to promote healing. 38 A recent study took a different approach, using macrophages that were “educated” with the conditioned medium of tumor-associated macrophages (TAMEMs). TAMEMs promoted healing in diabetic wounds more effectively than any other macrophage subset used (M0, M1, or M2). TAMEMs had both pro-healing and anti-inflammatory properties. The authors defined a group of cytokines and growth factors secreted by TAMEMs, which could recapitulate the effect. 57
Effects of diabetes on macrophage functions
In this section, we discuss the effect of diabetes on metabolism, myelopoiesis, and epigenetic regulation of macrophages.
Changes in metabolic profiles of diabetic macrophages
It is now well established that macrophage functions are strongly associated with distinct metabolic profiles (reviewed by Eming SA et al. 58 ). In steady state, tissue-resident macrophages are metabolically characterized by oxidative phosphorylation. Upon wounding, infiltrating monocyte-derived macrophages and activated tissue macrophages use glycolysis for their energetic needs, resulting in the production of pyruvate and lactate. 58 Increased lactate production is associated with inflammatory macrophages; however, lactate also signals to neighboring macrophages to transition toward the reparative phenotype (Fig. 4). Glycolysis fuels the pentose phosphate pathway, which leads to the generation of nicotinamide adenine dinucleotide phosphate (NADPH) and reactive oxygen species (ROS) production, but also promotes the generation of precursor nucleotides and amino acids to support local proliferation. 59 During the inflammatory phase, the Krebs cycle in macrophages is fragmented leading to dampened respiration, and the production of succinate, 58 which promotes local inflammation, as well as itaconate, which dampens local inflammation via inhibition of NLRP3 inflammasome. 60 Itaconate also induces nuclear transcription factor-erythroid 2-related factor 2 (NRF2), which promotes antioxidant responses, and triggers the activation of the activating transcription factor 3, which regulates inflammatory gene expression. 61 Thus, changes in macrophage metabolism early in wound healing support the local proinflammatory environment, while in parallel set the stage for reparative functions. After the inflammatory phase, reparative macrophages gradually revert to oxidative phosphorylation, a normal Krebs cycle, and fatty acid oxidation for their energetic needs (Fig. 4). 58
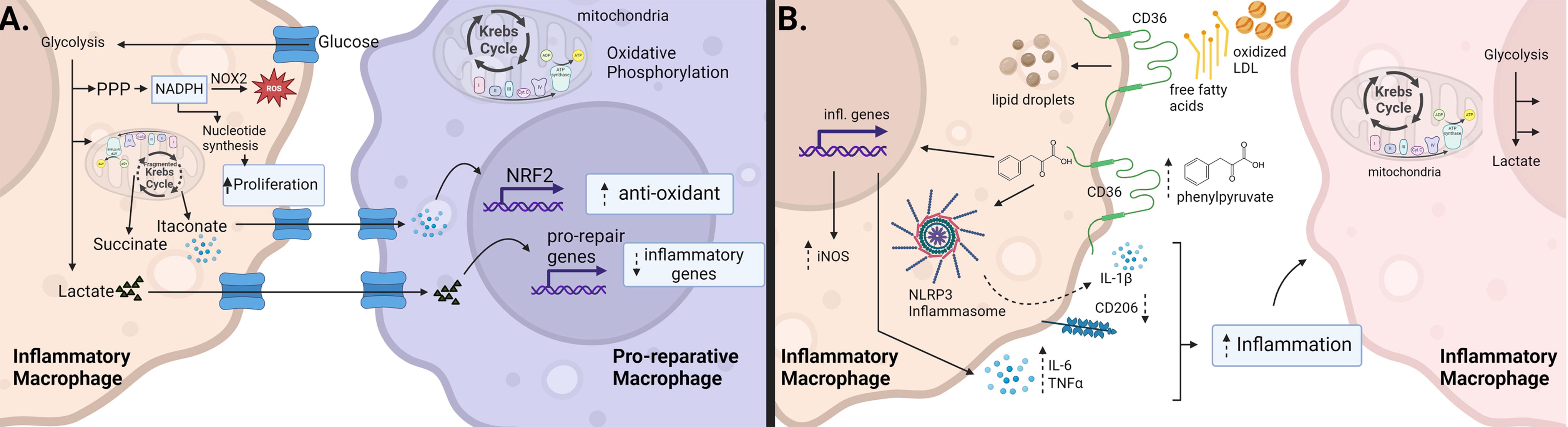
Diabetes alters the metabolic profile of macrophages. Inflammatory macrophages utilize glycolysis to produce lactate. Glycolysis also promotes the pentose phosphate pathway, which generates NADPH. NADPH can be used by NOX to produce reactive oxygen species but also generates precursors for nucleotides and amino acids thus promoting proliferation. In addition, the Krebs cycle is fragmented resulting in the production of succinate (inflammatory mediator) and itaconate (anti-inflammatory mediator). Itaconate produced by inflammatory macrophages acts on neighboring macrophages to promote an anti-inflammatory/reparative phenotype. Lactate can also be utilized to regulate inflammatory gene expression in neighboring macrophages. Reparative and tissue-resident macrophages utilize oxidative phosphorylation for their energy needs. In high-fat environments (as is often the case in diabetes), macrophages use CD36 to sequester free fatty acids and oxidized LDL, which they store in lipid droplets. CD36 also allows the entry of phenylpyruvate into the cell, which promotes inflammatory gene expression and NLRP3 inflammasome activation. Later in the wound healing process, inflammatory macrophages persist; however, they utilize both increased glycolysis and increased oxidative phosphorylation. Black arrows denote positive regulation; dotted arrows indicate increases (up) or decreases (down) of cytokines/growth factors/cell functions, or indirect effects (curved). CD: cluster of differentiation; IL, interleukin; iNOS, inducible nitric oxide synthase; NOX, NAPDH oxidase; NLRP3, NOD-like receptor pyrin domain containing 3; NRF2, nuclear transcription factor-erythroid 2-related factor 2; PPP, pentose phosphate pathway; TNF-α, tumor necrosis factor-alpha.
Metabolic disorders (hyperglycemia, obesity, insulin resistance, all diabetes hallmarks) promote an activation profile that is distinct from traditional inflammatory pathways. These macrophages increase the free fatty acid receptor CD36, which contributes to phagocytosis/efferocytosis in normal healing wounds but is often associated with inflammatory macrophages under diabetic conditions. 62 In the presence of increased fat levels (as is often the case in diabetic conditions), upregulation of CD36 in macrophages leads to increased uptake of fatty acids and oxidized lipoproteins, which can be stored in lipid droplets. 63 Furthermore, metabolomic analysis of chronic DFUs showed increased levels of phenylpyruvate, a phenylalanine metabolite. When tested in animal studies, phenylpyruvate was shown to delay excision wound healing. Phenylpyruvate enters macrophages via CD36 and promotes proinflammatory macrophages by increasing NLRP3 stability. This was accompanied by increased levels of Il-1β and other inflammatory cytokines (Fig. 4). 64
The effect of high glucose on macrophage function is debated. Short-term exposure to high glucose causes rapid reprogramming of monocyte/macrophages, manifested by increased IL-10 expression and secretion. IL-10 reduces glucose uptake in an autocrine/paracrine manner and transforms monocytes into an anti-inflammatory phenotype by dampening signaling through toll-like receptors. 44 Reduction in glucose uptake and macrophage transformation into an anti-inflammatory phenotype, due to short-term exposure to hyperglycemia, was corroborated in the STZ-induced animal model. 65 In contrast, prolonged exposure to glucose shifts macrophages into a proinflammatory state. 44 However, the observed shift toward glycolysis in diabetic conditions may be independent, or due to indirect effects, of increased glucose uptake and/or specific glucose metabolites. 65 Furthermore, insulin resistance was shown to promote glycolysis, and a shift toward a M2-like polarization in high-fat diet mice. 66
Additional dysregulations of macrophage metabolic profiles have been documented in patients with diabetes such as lower plasma concentrations of maresin 1, a docosahexaenoic acid pro-resolution metabolite produced by macrophages. 67
Efferocytosis, the clearance of apoptotic cells, initially occurs during the inflammatory phase when macrophages are highly glycolytic. 58 Efficient efferocytosis is associated with upregulation of the glucose transporter 1, and transiently increases glycolysis. 68 This occurs via 6-phosphofructo-2-kinase/fructose-2,6-bisphosphatase 2, a distinct glycolytic pathway from the one associated with inflammatory macrophages. 68 Efferocytosis-induced glycolysis elevates lactate levels, which induces neighboring macrophages to express pro-reparative genes. 58,68 This partly explains why efferocytosis facilitates the transition toward the proliferative phase. During the proliferative phase, macrophages shift their metabolism toward oxidative phosphorylation, which enhances efferocytosis. 58 Reparative macrophages express Arg1 (to metabolize arginine for the generation of polyamines, which promote cell proliferation), and L-proline (a crucial amino acid for collagen synthesis). Interestingly, efferocytotic cells can acquire arginine (and ornithine) from apoptotic cells. 69
Diabetic macrophages exhibit impaired efferocytosis, but the underlying mechanism(s) are not well delineated. However, increased expression of enzymes that degrade receptors that mediate efferocytosis and reduced PPARγ activity have been both associated with impaired efferocytosis in diabetes. 70
Effect of diabetes on myelopoiesis
Diabetic patients have increased circulating monocyte populations. 4 Similarly, diabetic mice show increased proliferation of hematopoietic stem progenitor cells and myelopoiesis resulting in more inflammatory monocytes into the circulation. 71 These monocytes have been associated with impaired wound healing in diabetic mice. 72 The release of increased numbers of myeloid cells is associated with decreased CXCL12 levels in the bone marrow. 71 CXCL12 is a retention signal for CXCR4 expressing quiescent hematopoietic stem cells, and other progenitor cells. Interestingly, a counter-regulatory pathway was identified in which diabetic mice upregulate the endothelial epidermal growth factor receptor (EGFR) signaling to dampen excessive myelopoiesis; this was also observed in diabetic patients marked by increased EGFR phosphorylation and lower transcripts of CXCL12 in endothelial cells (Fig. 5). The two pathways (i.e., CXCL12 downregulation and EGFR signaling) appear to act independently. A different study showed that the differentiation of diabetic hematopoietic stem cells toward monocytes is impaired due to increased oxidative stress and upregulation of the DNA methyltransferase 1; however, the polarization of macrophages was skewed toward an inflammatory M1 phenotype. 42
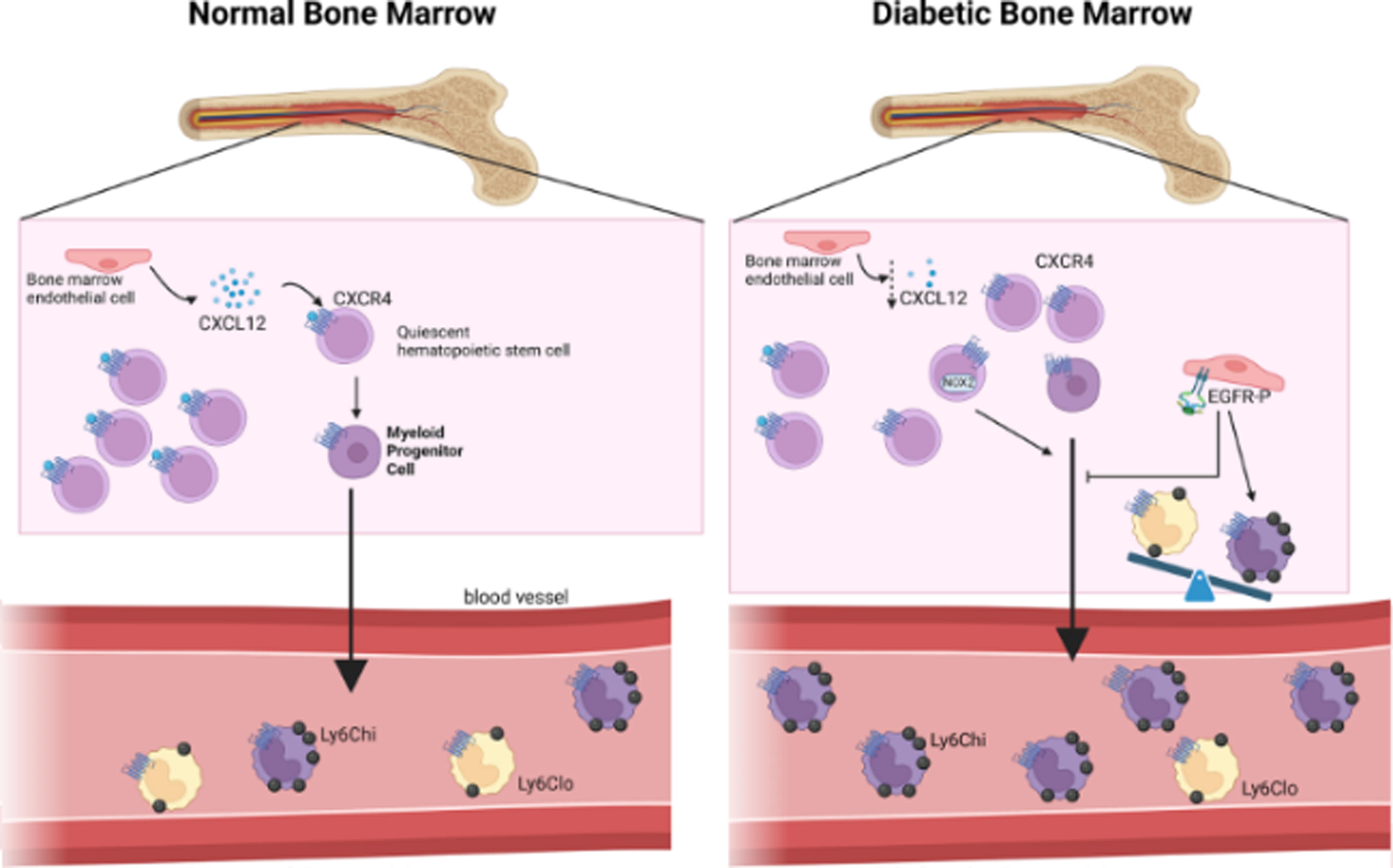
The impact of diabetes on myelopoiesis. Bone marrow endothelial cells (and bone marrow stromal cells, not shown) produce CXCL12, a signal that maintains hematopoietic stem cells (and other progenitor cells, not shown) in quiescence and acts as a retention signal via binding to CXCR4. In diabetic conditions, the production of CXCL12 in the bone marrow is reduced, resulting in increased monocyte release in the circulation. These monocytes are mostly inflammatory. Diabetes also promotes a counter-regulatory pathway in which bone marrow endothelial cells have increased phosphorylated EGFR, which results in dampened myelopoiesis. Black arrows denote positive regulation; blunted lines denote inhibitory effect; dotted arrows indicate increases (up) or decreases (down) of cytokines/growth factors/cell functions. CXCL, CXC motif cytokine ligand; CXCR, CXC motif cytokine receptor; EGFR, endothelial epidermal growth factor receptor; Ly6C, lymphocyte antigen 6 complex.
Epigenetic changes in diabetic macrophages
Numerous epigenetic modifications associated with diabetes promote inflammatory macrophage polarization. 73 Prolonged hyperglycemia induces permanent functional changes in macrophages similar to “trained immunity,” documented in innate immune cells acquiring immunological memory after exposure to specific stimuli. 74
BMDMs isolated from STZ-induced diabetic mice exhibit altered chromatin accessibility and heightened inflammatory responses, as well as impaired M2 polarization. 75 Transfer of diabetic, but not of normoglycemic, BMDMs to mice prone to atherosclerosis (mice deficient in low-density lipoprotein receptor; Ldlr−/− mice) induces atherogenic plaque formation, indicating inherent aberrant activation of these cells. The pattern of open chromatin suggested that these changes are driven by the transcription factor Runx1 (runt-related transcription factor 1). 75 Leukocytes and macrophages from atherosclerotic plaques of diabetic patients also showed enrichment of Runx1 target products. 75
Macrophages isolated from the wounds of diabetic patients exhibit reduced trimethylated histone H3 lysine 9 (H3K9me3), driven by decreases in methyltransferase Setdb2 (SET domain bifurcated histone lysine methyltransferase 2). This inhibits the recruitment of nuclear factor kappa-light-chain-enhancer of activated B cells (NF-κB), resulting in decreased levels of inflammatory cytokines. Setdb2 expression is induced by increased levels of IFN-β early after injury in normal wounds. In diabetic wounds, IFN-β levels remain persistently low, suppressing Setdb2 expression, thereby allowing inflammatory cytokine expression (Fig. 6). 76
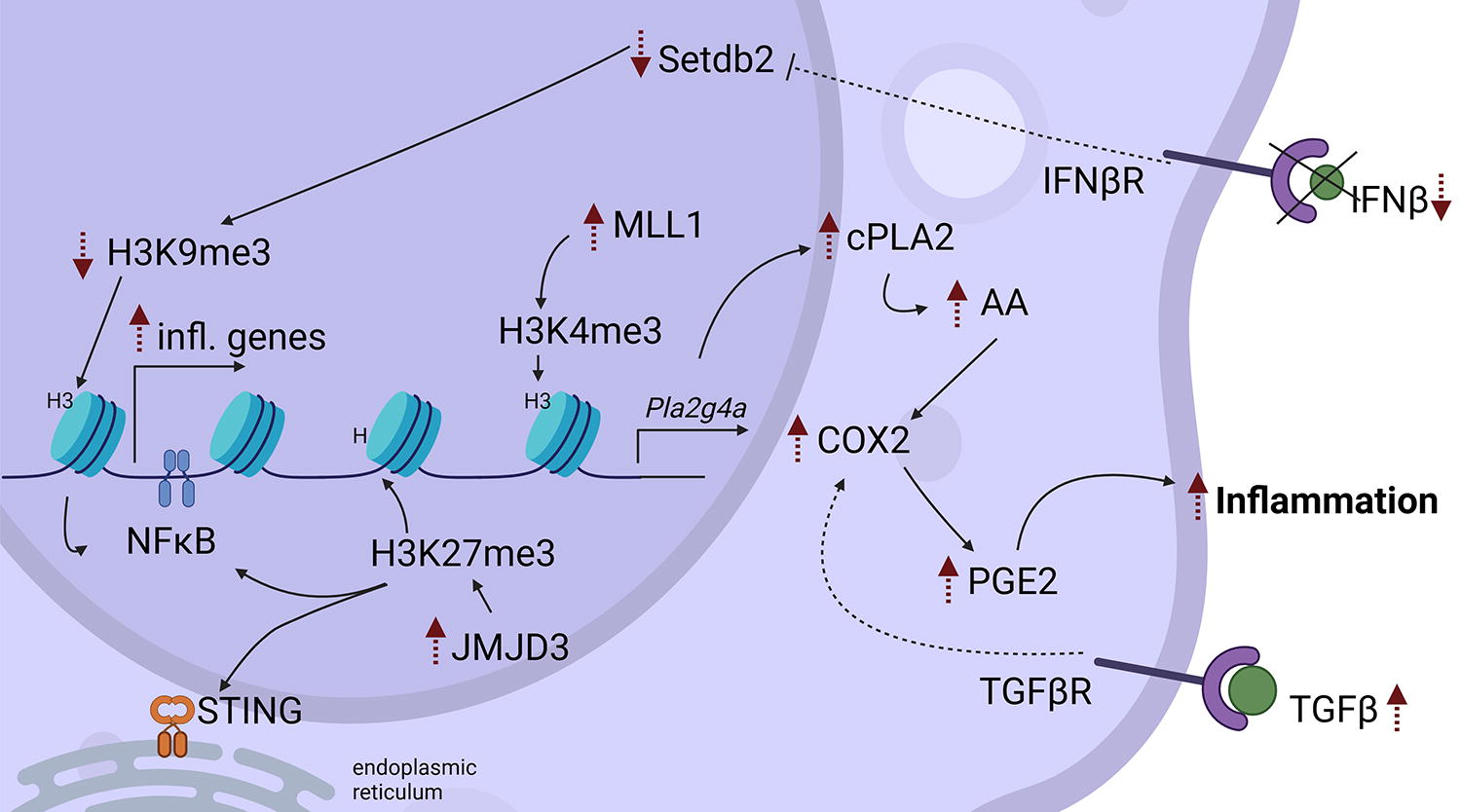
Diabetes induces epigenetic modifications in macrophages. Diabetic wounds have low levels of IFNβ, thus reducing the levels of Setdb. This leads to decreased H3K9me3 levels resulting in increased NFkB-mediated expression of inflammatory cytokines. MLL1 is upregulated in diabetic wounds, and this results in increased H3K4me3 levels, which promotes cPLA2 expression. cPLA2 releases AA, which is metabolized by COX2 into PGE2, a proinflammatory lipid mediator. JMJD3 is also increased in diabetic macrophages resulting in demethylation of H3K27me3, which results in NFkB- and STING-mediated inflammatory gene expression. Black arrows denote positive 1 regulation; dotted arrows indicate increases (up) or decreases (down) of cytokines/growth factors/gene expression/cell functions, or indirect effects (curved). COX-2, cyclooxygenase- 2; cPLA2, cytosolic phospholipase A2; H3K, histone 3 lysine; IFN, interferon; JMJD3, Jumonji domain-containing protein D3; Me3, trimethylation; MLL1, mixed lineage leukemia; NF-κB, nuclear factor kappa-light-chain-enhancer of activated B cells; PGE2, prostaglandin E2; Pla2g4a: phospholipase A2 group IVA (the gene encoding for cPLA2); SASP, senescence-associated secretory phenotype; Setdb2, SET domain bifurcated histone lysine methyltransferase 2; STING, stimulator of interferon genes; TGF-β, transforming growth factor-beta.
Macrophages from diabetic patients and diabetic mice display elevated expression of MLL1 (histone methyltransferase mixed lineage leukemia −1). MLL1 increases H3K4me3, which promotes TNF-α and IL-1β expression, thus amplifying inflammatory cytokine production. 77 In addition, MLL1 upregulates cPLA2 (cytosolic phospholipase A2), an enzyme that releases arachidonic acid (AA) from cell and nuclear membranes. In parallel, TGF-β, which is highly expressed in diabetic wounds, supports the expression of cyclooxygenase- 2 (COX-2). 78 COX-2 catalyzes the conversion of AA to intermediates culminating in the production of prostaglandin E2 (PGE2), which promotes inflammation and impairs phagocytosis by macrophages (Fig. 6). Pharmacological inhibition of COX-2 improved diabetic wound healing and mitigates inflammatory responses. 78
Demethylation of H3K27me3 by the histone demethylase Jumonji domain-containing protein D3 (JMJD3) also promotes inflammatory macrophages in diabetes. 48,79 Low levels of H3K27me3 induce STING-(stimulator of interferon genes) and NF-κB-dependent macrophage inflammatory responses. While this response promotes normal wound healing in the early stages, persistent JMJD3 upregulation in diabetic wound macrophages results in elevated TNF-α and IL-1β production during the late stages of healing (Fig. 6). 79 Adoptive transfer of diabetic macrophages impairs wound healing in control mice. 48 JMJD3 and STING are also elevated in diabetic patients’ wounds compared with non-diabetic controls. 79
Hyperglycemia induces upregulation of HDAC3 (histone deacetylase 3), promoting macrophage inflammation in STZ-treated diabetic mice. 80 Moreover, HDAC3 inhibition was shown to increase expression of CD206 in macrophages, promoted an anti-inflammatory wound environment, and improved angiogenesis. 80
Epigenetic modifications within the context of diabetes can also occur via long-non-coding RNAs (lncRNAs). For example, dynamin 3 opposite strand (Dmn3os) is an lncRNA induced via NF-κB activation in db/db mouse macrophages resulting in multiple histone modifications and amplified inflammation. 81 Hyperglycemia upregulates the lncRNA growth arrest-specific 5 (GAG5) in db/db wound fibroblasts and macrophages and impairs wound healing through the promotion of macrophage inflammation by upregulating iNOS, IL-1β, and TNF-α via the STAT1 (signal transducer and activator of transcription 1) signaling pathway. 82 Furthermore, NADPH oxidase 2 (NOX2)-dependent ROS production in macrophages was shown to be regulated by the lncRNA Lethe via NF-κB signaling, in which Lethe binds to the p65 subunit of NF-κB, preventing nuclear translocation to prevent NOX2 expression. 83 Diabetic macrophages increased NOX2 expression via Lethe, promoting ROS production and inflammation. 83
In addition to miR-21 discussed earlier, 32 other microRNAs (miRNAs) can also modulate diabetic macrophage function. EVs isolated from mice fed a low-fat diet (lean EVs) have been shown to contain miR-222-3p, a modulator of inflammation. 84 Lean EVs injected around db/db mouse wounds accelerated wound healing, promoted angiogenesis and CD206 expression, downregulated TNF-α and iNOS expression, and increased Arg1 expression in macrophages. Epidermal stem cell EVs decrease inflammation and promote anti-inflammatory macrophage polarization, as well as angiogenesis and cell proliferation and differentiation. 85 EVs released from proinflammatory macrophages contain miR-503, which impairs endothelial cell function, hindering wound healing. 86
Neutrophils
Neutrophils are the most extensively studied granulocytes. They are produced daily in the bone marrow at high rates, following well-defined steps. 87,88 A small fraction of neutrophils is released into the circulation, whereas the rest remain in reserve in the bone marrow via a CXCR4-mediated mechanism. 88
Neutrophils in wound healing
The main markers used for neutrophils are CD11b+Ly6G+ (lymphocyte antigen 6 ) in mice, and CD15+CD66b+CD16+CD14- in humans. These markers, however, also label polymorphonuclear myeloid-derived suppressor cells (MDSCs) but LOX1 (oxidized low-density receptor 1) and CD84 were recently identified as possible MDSC markers. 89 Neutrophils can also be categorized as N1, a subset that promotes inflammation and is defined by iNOS, IL-6, IL-1β, and TNF-α expression, and as N2, a subset that regulates inflammation and is defined by expression of CD206, Arg1, Ym1 (chitinase-like protein 3; Chil3), and IL-10. 90,91 Below we describe neutrophil functions in normal healing wounds and their malfunctions in diabetic wounds.
Neutrophils in normal wounds protect against infection and indirectly promote healing
Renowned as the “first line of defense,” neutrophils rapidly arrive at the wound site, where they initially contribute to inflammation, pathogen clearance, and ultimately resolution of inflammation. 92
Inflammatory phase
Initially, neutrophils migrate into the wound in response to danger-associated molecular patterns released during injury and pathogen-associated molecular patterns (PAMPs) of invading microorganisms. 93 This migration occurs in multiple waves orchestrated by ∼30 neutrophil receptors and multiple signaling pathways. 94,95 One of the most studied signals is the formyl peptide receptor FPR) recognition and response to N-formyl peptides (such as fMet-Leu-Phe; fMLP), which is released in the wound by both damaged cells and invading bacteria. 96 Local skin cells and neutrophils produce chemokines (e.g., CCL3, CXCL1, and CXCL2) that mediate subsequent neutrophil migration waves by engaging auxiliary chemokine receptors, such as CCR1, CXCR1, and CXCR2. 97,98 Neutrophils also produce chemotactic lipid mediators such as LTB4 (leukotriene B4), which signals via BLT1 (leukotriene B4 receptor 1) (Fig. 7). High levels of LTB4 at the wound site promote the migration of additional neutrophils from distal sites. 99
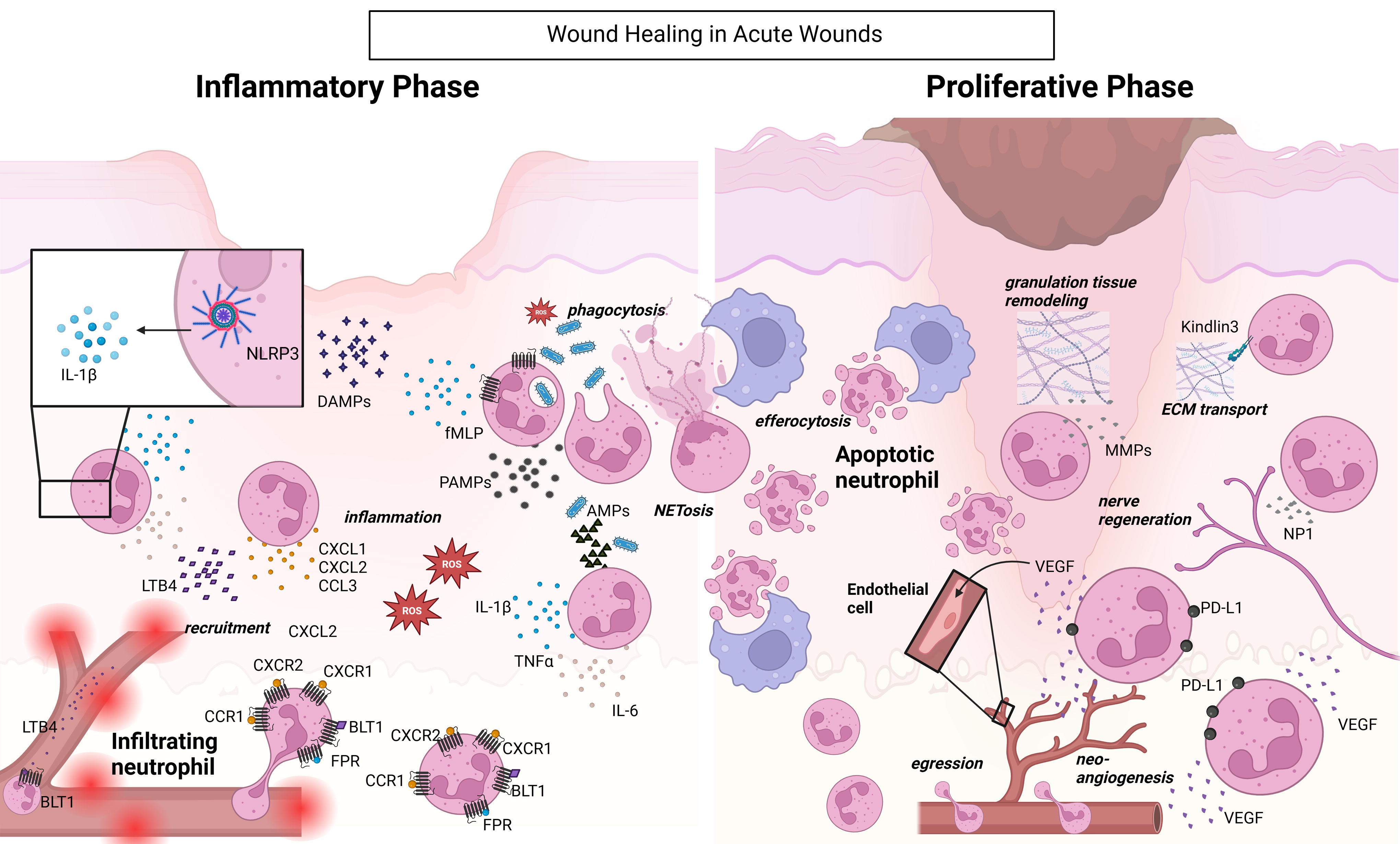
Neutrophil functions promote healing in normal-healing wounds. In normal-healing wounds, neutrophils are recruited via various receptors, including FPR, BLT1, CCR1, CXCR1, and CXCR2, in response to fMLP, LTB4, DAMPs, and PAMPs generated at the wound site. Within the wound, neutrophils themselves secrete chemoattractants such as CXCL1, CXCL2, and LTB4. NLRP3 activation in neutrophils results in the release of IL-1β. Neutrophils also produce inflammatory cytokines, such as TNFα and IL-6, to further amplify inflammation. In addition, they secrete AMPs and NETs to protect against invading pathogens. Apoptotic neutrophils are efferocytosed by macrophages, signaling the transition of macrophages toward a reparative phenotype, thus triggering the proliferative phase. Remaining neutrophils promote healing by transporting the ECM, promoting nerve regeneration and neoangiogenesis, during the proliferative phase. Black arrows denote positive regulation. AMPs, antimicrobial peptides; BLT1, leukotriene B4 receptor 1; CXCL, CXC motif cytokine ligand; CXCR, CXC motif cytokine receptor; DAMPs, danger-associated molecular patterns; ECM, extracellular matrix; fMLP: FPR, formyl peptide receptors; IL, Interleukin; LTB4, leukotriene b4; NETs, neutrophil extracellular traps; NP-1, neutrophil peptide 1; PAMPs, pathogen-associated molecular patterns; PD-L1, programmed cell death ligand 1; ROS, reactive oxygen species; TNF-α, tumor necrosis factor-alpha; VEGF, vascular endothelial growth factor.
Once in the wound, neutrophils exert multiple functions to protect the host from infection. They phagocytose pathogens and release a range of antimicrobial peptides (AMPs) and enzymes stored within their granules that promote inflammation and have direct bactericidal effects. 97,100 They destroy invading pathogens by producing copious amounts of ROS through NADPH and NOX, and myeloperoxidase, 101 and enhance local inflammation by producing proinflammatory cytokines, such as IL-6, IL-1β, and TNF-α. 101,102 In addition, neutrophils deploy extracellular traps (NETs), composed of cell-free DNA, citrullinated histones, neutrophil-derived proteins, and AMPs to immobilize and kill invading pathogens such as bacteria, viruses, parasites, or fungi (Fig. 7). 103,104
To avoid excessive inflammation, neutrophils are tightly regulated by various cells at the wound site such as keratinocytes, fibroblasts, or other immune cells. 105 Tissue-resident macrophages regulate neutrophil-induced inflammation by surrounding them, thus preventing chemoattractant signaling that promotes further neutrophil recruitment. 106 Neutrophils themselves can regulate their migration via desensitizing their G-protein-coupled receptors that respond to chemoattractants, thereby limiting their accumulation. 107 Elevated NET levels damage the surrounding tissue, thus contributing to local inflammation. The formation of NETs can be regulated by macrophage-mediated NET phagocytosis. 108 It has been documented that resolvins (inflammation pro-resolving lipid mediators) facilitate NET clearance. 109
The effect of neutrophil depletion in wound healing is debated. Depletion of neutrophils by anti-Gr1 antibody improved wound healing in uninfected mice, 110 yet delayed healing in aged mice. 111 A limitation of these studies may be that the anti-Gr-1 antibody recognizes both Ly6G and Ly6C, and the latter is also expressed in inflammatory monocytes/macrophages. 112 Depletion of neutrophils using the specific anti-Ly6G antibody (1A8) delayed healing in a burn injury wound model, suggesting an important role for neutrophils, at least in this model. 113
Proliferative phase
The proliferative phase of normal healing wounds is marked by dampened presence of neutrophils. The majority of neutrophils undergo apoptosis leading to their efferocytosis by macrophages, a process that acts as a signal for macrophages to switch to a reparative phenotype. 114 Multiple mechanisms have been associated with the recognition of apoptotic neutrophils by macrophages. 115 In addition, neutrophils may egress from the tissue to reenter the circulation (Fig. 7). 100 Interestingly, it was recently shown that mature and highly inflammatory neutrophils can produce high levels of itaconate, the anti-inflammatory metabolite produced via a fragmented Krebs cycle. Instead of releasing it to the extracellular space, neutrophils retain itaconate intracellularly. It is hypothesized that itaconate carrying neutrophils egress from the wound and relocate to the bone marrow, where they control myelopoiesis. 116
It is now well established that neutrophil subsets also exhibit pro-reparative functions, such as stimulation of angiogenesis and promoting tissue remodeling via extracellular matrix degradation. 93 A new neutrophil function in the presence of a respiratory infection was recently described during wound healing. In a mouse model of burn injury, peripheral neutrophils were shown to upregulate programmed cell death ligand 1 (PD-L1). 113 PD-L1 expressing neutrophils secrete high levels of VEGF to promote angiogenesis but exert limited bactericidal activities. In burn wound injury, PD-L1+ neutrophils accumulate in the lung (via PD-L1 binding to PD1 [programmed cell death protein 1] expressed by lung endothelial cells), delaying their migration to the wound. 113 This temporary retention in the lung ensures timely arrival of these proangiogenic neutrophils at the wound, aiding the tissue to heal properly. Blocking PD-L1+ neutrophil accumulation in the lung by the anti-PDL1 antibody reduces the risk of lung infection, but adversely impairs wound healing by promoting premature angiogenesis, due to excessive VEGF production and enhanced epidermal growth. 113
Other pro-reparative functions of neutrophils include nerve regeneration by producing neutrophil peptide 1, 117 and transporting extracellular matrix from healthy areas to the wound site to promote formation of granulation tissue (Fig. 7). 118
Neutrophil dysfunctions delay healing in diabetic wounds
Multiple dysfunctions have been associated with neutrophils in diabetic wounds. These abnormalities impair their ability to combat infections and later to promote tissue repair. This results in exacerbation of the healing process leading to severe complications in diabetic wounds.
Inflammatory phase
Similarly to macrophages, neutrophil infiltration is delayed, and their functionality is impaired in diabetic wounds. Multiple studies have documented impaired neutrophil migration to diabetic wounds. Neutrophil chemotaxis via FPR, the primary chemotactic receptor that initiates the first wave of neutrophil responses in wound and toward infection, 94,119 is impaired in mouse and human neutrophils in hyperglycemic conditions and in murine diabetic wounds. 120 This defect was shown to be overcome by topical application of CCL3 (a proinflammatory cytokine), which increased neutrophil trafficking by engaging the auxiliary receptor CCR1. This resulted in a significant reduction in infection burden and improved healing in diabetic wounds. 120 One of the reasons for this delayed neutrophilic migration to the site of infected diabetic wounds has been linked to the deficiencies in the bactericidal activity of diabetic neutrophils. Due to dampened bactericidal neutrophilic activity, Pseudomonas aeruginosa-infected diabetic wounds had decreased amounts of bioavailable PAMPs compared with non-diabetic wounds. 121 This decrease in PAMPs dampened toll-like receptor signaling in db/db murine diabetic wounds and slowed further neutrophil accumulation. Topical application of LPS promoted neutrophil trafficking and activity at the wound site and diminished wound infection and promoted healing in this diabetic animal model of infection (Fig. 8). 121

Dyreagulation of neutrophils in diabetic wounds results in impaired healing. Neutrophil recruitment is delayed in diabetic wounds. Phagocytosis of pathogens by neutrophils is impaired, resulting in less bioavailable PAMP generation. Neutrophils enter the wound later in the healing process and persist in the proliferative phase. During this phase, they continue to promote inflammation and release excess NETs; however, these NETs do not function properly. These impairments in neutrophil function prevent resolution of inflammation and wound healing. Black arrows denote positive regulation; dotted arrows indicate increases (up) or decreases (down) of cytokines/growth factors/gene expression/cell functions. AMPs, antimicrobial peptides; CXCL, CXC motif cytokine ligand; DAMPs, danger-associated molecular patterns; IL, interleukin; LTB4, leukotriene b4; NETs, neutrophil extracellular traps; PAMPS, pathogen-associated molecular patterns; ROS, reactive oxygen species.
Proliferative phase
Neutrophils accumulate and persist in the wound during the later stages of diabetic wound healing. 53,120,121
They exhibit increased NETosis in the chronic phase of diabetic wounds and this is associated with poor healing in DFUs and in diabetic mouse wounds (Fig. 8). 122 The increase in NET presence in diabetic wounds correlates with NLRP3 inflammasome activity 123 and sustained IL-1β production, contributing to inflammation. 124 NLRP3-mediated NETosis can be regulated by MFG-E8 milk fat globule epidermal growth factor VIII (MFG-E8). 125 Despite increased MFG-E8 levels in diabetes, it fails to control NETosis due to glycation-related modifications. Treating MFG-E8-/- diabetic mice with exogenous MFG-E8 controls NET formation and inflammasome activation, accelerating healing. 125 Furthermore, diabetic neutrophils exhibit impairments in ROS upregulation at infected wound sites. Normal neutrophils increase ROS production to meet demand upon infection. Diabetic neutrophils produce ROS in steady state, but struggle to further increase ROS production at the site of infection. This leads to impaired bactericidal activity. 53,126,127
Neutrophil depletion 110 and alleviation of neutrophil infiltration by a serotonin reuptake inhibitor or a beta-adrenergic blocker accelerate healing in diabetic mouse wounds. 128,129 These reports highlight that insufficient neutrophil responses early following an injury render diabetic wounds susceptible to infection. Conversely, exuberant and prolonged neutrophil activity during the later stages of healing contributes significantly to tissue damage and impairs the proper healing process.
Effects of diabetes on neutrophil function
The hallmarks of diabetes, hyperglycemia, insulin resistance, and AGEs, all impair neutrophil function. 127,130 Below we discuss metabolic and epigenetic changes observed in neutrophils in diabetic conditions.
Changes in metabolic profiles of diabetic neutrophils
Neutrophils are metabolically active cells and rely on glycolysis and respiratory burst in steady state. During their effector responses, they use gluconeogenesis and glycogenesis and increase their pentose phosphate pathway. 131 The pentose phosphate pathway produces large quantities of NADPH and promotes ROS production. In addition, mature neutrophils can also utilize glutaminolysis in the mitochondria for the generation of NADPH and ROS. 132 Unexpectedly, neutrophils do not rely on glycolysis and the pentose phosphate pathway for their energetic needs under diabetic and hyperglycemic conditions. Instead, they use the polyol pathway during which glucose is metabolized to sorbitol and oxidizes NADPH to NADP+. 133 This pathway leads to the accumulation of sorbitol and is associated with NET formation and increased ROS generation in steady-state circulating neutrophils. 133 This constitutive NET formation in T2D patients is linked to impaired responses to infection. The metabolic shift toward the polyol pathway depletes the levels of available NAPDH, resulting in impaired NET formation in response to bacterial antigens. 133 Furthermore, lack of NADPH results in impaired activity of glutathione and other antioxidants, leading to increased ROS levels.
Epigenetic changes in diabetic neutrophils
A handful of epigenetic modifications have been identified to regulate neutrophil function, 73 however, studies examining how these are altered in diabetes are sparse. Histone 3 citrullination (conversion of arginine to citrulline) via peptidyl arginine deiminase 4 (PAD4) is a key step in NET formation. H3 citrullination results in the reduction of the protein’s positive charge (rendered by arginine), which loosens the bonds between histones and the negatively charged DNA. 134 Increased levels of PAD4 in patients with DFUs and diabetic mouse wounds increased histone 3 citrullination and their propensity to form NETs (Fig. 9). 122,134
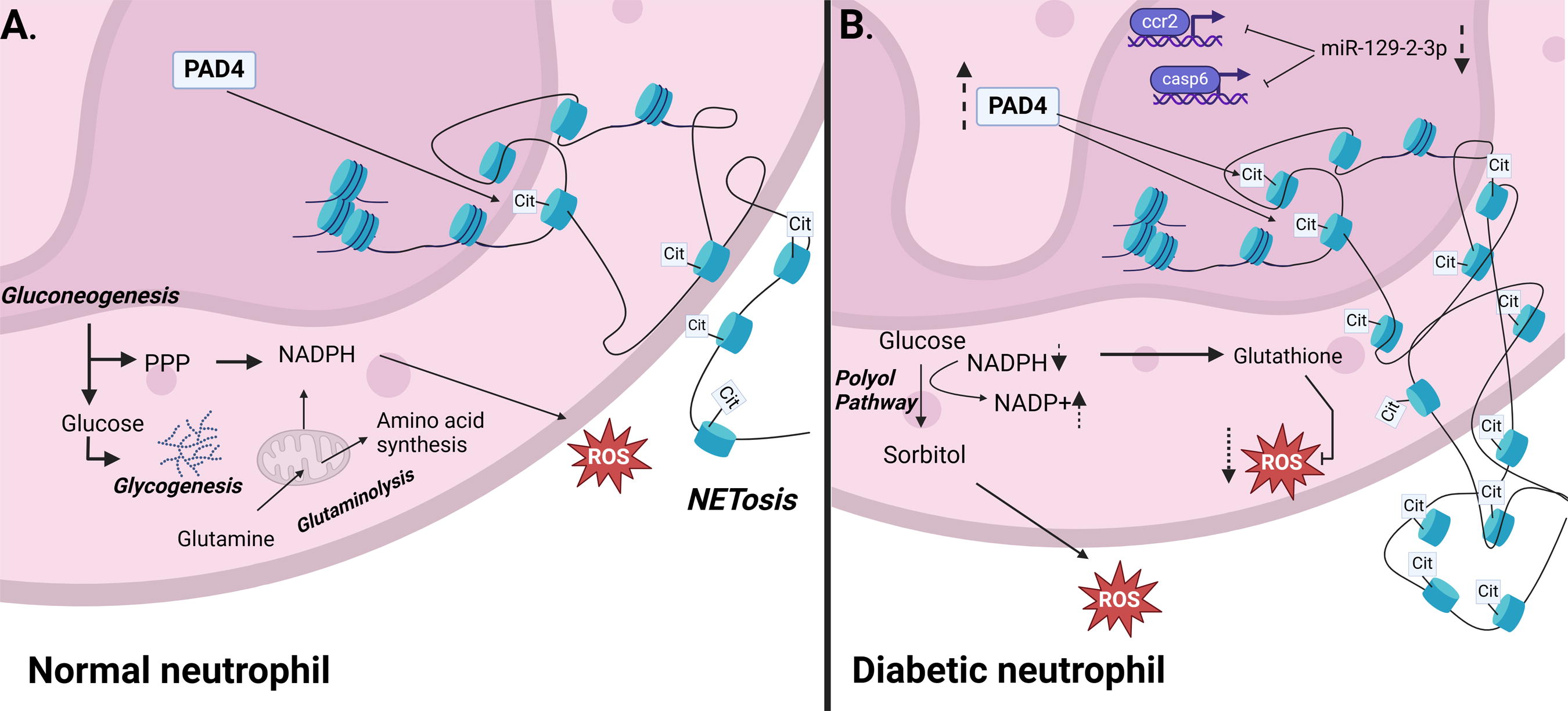
Diabetes disrupts epigenetic regulation inneutrophils.
Diabetic neutrophil function can also be regulated by miRNAs. miR-146a which regulates neutrophil infiltration during the later stages of healing, is decreased in diabetic wounds resulting in delayed wound healing. 135 miR-129-2-3p regulates the expression of caspase-6 and CCR2 and is downregulated in diabetic neutrophils, thus contributing to their malfunction. 136 The expression of circular RNAs, regulators of miRNAs, with miR-129-2-3p and other miRNA binding sites were also found to be altered in diabetic neutrophils (Fig. 9). 136 Furthermore, lncRNAs have been shown to regulate neutrophil activity in other conditions, 137 but studies assessing their effect in diabetic neutrophils are currently lacking.
Macrophage and neutrophil dysfunctions in DFUs
Similarly to diabetic rodents, the functions of monocyte/macrophages and neutrophils are compromised in diabetic patients. 4,138,139 Deregulated numbers of monocytes have been associated with diabetic conditions, 139 and elevated ratios of circulating neutrophils to lymphocyte are associated with a higher risk of non-healing DFU development. 140
To study healing mechanisms taking place at the DFU wound site, researchers usually use tissues isolated during debridement. Debridement is a process during which necrotic tissue is removed and has the potential to “reset” the wound and to promote healing (healing DFUs). Debrided DFUs that do not heal within 12 weeks are considered non-healing.
Transcriptomic analyses of wound tissues showed that after debridement non-healing DFUs exhibit less infiltration, proliferation, and activation of immune cells. 141 –143 Comparison between healing and non-healing DFUs revealed increased levels of inflammatory genes in healers compared with nonhealers (Fig. 10). 142
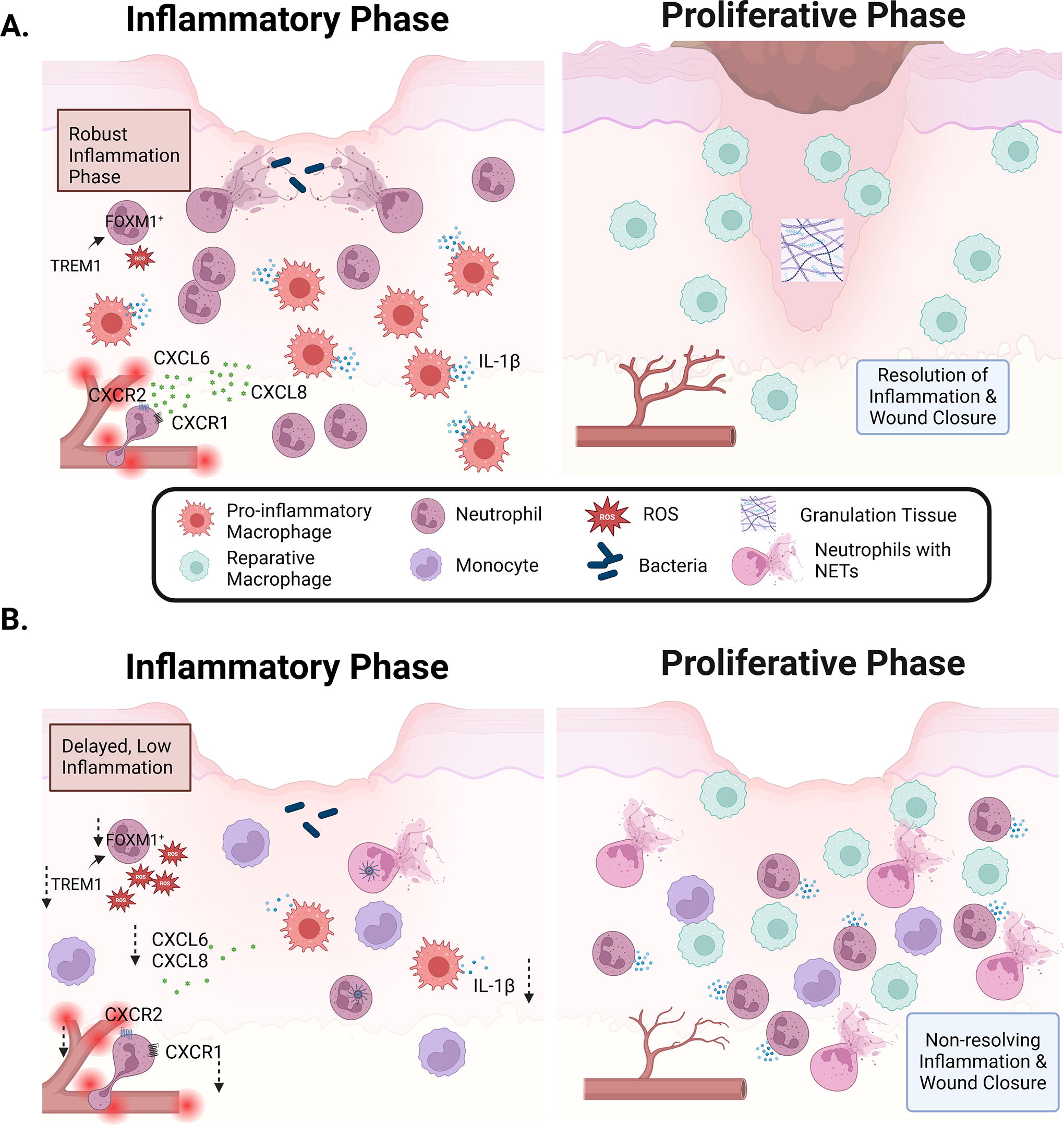
Roles of macrophages and neutrophils in healing and non-healing diabetic foot ulcers (DFUs).
Decreased neutrophilic presence in non-healing DFUs has been documented in multiple studies. Compared with healing DFUs, exudates of non-healing DFUs have decreased levels of CXCL6, a potent neutrophil chemoattractant. 144 In addition, lower IL-1β tissue levels were detected in non-healing DFUs, which was also linked with decreased neutrophilic presence and activity at the wound site (Fig. 10). 145
Interestingly, a specific subset of neutrophils expressing FOXM1 (forkhead box M1), a transcription factor associated with proliferation, was shown to be decreased in non-healing DFUs. In normal healing acute wounds, FOXM1 expressed by wound neutrophils promoted ROS levels and NET formation. These FOXM1+ neutrophils migrate to normal healing wounds in response to TREM1 (triggering receptor expressed on myeloid cells) activation. TREM1 can be expressed by various myeloid cells and is thought to enhance the local immune response. 146 Non-healing DFUs were associated with decreased levels of tissue TREM1 and lower numbers of FOXM1+ neutrophils. 141 Furthermore, decreased levels of the neutrophil receptors CXCR1, CXCR2, and of one of their ligands (CXCL8) were documented in non-healing compared with healing DFUs (Fig. 10). 141
In addition to their sparse presence early after debridement, neutrophils in non-healing wounds do not function properly. Diabetic patients with DFUs have increased NETosis markers both in the circulation and at the wound site compared with diabetic patients without DFUs. 122,141 This increased NETosis is associated with NLRP3 inflammasome activation. 125
Notably, while monocytes are readily observed in DFUs, macrophages are mostly absent.43 A recent study showed that macrophages in DFUs are characterized by increased expression of apoptotic genes, suggesting that macrophages may be short-lived in the DFU environment.34 Intriguingly, DFUs also showed increase markers of efferocytosis. However, due to the decrease in macrophage numbers, the cells primarily responsible for efferocytosis, the efficiency of apoptotic cell removal in DFUs remains uncertain. 147,148
Macrophages from healing DFUs express inflammatory genes (notably Il1b, in addition to S100a and S100b) early in the healing process, likely resulting in the successful triggering of the resolution phase and therefore promoting wound healing. 149 In contrast, non-healing DFUs exhibit prolonged inflammation despite the presence of pro-reparative macrophages, and they were shown to be unable to progress into an effective proliferation phase. 143
Furthermore, and similarly to mouse studies, 48,66,120,121,149 time-course analysis indicated that non-healing DFUs exhibit delayed inflammatory macrophage presence. Healing-DFUs exhibited a robust acute inflammatory phase in the first few weeks following debridement but these responses dampen starting at the third week following debridement. 150 On the contrary, non-healing DFUs had low inflammation in the first few weeks following debridement but began to increase after week 3 and continued into week 4. Scoring DFUs based on these inflammatory markers was suggested to be an accurate predictor of whether a DFU would heal in a timely manner or become chronic. 150
These suggest that an acute threshold of inflammation levels must be reached to successfully transition to a resolving and reparative phase. 143
CONCLUSIONS
Diabetic conditions impair the functionality and migration patterns of both neutrophils and macrophages in wounds, perpetuating unresolved inflammation and hindering healing. Moreover, increased myelopoiesis and metabolic impairments further exacerbate complications associated with diabetes.
Efficient wound healing typically involves the initial infiltration of neutrophils and macrophages into the wound site and a robust inflammatory phase. Subsequently, inflammation diminishes as wound neutrophils and macrophages transition toward anti-inflammatory and pro-reparative phenotypes, ultimately leading to successful wound closure. In diabetic conditions, both the initial inflammatory responses of neutrophils and macrophages and their subsequent transition to anti-inflammatory and pro-reparative phenotypes during later stages of wound healing are dysregulated. In addition, diabetic neutrophils and macrophages exhibit many functional impairments, which further contribute to defective infection control and impaired healing.
Diabetic macrophages display persistent epigenetic and metabolic changes and express markers associated with both inflammatory and pro-reparative phenotypes, but their activation process lacks linear progression. This results in a deficient presence of robust inflammatory macrophages early in the healing process and a flawed transition to the pro-reparative phenotypes, which are crucial for successful healing. Understanding the mechanisms that promote early recruitment of inflammatory macrophages to the wound site and facilitate their transition to a pro-reparative phenotype represents a potential treatment strategy for DFUs.
Diabetic neutrophils are constitutively activated but fail to clear infections at the diabetic wound site, because of dysregulation in their influx dynamics and their bactericidal functional impairments. Recent research highlights the significance of inadequate neutrophil influx during the inflammatory phase early after injury as a major culprit for making diabetic wounds vulnerable to infection and impaired healing. They also highlight the therapeutic potential of immunomodulators that can enhance infection control and stimulate healing by jumpstarting the neutrophil response in the diabetic wound during the acute phase of healing early after injury.
The roles of neutrophils and macrophages in the wound healing process are still under investigation. Current studies suggest that healing DFUs exhibit a robust early inflammatory phase and transition to a productive proliferative phase, ultimately leading to effective wound closure. In contrast, non-healing DFUs are characterized by delayed infiltration of inflammatory cells at the wound and display defective activities resulting in non-resolving inflammation. Understanding the mechanisms of neutrophil and macrophage activities at the wound site offers a novel pathway for the development for new DFU therapies.
Footnotes
ACKNOWLEDGMENT
Figures were created with biorender.com
ABOUT THE AUTHORS
AUTHOR DISCLOSURE AND GHOSTWRITING
The authors declare no conflicts of interest and no ghostwriters. The manuscript was cowritten by the listed authors.
FUNDING INFORMATION
This work was supported by the Shriners Hospitals for Children (71002-NCA-23) and NIH NIGMS (R01GM135279) to AMS, and NIH NIDDK (R01DK107713, R01DK135557) to SHS.