
Abstract
Significance:
It has long been hypothesized that naturally occurring electric fields (EFs) aid wound healing by guiding cell migration. Consequently, the application of EFs has significant potential for promoting wound healing. However, the mechanisms underlying the cellular response to EFs remain unclear.
Recent Advances:
Although the directed migration of isolated single cells under EFs has been studied for decades, only recently has experimental evidence demonstrated the distinct collective migration of large sheets of keratinocytes and corneal epithelial cells in response to applied EFs. Accumulating evidence suggests that the emergent properties of cell groups in response to EF guidance offer new opportunities for EF-assisted directional migration.
Critical Issues:
In this review, we provide an overview of the field of collective electrotaxis, highlighting key advances made in recent years. We also discuss advanced engineering strategies utilized to manipulate collective electrotaxis.
Future Directions:
We outline a series of unanswered questions in this field and propose potential applications of collective electrotaxis in developing electrical stimulation technologies for wound healing.
Yan Zhang, PhD

Min Zhao, PhD, MD

SCOPE AND SIGNIFICANCE
This review describes the diverse characteristics of collective electrotaxis and compares it to single-cell electrotaxis. In addition, this article presents advanced engineering strategies for studying collective electrotaxis. The primary objective of this review is to summarize the existing information and create an up-to-date educational resource for researchers who are new to this topic. Furthermore, this review emphasizes the importance of this emerging field and draws attention to it.
TRANSLATIONAL RELEVANCE
The application of electric fields (EFs) to promote wound healing has gained widespread attention from biomedical engineers, and many EF-assisted bandages have since been designed. The emergent property of synergies exhibited by cell groups in response to EF guidance and the current demonstration of a brilliant electrical stimulation strategy based on the emergent property of synergies presented in this review will expand the design thinking of designers in this field and pave the way for the next generation of EF-assisted bandages.
CLINICAL RELEVANCE
The success of electroceutical bandages in controlling infection in chronic wounds has shown the efficiency, safety, and ease of EF-assisted bandages in clinical settings. EF-assisted bandages, which promote wound healing by accelerating directional cell migration, hold great potential to be the next breakthrough. A deep understanding of the electrical guiding mechanism for cell migration under physiological conditions must be achieved to develop effective clinical applications for wound healing. The discoveries in collective electrotaxis reviewed in this article deepen our understanding not only of cell electrotaxis but also of the natural process of wound healing.
BACKGROUND
Directional migration is crucial in wound healing, embryo development, and cancer invasion. 1,2 In recent years, mounting evidence has suggested that cells often move as tightly or loosely associated cohesive groups during these processes. 2 This type of migration is known as ‘collective cell migration.’ Cells undergoing collective migration maintain physical and functional connectivity with each other, polarizing at the cell group level. Cells collectively migrate in various forms in vivo, including two-dimensional patches of cells migrating across tissue surfaces, e.g., wound healing, or multicellular germination traversing three-dimensional tissues, e.g., vascular outgrowth or tumor invasion. 2 The most attractive phenomenon in collective cell migration is the emergence of synergies from the activities of individuals that exceed what can be predicted from their individual capabilities, enabling cells to perceive and respond to signals in complex and dynamic environments. Cells in a group migrate more consistently and efficiently than isolated single cells. 3 Cells in a group can also detect the substrate geometry and migrate accordingly in a cell-density–dependent manner. 4 Furthermore, keratinocyte monolayers can crossover the heterogeneous extracellular matrix by forming epithelial bridges and maintaining tissue integrity. 5 The growing interest in the field has led to a better understanding of the emergent behaviors and underlying mechanisms of directional collective cell migration. Contact inhibition of locomotion, 6 traction force transmission, 7,8 and actomyosin contraction 9 are critical for collective cell migration. Many literature reviews on the mechanisms underlying collective cell migration have been published. 2, 10 –12
EFs rise naturally in wounds. Numerous experiments over nearly two centuries from different laboratories have demonstrated the existence of wound EFs. 13 –16 Physiological EFs (42–500 mV/mm) accumulate in normal tissue up to 500 micrometers away from the edge of the skin and corneal wounds and point toward the wound center. These wound-induced EFs persist until the migrating epithelial cells at the leading edge reseal the wound. 13 This phenomenon is receiving increased attention because of its powerful ability to direct cell migration and its ability to facilitate wound healing (Fig. 1). For decades, researchers have focused on studying the molecular signals involved in electrotaxis 19,20 and the role of charged components on cell membranes in the EF sensing of isolated single cells. 21 –24 Only recently has experimental evidence demonstrated that large sheets of keratinocytes or corneal epithelial cells collectively migrate in response to applied EFs. 15,18, 25,26 While some phenomena of collective electrotaxis resemble those of isolated cells, there are also distinct coordinated movements of cohesive groups.

Cells migrate collectively during wound healing, and EFs facilitate directional collective migration.
Using advanced tools such as cell patterning, 17 particle image velocity, 17 traction force microscopy, 27,28 microfabrication, 29 –31 and electrical engineering, significant progress has been made in unraveling the hidden secrets of collective electrotaxis. The aim of this literature review is to explain how EFs direct collective migration, investigate how cell sheets sense and integrate EF signals, and engineer techniques for programming collective electrotaxis and propose potential directions for future research.
DISCUSSION
A cell group carries out electrotaxis as a single unit—“a supercell”
Supracellular migration is a type of collective migration characterized by the movement of a group as a cohesive unit rather than individual members. 11 This phenomenon has been observed in numerous collective cell migration processes, such as polarity, cytoskeletal organization, force transmission, and motility flows. These observations have been extensively reviewed in the literature. 11 In addition, supracellular migration has been observed on multiple levels during collective electrotaxis.
First, cell groups migrate as a unified entity and generate a wave-like propagation pattern of motility. When an EF of 200 mV/mm is applied to human skin keratinocyte cell sheets at a scale of square millimeters through a classical electrotaxis chamber, 17 collective migration becomes evident within 30–60 min. 26 The leading edge, the free edge with a default migration direction toward the EF-guided direction, extends forward, while the rear edge, with a default migration direction opposite to the EF-guided direction, retracts to migrate toward the anode. As a result, the entire sheet migrates in a directed manner. The cell sheet maintains its integrity and resembles a large organized unit that moves in response to the EF 26 (Fig. 2A–C). Moreover, the expansion of the cell sheet is restrained both parallel and perpendicular to the EF lines 26 (Fig. 2, Table 1). Directional collective migration of keratocytes in zebrafish 25, 40 and goldfish 41 has also been observed in vivo in response to EFs of 200∼256 mV/mm. Using the advanced motility visualization technique of particle image velocimetry, researchers observed a three-phase wave-like propagation pattern of directionality across the cell sheet. This wave initiation occurs at the leading edge and ultimately leads to the directional migration of cell sheets as coherent units. The propagation rate slowed when the magnitude of the EF decreased (Fig. 2D, E). In addition, the propagation wave can be disrupted by disruption of cell–cell adhesion, 25,26 suggesting the vital role of cell–cell interactions in the formation of motility waves. In the keratocyte cell sheet of zebrafish, the synchronous motility state induced by inhibition of phosphoinositide 3-kinase (PI3K) across the cell cluster before electrical stimulation can prevent the occurrence of directional waves after stimulation. 25 This indicates that the generation of directionality waves relies on the directional migration inhomogeneity within the cell sheet before EF stimulation.

Well-patterned large epithelial sheets further revealed the EF-induced propagation dynamics of collective electrotaxis.
Comparison of the electrotaxis of isolated single cells and cell sheets
+, higher/stronger compared with isolated cells in the absence of EFs.
++, higher/stronger compared with isolated cells in EFs.
Second, molecular signals are activated in the cell sheet as a cohesive unit during collective electrotaxis. PI3K plays a crucial role in the signaling network at the leading edge of single cells in the EFs. 18 Inhibiting PI3K impairs the directed migration of single cells in EFs in various cell types. 18 Notably, for zebrafish keratocytes, inhibiting PI3K leads to a reversed directional migration of isolated zebrafish keratocytes at EFs of 400 mV/mm. 39 Moreover, the velocity gradient in zebrafish keratocyte sheets reversed upon PI3K inhibition. This can be explained by the fact that PI3K predominantly affects the peripheral cells in the sheet, whereas the inner cells appear independent of PI3K. By increasing the contractile force in the posteriormost cells, the inner cells sufficiently establish a directional response to EFs, resulting in a reverse velocity gradient 25 (Table 1). The study strength will be enhanced by visualizing the activation of PI3K across the cell sheet in EFs. The distinct response of isolated cells and cell sheets of zebrafish keratocytes to PI3K inhibition also implies cohesive activation of molecular signals at the supracellular level in the cell sheet. Further investigation is required to explore how other cell groups respond to PI3K inhibition in response to EFs.
Third, cell groups exhibit more efficient electrotaxis. Cell–cell interactions are innate characteristics of collective cell migration that promote coordinated collective movement, and tight cell–cell adhesion plays a crucial role in tissue function. In MDCKII cell sheets, an EF of 300 mV/mm increases the correlation length parallel to the field axis, whereas weaker correlations are observed in the orthogonal axis. There seems to be a trade-off between these two correlation lengths. 34 In experiments using MDCKII cells and primary cultured bovine corneal epithelial cell sheets with fewer than 100 cells, a size-dependent response to an EF of 200 mV/mm was observed. Compared with those of isolated single cells, monolayers of cells displayed greater directness, increased speed, and improved migration efficiency (Fig. 3A-I). E-cadherin, but not gap junctions, is essential for hyperefficiency collective cell migration. 35 A similar phenomenon has also been observed in zebrafish keratocyte sheets. 25 However, for larger cell sheets, such as human skin keratinocytes containing more than 1000 cells, no significant differences in speed or directionality were observed between cell sheets of different sizes in response to EF guidance. 26 Similar hypersensitivity and efficiency of cell clusters have also been observed in collective chemotaxis 3 and durotaxis. 8 It is widely accepted that once the cell number exceeds a certain threshold, no further significant improvement in hypersensitivity or efficiency can be detected, as the coordination of collective cell migration reaches its maximum potential. 11 The same principle applies to collective electrotaxis. Furthermore, paracrine effects have also been found to play a role in coordinating the polarization of leader cells in collective electrotaxis. 42

More sensitive and efficient electrotaxis in cells collectively in cell groups than in cells in isolation.
Fourth, electrical stimulation can modulate the mechanical state of the cell sheet as a unit. In isolated single MDCKII cells, the traction force is primarily concentrated on the cell periphery, with force vectors pointing toward the center of the cell. The traction force of isolated cells does not exhibit specific directionality, and the average directionality values are close to 0. EF stimulation did not significantly change the traction force distribution in isolated cells. 35 A similar traction force distribution was also found in isolated primary bovine osteoblasts and human osteosarcoma cells in EFs. 38 In MDCKII cell sheets, traction force analysis showed that traction force is also mainly concentrated on the edge of the cell sheet. The leading cells within the cell sheet generate traction force directed toward the rear of the cell sheet. An EF of 200 mV/mm enhances the directed traction force of leading cells, but has no significant effect on cells in the center or rear edge of the cell sheet, suggesting that the orientation of the traction force at the leading edge plays an important role in the collective electrotaxis of cell sheets 35 (Fig. 4A, Table 1). In addition, traction force microscopy revealed traction force and intercellular stress in the center of the human keratinocyte cell sheet aligned perpendicular to the EF vector (50 mV/mm) during electrotaxis 33 (Fig. 4B–C, Table 1). Researchers have also found that an EF of 300 mV/mm significantly biases the collective strain rate wave propagation perpendicular to the EFs in MDCKII cell sheets. 34
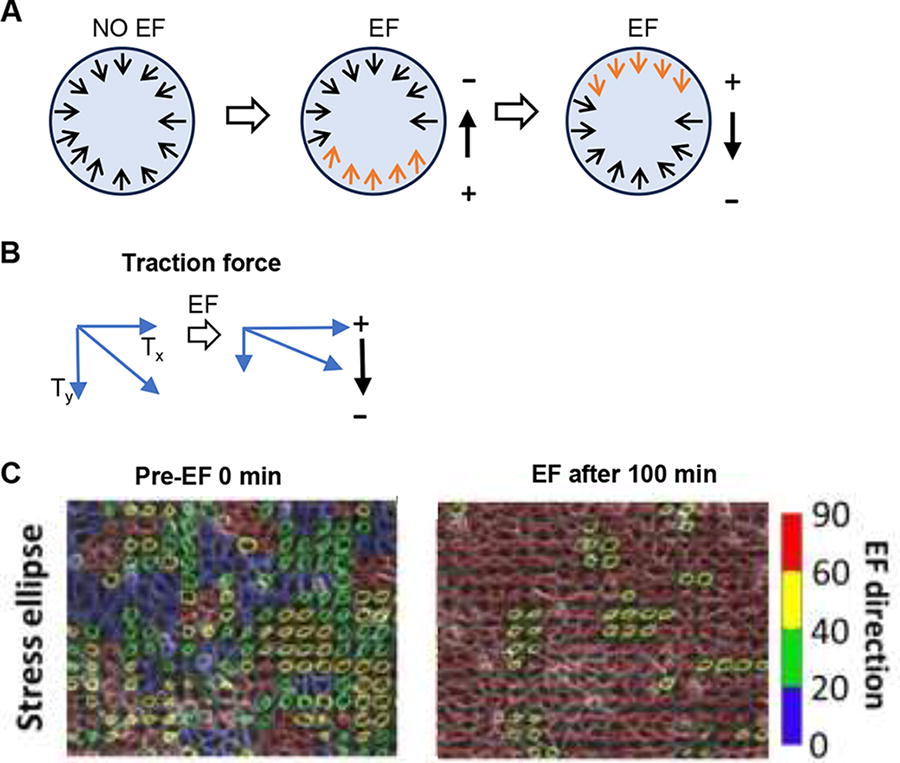
EFs modulate the mechanical state of collective migrating cell sheets.
A cell group senses EFs—individual sensing versus collective sensing
It is widely recognized that isolated individual cells can migrate in a directed manner toward either the anode or cathode with uniform EFs. However, collective cell migration is highly complex. First, the variation in both the strength and direction of the EF cannot be neglected when considering the migration of a large group of cells, particularly in vitro. Second, cells in a cell cluster are coupled with each other through cell–cell adhesion and gap junctions. This coupling allows for the possibility that cells within the cluster are integrated physically, chemically, and electrically, forming a larger supracellular structure. However, it remains an open question whether individual cells in a cluster can independently sense EF guidance.
As proposed by Sun et al., 25 several points support the individual and independent sensing of EFs by cells in a cluster. First, the threshold at which zebrafish keratocyte cell groups sense an EF is the same for individual cells. 25 Conversely, Li et al. 35 and Zhu et al. 32 reported that the MDCKII and breast cancer cell clusters appear more sensitive to EF guidance than isolated cells. In an EF at low strength, monolayer cells migrate directionally, whereas isolated cells do not display any directional migration. 35 We agree that cells in groups and isolated cells perceive EFs consistently. This behavior can be better described as hyperefficiency but not as hypersensitivity of cell groups to weak EF guidance. Similar phenomena have also been observed in the collective chemotaxis 3 and durotaxis. 8 Second, Sun et al. reported that in a PI3K inhibitor-pretreated cell group, cells located in different regions of the cell sheet responded synchronously to the EF guidance. 25 The region dependence in response to EFs may primarily be due to the distinct motility state of the cell before EF stimulation rather than differences in the sensing of EFs. 25,26 Third, initiating collective motility takes a similar amount of time for cells in the bulk of a sheet under EFs as for single cells to polarize under EFs. 25 Furthermore, previous studies have shown that cells can migrate toward convergence when subjected to a tripartite field generated by a ‘T’-shaped chamber. 36 With a microfluidic chip equipped with multiple electrodes, our previous study showed that collective cell migration closely followed the field lines created by convergent EFs without being constrained by the chamber (Fig. 5A–C). 26 In summary, we propose that cells in groups can sense EFs individually and independently.
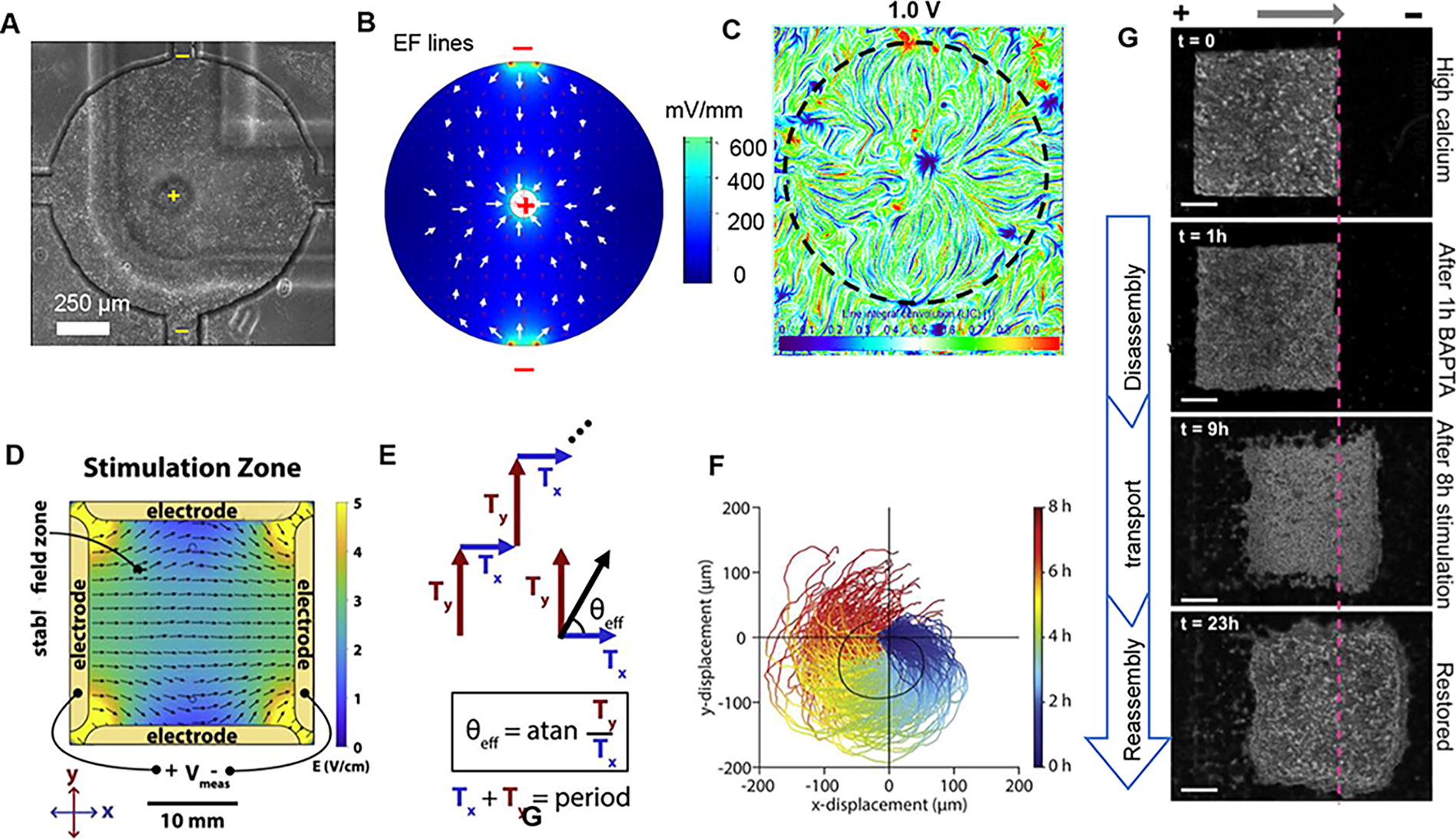
Engineering collective electrotaxis.
Cells collectively integrate directional cues
It is challenging to separate EFs from other directional cues in the microenvironment of a cell group, both in vivo and in vitro. How cells integrate EF signals and other directional cues to make collective decisions remains unclear. In a precisely patterned square monolayer of human skin keratinocytes, the directional cues arising from free edges were effectively regulated. When an EF of 200 mV/mm was applied, cells located at the frontier of the region exhibited direct migration toward the external EF guidance, aligning with the original migration direction. In contrast, cells located elsewhere initially had different directionality but gradually adopted a migratory trajectory like the leading-edge cells over time. Specifically, cells positioned in the forefront area exhibited cathodal migration, which was followed 20 min later by the trailing rear region cells, showing similar directionality and speed within the cell sheet. However, cells in the side region migrated consistently with bias along the EF lines. At the rear edge of the cell cluster, a tug of war between different strengths of EF guidance and free edge signals was observed. 26 This phenomenon has also been observed in MDCK epithelial monolayers, 34 corneal epithelium cells, 34 breast cancer cells, 32 and zebrafish keratocytes. 25 These results imply that cells in the cell sheet integrate EFs and free edge direction signals by following the vector-like addition interaction.
Furthermore, once a three-hour EF (300 mV/mm) stimulation is halted, the tissue retains a directional memory that gradually decays exponentially. This lasting effect typically lasts up to 4 h in MDCK epithelial monolayers, and its duration depends on the cell density. A higher cell density leads to a longer-lasting effect duration (Table 1). The lasting effect on the spatial correlations of the cell sheet can reach 6–10 h. 34 A lasting effect has also been observed in zebrafish keratocytes after 30 min of EF-induced migration (EF = 200 mV/mm). 25 The minimum duration of EF stimulation required for a cell group to acquire memory has not been thoroughly investigated. The lasting effect influences how cells integrate directional signals that emerge in a specific order. A similar lasting effect is observed when the EF reverses. During the first hour following EF reversal, most cells in the cell sheet maintain their original migration behavior in human skin keratinocyte cell sheets and MDCK epithelial monolayers. 26,34 However, after the initial hour, these cells gradually reversed their migration direction. Moreover, the regions at the right and left of the cell sheet gradually undergo a U-turn after the EF reversal. 26 This U-turn phenomenon has also been observed in MDCKII cell monolayers 36 and zebrafish keratocyte sheets 25 in response to reversed EF. It is believed that this U-turn is a dynamic consequence of combining the gradual disappearance of the lasting effect of the previous velocity, the emergence of increasing new EF guidance, and constant guidance from the free edges. 26 These results suggest that the lasting effect of post-EF stimulation also contributes to the integration of directional signals in cell groups as a vector.
Engineering collective electrotaxis of the cell group
Gaining a fundamental understanding of how cells collectively respond to an EF is essential for engineering collective electrotaxis. This understanding encompasses several key aspects, including the threshold at which a group of cells responds to an EF, the time it takes for the group to respond, and the subsequent effects of EF stimulation. Furthermore, comprehending the underlying principles that govern the integration of directional cues and the intercellular interactions guiding cell groups can further enhance the engineering of collective electrotaxis. Building upon this knowledge, chips with multiple biocompatible electrodes have been designed to facilitate the engineering of collective electrotaxis (Table 2). These chips can generate convergent directed current EFs, which have demonstrated the ability to accelerate wound healing in monolayer tissues in vivo (Fig. 5A–C). 26,37,45 In addition, researchers have developed a system that integrates four independent electrodes controlled by a computer to enable two-dimensional manipulation through a sequence of independent migration command pulses along the x- or y-axis (Fig. 5D–F). 43 In primary mouse skin monolayers, inhibition of E-cadherin–dependent cell-cell adhesion was observed to enhance the controllability of electrotaxis. A strategy involving disassembly, collective transport, and reassembly was then developed to enhance the effectiveness of electrotaxis in primary mouse skin monolayers (Fig. 5G). 44 Moreover, by applying EFs of 500–1,000 mV/mm in three-dimensional kidneyoids and gut organoids, cyst symmetry is disrupted through collective electrotaxis. 45
Various systems for collective electrotaxis
Summary and future directions
The wide occurrence of collective cell migration in vivo and the distinct response of cell groups to EF guidance compared with that of isolated cells imply the importance of considering collective electrotaxis in EF-aided wound healing. While numerous clinical devices claim to facilitate wound healing through the application of physical electrical stimulation and may have unknowingly induced collective electrotaxis in previous clinical practices, implementing an advanced EF-aided wound healing clinical device based on collective electrotaxis has not been attempted. The disassembly-collective transport-reassembly strategy in electrotaxis cell groups 44 suggests that future EF stimulation strategy could combine EF stimulation with medicine to induce collective cell migration. Previously reported convergent cell collective migration induced by convergent EFs 26,37,47 and 2D manipulation of collective cell migration by pulsed electrical stimulation 43 promise a precisely controlled collective electrotaxis in tissues in the future. Moreover, the improved response to EF guidance observed in the collective migration 35 allows us to achieve the desired control of cell migration with lower levels of EF stimulation in the cell group compared with isolated single cells. The extended postelectrical stimulation lasting effect in collective cell migration enables short-term EF stimulation to achieve long-term direct cell effects. 34 This also suggests the potential for regional EF stimulation to drive directional collective migration of large cell groups. All the above findings strongly demonstrate the new opportunities for aided wound healing presented by collective electrotaxis. We believe that a deep understanding of collective electrotaxis could lead to novel engineering techniques capable of mobilizing groups of cells to enhance wound healing.
There are also several challenges in applying collective electrotaxis to promote wound healing.
First, some unexpected phenomena have been observed in experiments involving specific cell types, which necessitate further exploration to determine whether these phenomena are specific to those cell types or are common characteristics of cell sheets. For instance, Cohen et al. reported that in MDCKII cell monolayers, leading-edge cells do not respond to the EF guidance. 36 In addition, mouse keratinocyte monolayers with strong cell–cell adhesion do not respond well to EF guidance, and leading-edge cells experience cell death in EFs. 44 Furthermore, in EF-directed zebrafish keratocytes, holes emerge and dissociation from the periphery occurs. 25
Second, more work needs to be done to understand the mechanism of collective electrotaxis. Inhibition of PI3K has resulted in very different outcomes for cell groups compared with single cells in the electrotaxis, 25 suggesting that while some molecular signaling pathways involved in collective electrotaxis resemble those involved in isolated cells, there might also be distinct mechanisms governing the coordinated movement of these cohesive groups. In addition, although current studies suggest that cells in a group sense EFs relatively independently, further experiments should investigate the possibility of electrical coupling within a cell group in EF sensing.
Finally, studies would benefit from an experimental model that resembles in vivo conditions. Current research primarily focuses on homogeneous cell monolayer collective migration on the square millimeter scale. However, it is crucial to explore how EFs direct the collective migration of heterogeneous cell populations at larger scales, both in vivo and in vitro. Coculturing epidermal keratinocytes and dermal fibroblasts in an electric field represents an initial but important step in investigating the electrotaxis of diverse cell populations. 48 Exploring this area could also be greatly facilitated by an advanced EF stimulation system capable of precisely controlling the EF distribution and minimizing water electrolysis. To gain a more comprehensive understanding of directed collective cell migration, it would also be valuable to conduct a comparative study that examines different directional cues within the tissue, such as chemotaxis and mechanical strain, in conjunction with electrical signals.
TAKE-HOME MESSAGES
Cells migrate collectively during wound healing, and EFs facilitate directional collective migration.
Cell group electrotaxis shows the emergence of synergies, as the cell group shows more efficient electrotaxis than isolated individual cells, and the cell group carries out electrotaxis as a single unit—“a supercell.”
Cell–cell interactions play a vital role in collective electrotaxis.
Cells in a group sense EFs relatively independently and convergent EFs induced convergent collective migration.
Collective electrotaxis offers new opportunities for developing electrical stimulation technologies for wound healing.
Footnotes
ACKNOWLEDGMENTS AND FUNDING SOURCES
The authors thank Cell Press, the Federation of American Societies for Experimental Biology, Springer Nature, the American Society for Cell Biology, and the National Academy of Sciences for permitting us to reuse figures in this article. The language in the article has been refined by Open AI’s ChatGPT 3.5. This work was supported by the National Natural Science Foundation of China (No. 51807142) and Hangzhou Normal University Research Start-up Funds (No. 2019QDL031) to Y. Z.
AUTHOR CONFIRMATION
M.Z. and Y.Z. conceived the original idea for this article, Y.Z. wrote the original draft, and all authors provided comments and made edits to the article and figures. All the authors have read and approved the final article.
AUTHOR DISCLOSURE AND GHOSTWRITING
The authors declare that they have no known competing financial interests or personal relationships that could have appeared to influence the work reported in this article. The content of this article was written by the authors listed. The language in the article has been refined by Open AI’s ChatGPT 3.5. All the listed authors have reviewed the final article and accept responsibility for its accuracy. No ghostwriters were used to write this article.
ABOUT THE AUTHORS