
Abstract
Objective:
Volumetric muscle loss (VML) leads to permanent muscle mass and functional impairments. While mesenchymal stromal cells (MSCs) and their secreted factors can aid muscle regeneration, MSCs exhibit limited persistence in injured tissue post-transplantation. Human placental-derived stem cells (hPDSCs), sharing surface markers with MSCs, demonstrate superior regenerative potential due to their fetal origin. Previously, a biosponge (BS) scaffold was shown to augment muscle regeneration post-VML. This study aims to coapply BS therapy and hPDSCs to further enhance muscle recovery following VML.
Approach:
A VML defect was created by removing ∼20% of the tibialis anterior muscle mass in male Lewis rats. Injured muscles were either left untreated or treated with BS or BS-encapsulated hPDSCs cultured under normoxic or hypoxic conditions. On day 28 postinjury, peak isometric torque was measured, and the muscle was harvested for analysis.
Results:
BS encapsulated hPDSCs subjected to hypoxic preconditioning persisted in larger quantities and enhanced muscle mass at day 28 postinjury. BS encapsulated hPDSCs cultured under normoxic or hypoxic conditions increased small myofibers (<500 µm2) percentage, MyoD protein expression, and both pro- and anti-inflammatory macrophage marker expression. BS encapsulated hPDSCs also reduced fibrosis and BS remodeling rate.
Innovation:
This study is the first to examine the therapeutic effects of hPDSCs in a rat VML model. A BS carrier and hypoxic preconditioning were investigated to mitigate low cell survival postimplantation.
Conclusion:
hPDSCs augment the regenerative effect of BS. Combining hPDSCs and BS emerges as a promising strategy worthy of further investigation.

INTRODUCTION
The global economic impact of trauma, including both medical costs and productivity losses, is estimated to exceed $400 billion annually. Trauma plays a significant role in the global burden of disease and injury. 1 Extremity wounds are the predominant form of trauma in military conflicts, often necessitating prolonged hospital stays and rehabilitation. 2,3 Among United States military personnel, soft tissue injuries to the extremities are the most common wounds sustained during wartime operations, 2 comprising a significant portion of surgical procedures for battlefield injuries. 4,5 While these injuries are typically the result of high-energy blast trauma, recent military conflicts have shown that the majority of injuries are to the extremities, frequently involving severe musculoskeletal injuries like volumetric muscle loss (VML). 2,3 Both the lower and upper extremities can be affected by VML, inducing significant functional deficits, muscle weakness, and chronic pain. 6 VML is also prevalent in nonmilitary contexts, often resulting from various traumatic incidents and medical conditions. 7 These include approximately 150,000 open fractures, 8 120,232 firearm injuries, 9 and ∼12,000 cases of soft-tissue sarcomas 10 annually.
Skeletal muscle, endowed for its regeneration capabilities, becomes irrecoverable due to the complete loss of cellular and acellular components such as the basal lamina and resident stem cell (or satellite cell) population. 11 –15 In addition, VML injury exhibits significant loss of contractile tissue, persistent inflammation, extensive fibrosis, and changes in muscle architecture. 12 –14,16,17 Current treatments for reconstructing or repairing this injury are extremely limited. A standard of care includes physical therapy, which can only strengthen the remaining muscle and cannot restore lost tissue. 18 Autologous muscle flaps have been used for VML wound coverage, demonstrating efficacy in reducing early complications. However, they carry the risk of donor site morbidity, exacerbated by the limited availability of donor muscle tissue. 19,20 A wide range of extracellular matrix (ECM) based scaffolds have been applied to animal models for VML repair. No regenerative or tissue-engineered products have received FDA approval for human VML injuries.
Placental-derived stem/stromal cells (PDSCs) are obtained from the chorionic tissue of the placenta, are of fetal origin, and can be obtained from full-term newborn deliveries with full informed consent. Thus, they are not of embryonic origin and avoid ethical and legal issues. Donors are tested and screened for communicable disease agents and diseases. An extensive quality assurance system is applied to each step of harvesting, isolation, manufacturing, and shipment. 21 Because placentas are considered biological waste, PDSCs can be obtained easily and noninvasively compared with MSCs from bone marrow or adipose tissues, without the need of surgical intervention or donor-site morbidity. As reviewed by Antoniadou and David, 22 placenta stem cells adhere to plastic, can be expanded by serial expansion as a cell line, have multilineage differentiation potential, and display features consistent with MSCs. PDSCs share cell surface markers with MSCs, which are CD73+ CD105+ SSEA+ CD45- HLA-DR-. Due to their fetal origin, PDSCs display better proliferation and differentiation capacity than MSCs isolated from adult donors. PDSCs have low immunogenicity in either an allogenic or xenogenic setting due to limited human leukocyte antigen expression.
The primary obstacles of MSC-based therapies include poor survival due to apoptosis, necrosis, or anoikis and the inability to maintain self-renewal. 23 Fewer than ∼5% of transplanted MSCs are reported to survive at the site of injection days after transplantation. 24,25 To circumvent these issues, MSCs have been transplanted with biomaterials. It has been repeatedly shown that MSCs injected with a biomaterial carrier material, typically a viscous protective ECM solution, exhibit better viability and retention in the transplanted tissue. 26 –28 Biomaterials provide adhesion sites and biomechanical support to transplanted MSCs 24,25,29,30 and are also expected to shield the MSCs from inflammatory cells and cytokines while simultaneously permitting the diffusion of oxygen, nutrients, and waste materials.
Hypoxic preconditioning, exposing cells to a low oxygen environment (1–5% O2), mimics the in vivo microenvironment and aids cell survival post-transplantation. Standard cell culture conditions (21% O2) may not sufficiently prime cells for hostile injury microenvironments, which typically lack oxygen, essential ECM components, and growth factors. Hypoxic exposure enhances therapeutic potential by promoting cell survival, proliferation, motility, trophic factor secretion, and metabolic adjustments post-transplantation. 31,32 This strategy demonstrates efficacy across various tissue injury applications. 33 –37
Previously, a biosponge (BS) scaffold composed of ECM proteins enhanced muscle regeneration and function after a VML injury in a rodent model. 38,39 In this study, human placental-derived stem cells (hPDSCs) were encapsulated within the BS to enhance muscle recovery further. We hypothesized that BS encapsulation coupled with hypoxic preconditioning would enhance the cell residence time in the VML defect region and improve their survival in a hostile and inflamed wound environment. We expected the ECM biosponge to provide a regenerative substrate to host cells 40 and the PDSCs to release various signaling molecules or growth factors to support muscle regeneration. 41 Therefore, the goal of this study was to determine the extent of functional muscle regeneration following BS-encapsulated hPDSC treatment of VML-injured muscle.
INNOVATION
The present study is the first to examine the therapeutic effects of hPDSCs in a rat model of VML injury. In this work, hPDSCs were delivered using a BS scaffold with hypoxic preconditioning. Using a BS carrier to deliver stem cells can mitigate poor cell survival and engraftment post-transplantation. In addition, BSs can reconstitute vital ECM proteins to the defect site, serve as a temporary scaffold for infiltrating cells, and provide mechanical support to the remaining musculature. Hypoxic preconditioning can further improve stem cell therapy by promoting cell persistence and trophic factor release.
Clinical relevance
There are currently no tissue-engineered therapies accessible for treating VML. The clinical utility of stem cell treatments is impeded by various challenges such as suboptimal in vivo engraftment, compromised bioactivity, and uncertain safety profiles. Using a biosponge carrier to deliver stem cells can address some of these challenges. Our results suggest that biosponge encapsulated and hypoxic preconditioned hPDSCs can improve muscle recovery following VML injury.
METHODS
Cell culture
hPDSCs were obtained from an existing Institutional Review Board (IRB#00056941) approved master cell bank at the Wake Forest Institute for Regenerative Medicine (WFIRM), Wake Forest University School of Medicine (Winston-Salem, NC). Placentas were obtained from healthy donors with informed consent. Placentas are deidentified after collection, and the current research team is not expected to request access to any patient information. All cell culture experiments were conducted according to a protocol approved by Saint Louis University’s Institutional Biosafety Committee (IBC protocol ID: 2021–00022). The hPDSCs were cultured in growth media containing α-minimum essential medium (65%; Gibco, Waltham, MA), AmnioMAX-C100 basal medium (17%; Gibco, Waltham, MA), fetal bovine serum (15%; Sigma Aldrich, St. Louis, MO), GlutaMAX-I CTS (1%; Gibco, Waltham, MA), AmnioMAX-C-100 supplement (2%; Gibco, Waltham, MA), gentamicin (2.5 μg/mL, Gibco, Waltham, MA), and 1% penicillin/streptomycin (Gibco, Waltham, MA) and used from passages 6–8 for all experiments. The hPDSCs were tested for multiple adventitious agents before freezing into 1 mL ampules. hPDSCs were stored in vapor phase liquid nitrogen until use.
Biosponge preparation
Sterile deionized water was heated to 50°C to make a 3 wt% porcine skin gelatin (Sigma Aldrich, St. Louis, MO) solution. After complete dissolution, 20 mM of 1-ethyl-3-(3-dimethyl aminopropyl) carbodiimide (EDC; Sigma Aldrich, St. Louis, MO) and 8 mM of n-hydroxysuccinimide (NHS; Sigma Aldrich, St. Louis, MO) were added to the gelatin solution. Rat tail collagen I (9.50 mg/mL; BD Corning, Corning, NY) was diluted in 1X phosphate-buffered saline (PBS, Gibco, Waltham, MA) to a concentration of 3 mg/mL. In a 6-well plate, 3.5 mL of the gelatin solution was combined with 1.5 mL of the collagen solution and 42 μL of laminin (LM)-111 solution (6 mg/mL; R&D Systems, Minneapolis, MN). The final concentrations of the components were 21 mg/mL of gelatin, 0.9 mg/mL of collagen, 50 μg/mL of LM-111, 14.6 mM of EDC, and 5.6 mM of NHS. The solution was polymerized in the refrigerator (4°C) for 1 h and was frozen overnight at −8°C. The well plate was then moved to −80°C for a minimum of 72 h before lyophilization for 12–14 h. Afterward, the BS was disinfected with 70% ethanol for 5 mins and washed twice with 1× PBS for 5 mins before being frozen at −20°C until needed.
Encapsulation of hPDSCs within the BS
Prior to cell seeding, BS was thawed, disinfected again in ethanol, and rinsed. The BS was then biopsy punched (10 mm), placed in a 48-well plate, and air-dried for 3–5 h in a sterile biosafety cabinet. Then, a cell suspension (50 μL) of hPDSCs at a density of 10 million cells per milliliter was added dropwise to each BS carefully. Cellular attachment of the BS took place in an incubator (37°C, 5% CO2) for 15–30 min before replenishing the cells with growth media (total volume of 500 μL). Cell culture supernatants were collected and replaced at days 1, 3, and 5 (n = 5/timepoint) from every hPDSC-BS. Enzyme-linked immunosorbent assay (ELISA) was used to quantify the following trophic factors: interleukin-6 (IL-6; Abcam, Cambridge, MA), hepatocyte growth factor (HGF; Invitrogen, Waltham, MA), vascular endothelial growth factor (VEGF; Abcam, Cambridge, MA), follistatin (Peprotech), amphiregulin (Abcam, Cambridge, MA), fibroblast growth factor 2 (FGF2; Abcam, Cambridge, MA), and hypoxia-inducible factor-1α (HIF-1α; Abcam, Cambridge, MA). The manufacturer’s instructions were followed for each ELISA.
In a separate experiment, hPDSCs were stained with Qtracker 525 (Invitrogen, Waltham, MA) following the manufacturer’s recommendations before encapsulation. The BS encapsulated hPDSCs (PDSC-BS) recovered for a day before the PDSC-BSs were fixed in 4% paraformaldehyde and then stained with 4′,6-diamidino-2-phenylindole (DAPI). The PDSC-BSs were imaged using laser scanning confocal microscope (Leica Confocal SP8, Leica Microsystems Inc.) at 10X magnification.
Hypoxic and normoxic culture of hPDSC-BS
As described above, a 10 mm BS disk was seeded with 500,000 hPDSCs in two separate 48-well plates (n = 5/time-point). Both plates were placed in the incubator and cultured under standard or normoxic (21% O2) culture conditions (N-PDSC-BS; N) for 24 h. After 24 h of recovery, one of the plates was subjected to hypoxia conditioning (H-PDSC-BS; H) in a hypoxic chamber (STEMCELL Technologies), which was flushed with a mixed gas composed of 3% O2, 5% CO2, 92% N2, and then transferred to the incubator (37°C, 5% CO2). The chamber was flushed every 24 h throughout the experiment. Cell culture supernatants were collected from days 1, 3, and 6 (n = 5/group/timepoint) for ELISAs.
Cell viability
A CyQUANT cell proliferation assay (Invitrogen, Waltham, MA) was performed to assess cell viability by quantifying the DNA content. After growth media was removed and a 2-min 1× PBS wash, the PDSC-BS was placed in a 2 mL centrifuge tube containing the manufacturer’s cell lysis buffer (300 μL). While the tube was on ice, the BS was finely minced to maximize the cell lysis buffer exposure to the hPDSCs within the BS. The minced PDSC-BS solution was then filtered using a cell strainer to remove BS fragments, and the volume of the filtered solution was recorded. The DNA standard solutions and lysed samples were added to a black 96-well plate according to the manufacturer’s instructions. An acellular BS was used as the background control. The fluorescence of the samples was recorded from Em: 480 nm and Ex: 520 nm to determine DNA concentration (SpectraMax i3, Molecular Devices). The DNA concentration (ng/mL) was multiplied by the sample volume to accurately determine the DNA content (ng)/sample.
Animal experiments
All animal work was conducted in compliance with the Animal Welfare Act, the implemented Animal Welfare Regulations, and in accordance with the principles of the Guide for the Care and Use of Laboratory. All animal procedures were approved by Saint Louis University’s Institutional Animal Care and Use Committee (Animal Protocol # 2645).
Adult male Lewis rats (10–12 weeks old; Charles River Laboratories) were housed in a vivarium accredited by the Association for Assessment and Accreditation of Laboratory Animal Care International, which provided water and food ad libitum. The rats were randomly assigned to experimental groups. The animals were weighed and anesthetized using 2.5% isoflurane (Covetrus North America, Dublin, OH). The surgical site was shaved and aseptically prepared using 10% povidone-iodine (Covetrus North America, Dublin, OH) and 70% isopropyl alcohol (Fisher Chemical, Waltham, MA). The skin of the lower leg was laterally incised to reveal the tibialis anterior (TA) muscle. The skin was separated from the fascia and underlying musculature using blunt dissection. An incision was made on the fascia, which was gently separated from the TA muscle using a blunt probe. The TA muscle was bluntly separated from the extensor digitorum longus and the tibia. To create the VML injury, a flat metal spatula was inserted underneath the TA muscle. A 6-mm biopsy punch removed ∼20% of the TA muscle’s mass. The biopsied mass was weighed for consistency. Any bleeding was controlled with light pressure using sterile gauze. The metal spatula was removed, and the injury was either left untreated or filled with an acellular or cellular BS (6-mm diameter). The BS was implanted seeded side face-up in the VML defect. The fascia was pulled over the BS and sutured together with Prolene (6–0; Ethicon) sutures. The skin incision was closed using simple interrupted Prolene (5–0; Ethicon) sutures followed by skin staples. Sustained release buprenorphine (1 mg/kg; Wedgewood Pharmacy, Swedesboro, NJ) was administered at the nape of the neck subcutaneously for pain management at the time of surgery. The surgery was executed unilaterally, leaving one leg uninjured as a contralateral control. Investigators were blinded to group allocation during data collection, and animals were identified by numbers.
Following the VML injury, the animals received No Repair (NR), an acellular BS (BS), N-PDSC-BS, or H-PDSC-BS treatments. For implantation, 6 mm biopsy punches of the BS were made and placed in a 96-well plate. After cellular attachment, the PDSC-BS were moved to a 48-well plate (∼24 h after cell seeding). The PDSC-BS was either cultured in normoxic or hypoxic conditions for 3 days before implantation. The animals were allowed 28 days of recovery. The peak isometric torque was measured, animals were euthanized, and the muscle tissue was harvested. Upon collection, the TA muscles were weighed and cut in half for either histological analysis or biochemical analysis. The upper half was preserved for histological analysis using 2-methyl butane (Fisher Scientific, Waltham, MA) super-cooled in liquid nitrogen. The bottom half of the TA muscle was flash-frozen in liquid nitrogen for biochemical analysis.
Histological analysis
The upper half of the TA muscle cut at the defect site was frozen in 2-methyl butane super-cooled by liquid nitrogen for 10 s. 39,40 The frozen muscles mounted on stubs using optimal cutting temperature embedding medium (Fisher Healthcare, Waltham, MA) were cryosectioned to obtain transverse cross-sections (15 µm). Histological analysis was performed on a single muscle cross-section, comprising both the defect region and the remaining muscle mass. The entire cross-section was used for analysis in certain assessments, whereas in others, multiple images per cross-section were utilized. The cross-sections were stained with hematoxylin and eosin (H&E) to visualize myofiber morphology and BS remodeling. Cell nuclei were identified using (DAPI; 1:100; Invitrogen, Waltham, MA) staining. Biodistribution of the hPDSCs within the injured muscles was visualized using anti-human Ku80 (1:100; C48E7, Cell Signaling Technology, Danvers, MA). 42 Muscle cross-sections were also stained with embryonic myosin heavy chain (eMHC, 1:10, F1.562; Developmental Studies Hybridoma Bank) to quantify regenerating muscle fibers. Macrophage infiltration was determined using CD68 (1:50; MCA341GA; Bio-Rad, Hercules, CA), and vasculature was stained with von Willebrand factor (vWF; 1:100; ab287967; Abcam, Cambridge, MA). The basal lamina of muscle cross-sections was stained with laminin (1:200; PA1-16730 from Invitrogen, Waltham, MA). ECM deposition was determined using wheat germ agglutinin (WGA; 1:200; W11262; Invitrogen, Waltham, MA). Neuromuscular junctions (NMJs) were identified using α-bungarotoxin (1:50; B13422, Invitrogen, Waltham, MA) and synaptophysin (1:100; XE3573801, Invitrogen, Waltham, MA). Appropriate fluorochrome-conjugated secondary antibodies (1:100; A11034, A11001, or A11012; Invitrogen, Waltham, MA) were used as previously described. 12,43,44
To quantify Ku80, CD68, eMHC, and vWF stains, 3–5 images at 5x magnification were taken to capture most of the muscle cross-section using a ZEISS Axiocam microscope. Ku80+ and CD68+ images were thresholded via ImageJ to determine the percentage area. eMHC+ fibers were manually quantified. AngioTool software was utilized to quantify capillary density (vWF+) in histological images. 45 For histological analysis, only 1–2 contralateral control muscles per experimental group were tested and pooled together into a single group.
Muscle cross-sections stained for H&E, laminin, WGA, α-bungarotoxin (α-BTX), and synaptophysin (Syn) were scanned to obtain composite images using either Leica SP6 Epifluorescence or Olympus BX614S (Saint Louis University). In the H&E composite images, the freehand selection tool in ImageJ was used to determine the size of the BS in each muscle. A custom-designed image analysis MATLAB program was used to quantify the myofiber cross-sectional area (CSA) from laminin+ stained images. 38,40 For NMJ analysis, colocalized acetylcholine receptor (AchR) clusters (α-BTX+) and presynaptic nerve terminals (Syn+) were manually quantified. WGA-stained muscle sections were quantified using ImageJ. The BS region was excluded from analysis in samples where the BS appeared morphologically distinct from the muscle tissue. WGA has been previously used in many studies to quantify fibrotic tissue deposition in skeletal 46,47 and cardiac muscle. 48,49
Protein expression analysis
The bottom half of the TA muscle (containing the defect and the surrounding musculature) was utilized for protein isolation. The TA muscle (∼50 mg) was homogenized on ice in 600 μL of 1x radioimmunoprecipitation assay lysis buffer (20–188; Sigma-Aldrich, St. Louis, MO) containing protease inhibitor (PI) cocktail (1:100; Sigma-Aldrich, St. Louis, MO). The protein concentrations were determined by a Pierce bicinchoninic acid assay Protein Assay Kit (Thermo Scientific, Waltham, MA). For each sample, 60 μg of total protein per sample was loaded on 4–20% Tris-glycine gels (Bio-Rad, Hercules, CA). The protein was then resolved by running a sodium sulfate-polyacrylamide gel electrophoresis. The protein bands were transferred onto nitrocellulose membranes, stained with Ponceau S (TOCRIS Bioscience, Minneapolis, MN) to confirm equal loading, and blocked in 5% nonfat dry milk prior to probing for MyoD (1:500, MA1-41017, Invitrogen, Waltham, MA), desmin (1:500, ab15200; Abcam, Cambridge, MA), inducible nitric oxide synthase (iNOS; 1:500; ab49999; Abcam, Cambridge, MA), and arginase (1:500; ab91279; Abcam, Cambridge, MA) overnight at 4°C. Appropriate horseradish peroxidase-conjugated secondary antibodies (1:500; Invitrogen, Waltham, MA) were applied for 1 h at room temperature. Proteins were detected using Clarity Western enhanced chemiluminescence Substrate (Bio-Rad, Hercules, CA), and membranes were imaged using the ChemiDoc Imaging System (Bio-Rad). Band intensity quantification was performed in ImageJ and then normalized to Ponceau 50 for quantitative analysis. For Western blotting analysis, only 1–2 contralateral control muscles per experimental group were tested and pooled together into a single group.
Peak isometric torque measurement
In vivo function testing of the anterior crural muscles was performed 4 weeks postinjury. 51 The rats were anesthetized using isoflurane (1.5–2.0%) on a heated pad. The skin was shaved, and the animals were moved to a heated test apparatus (Model 806D, Aurora Scientific, Canada) to maintain normal body temperature. The knee was then clamped to secure the leg. After ensuring the leg was secured, the animal’s foot was positioned on the footplate of the dual-mode motor (Model 305C, Aurora Scientific, Canada). The heel was placed in the groove of the footplate, and adhesive tape was used to fasten the foot to the pedal. The foot was positioned perpendicularly to the tibia, forming a 90° angle. Two platinum electrodes were inserted subcutaneously on each side of the common peroneal nerve to stimulate the anterior crural muscles and elicit dorsiflexion. The optimal current amplitude (30–35 mA) was set with a series of twitches. Isometric tetanic contractions were elicited at 150 Hz (0.1 ms pulse width, 400 ms train). The data were acquired using the Dynamic Muscle Control/Dynamic Muscle Analysis software (Aurora Scientific, Canada). For torque measurement, only 2–3 contralateral control muscles per experimental group were tested and pooled together into a single group.
Statistical analysis
The data are presented as mean ± standard error of the mean, with significance set at p < 0.05. GraphPad Prism 10 for Windows was used to perform statistical analysis and data graphing. A one-way analysis of variance (ANOVA) with Fisher’s least significant difference post hoc test was performed to detect statistical significance. A two-way ANOVA was performed to analyze the fiber-size distribution and in vitro data collected on hypoxic and normoxic cultures.
RESULTS
BS encapsulated hPDSC culture
Confocal microscopy revealed that hPDSCs (DAPI+Qtracker+) were uniformly distributed throughout the BS structure and infiltrated to a depth of about 300 microns (Supplementary Fig. S1). On days 1–5 of culture, the hPDSCs remained viable within the BS, determined by their DNA content (Supplementary Fig. S1; Interaction: p = 0.6754), and continued to secrete HGF (Supplementary Fig. S1; ANOVA p = 0.0015, p = 0.0054, respectively), amphiregulin (Supplementary Fig. S1; ANOVA p = 0.0349), follistatin (Supplementary Fig. S1; ANOVA p = 0.0007), and IL-6 (Supplementary Fig. S1; ANOVA p = 0.0328).
BS encapsulated hPDSCs were cultured under normoxic (21% O2) and hypoxic (3% O2) conditions. Cells remained viable in both hypoxic and normoxic environments, and no differences in DNA content were observed between days 1–5 (Fig. 1A). H-PDSC-BS released higher HIF-1α on day 1 and 3 relative to N-PDSC-BS (Fig. 1B; Interaction p = 0.0447, time effect p = 0.4637, treatment effect p = 0.0437, post hoc p = 0.0221, p = 0.0393, respectively). The release of a pro-angiogenic factor, FGF-2, was significantly higher on day 3 in the hypoxic group (Fig. 1C; Interaction p = 0.2565, time effect p = 0.0677, treatment effect p = 0.0317, post hoc p = 0.0100). The hypoxic group released more VEGF on day 1 relative to the normoxic group (Fig. 1D; Interaction p = 0.0003, time effect p < 0.0001, treatment effect p = 0.0178, post hoc p < 0.0001). Interestingly, the H-PDSC-BS group suppressed the production of a myogenic marker, HGF (Fig. 1E; Interaction p = 0.0206, time effect p = 0.0165, treatment effect p = 0.0086, post hoc p = 0.0012), and follistatin on day 6 relative to the N-PDSC-BS (Fig. 1F; Interaction p = 0.5087, time effect p = 0.4337, treatment effect p = 0.0248; post hoc p = 0.0287). IL-6 showed no differences between experimental groups and time points (Fig. 1G). Based on these results, three days of hypoxic or normoxic conditioned hPDSC-BS were used for in vivo experiments.

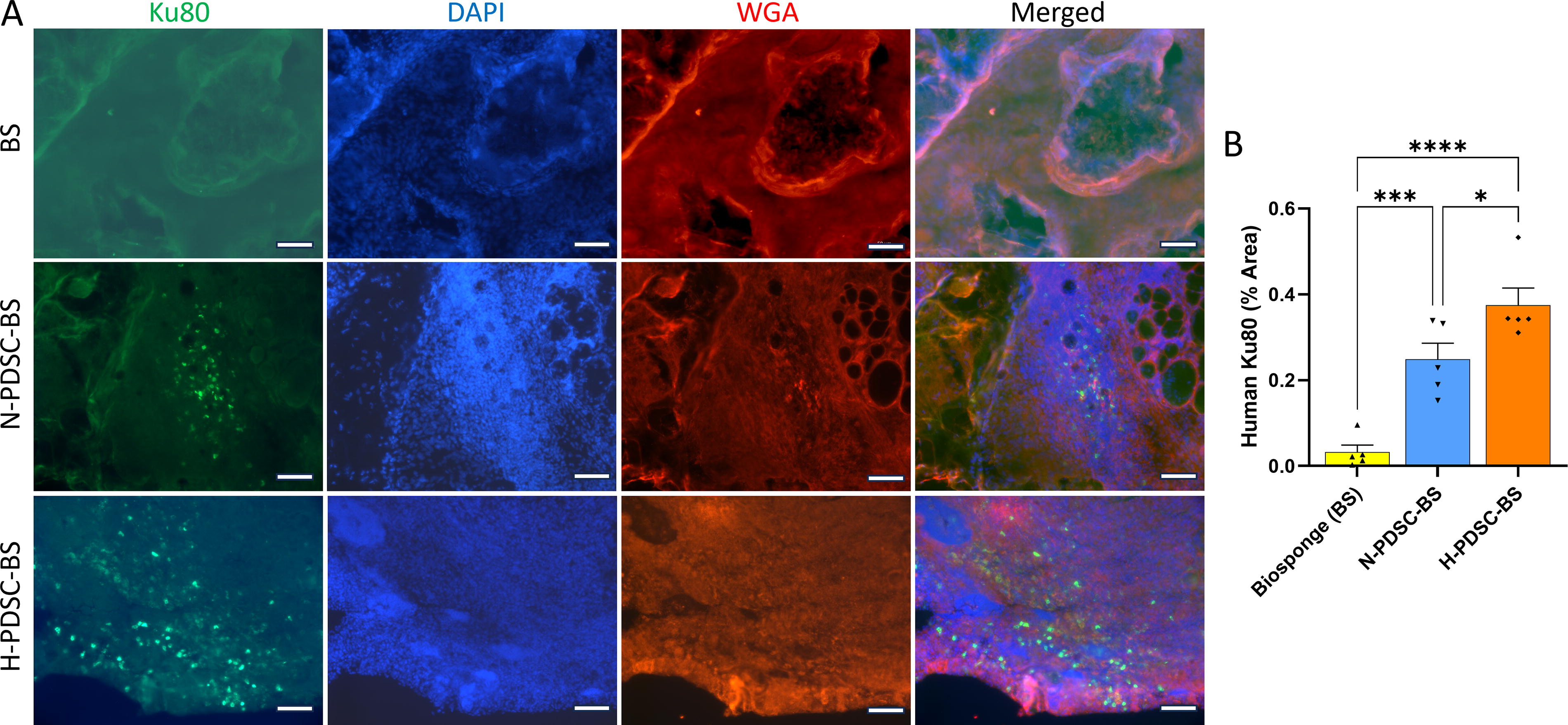
hPDSC biodistribution
The nuclei of the hPDSCs (human Ku80+) were visualized within muscle transverse cross-sections. Both N- and H-PDSC-BS groups showed the presence of human Ku80+ cells on days 7 (Supplementary Fig. S2) and 28 postinjury (Fig. 2A). As expected, the acellular BS did not show any positive staining for Ku80. The hPDSCs were confined to the defect area and the boundary of the BS and did not migrate into the remaining musculature (Supplementary Fig. S3). Quantifying the Ku80+ stain indicated that the hypoxic group retained more cells than the normoxic group (Fig. 2B; p = 0.0195; ANOVA p < 0.0001). Both cellular groups were significantly higher than the BS group (BS vs. N: p = 0.0006; BS vs. H: p < 0.0001). Qualitative assessment suggests that the percentage of Ku80+ hPDSCs was very low relative to the total number of DAPI+ nuclei in the defect region.
Muscle mass, function, and morphology
Muscle cross-sections stained with H&E showed extensive cellular infiltration into the VML defect (Fig. 3A). The BS could be identified histologically, and its area was quantified in the treated groups. The acellular BS appeared fragmented and degraded with extensive cellular infiltration. In contrast, the BS in both hPDSC groups appeared largely intact, with minimal cellular infiltration inside its porous structure. The BS area quantified was larger in the hPDSC groups relative to the acellular group (Fig. 3B; ANOVA p = 0.0037). Muscle cross-sections were also stained with eMHC (Fig. 3C), and quantification revealed that the experimental groups had similar counts of eMHC+ regenerating fibers (Fig. 3D; ANOVA p = 0.0697). For the BS and the NR groups, the eMHC+ myofibers were found within the defect region. In the N- and H- groups, the eMHC+ myofibers were observed on the edge of the defect and at the interface of the BS and the remaining muscle mass.

There were no differences between experimental groups with respect to biopsy mass that was removed to create the VML injury (Fig. 4A; ANOVA p = 0.9163). At 28 days postinjury, the body weight of animals was recorded (Supplementary Fig. S4; ANOVA p = 0.5234). The TA muscle mass normalized to body weight at 28 days postinjury was significantly reduced in the VML-affected limbs relative to the contralateral (CON; C) limbs (Fig. 4B; ANOVA p < 0.0001). The TA muscle mass in the BS (p = 0.0195) and H-PDSC-BS (p = 0.0333) treated groups was higher than the NR group. Analysis of the raw TA muscle mass (Supplementary Fig. S4; ANOVA p < 0.0001) revealed that only the H-PDSC-BS had higher muscle mass than the NR group (p = 0.0341).
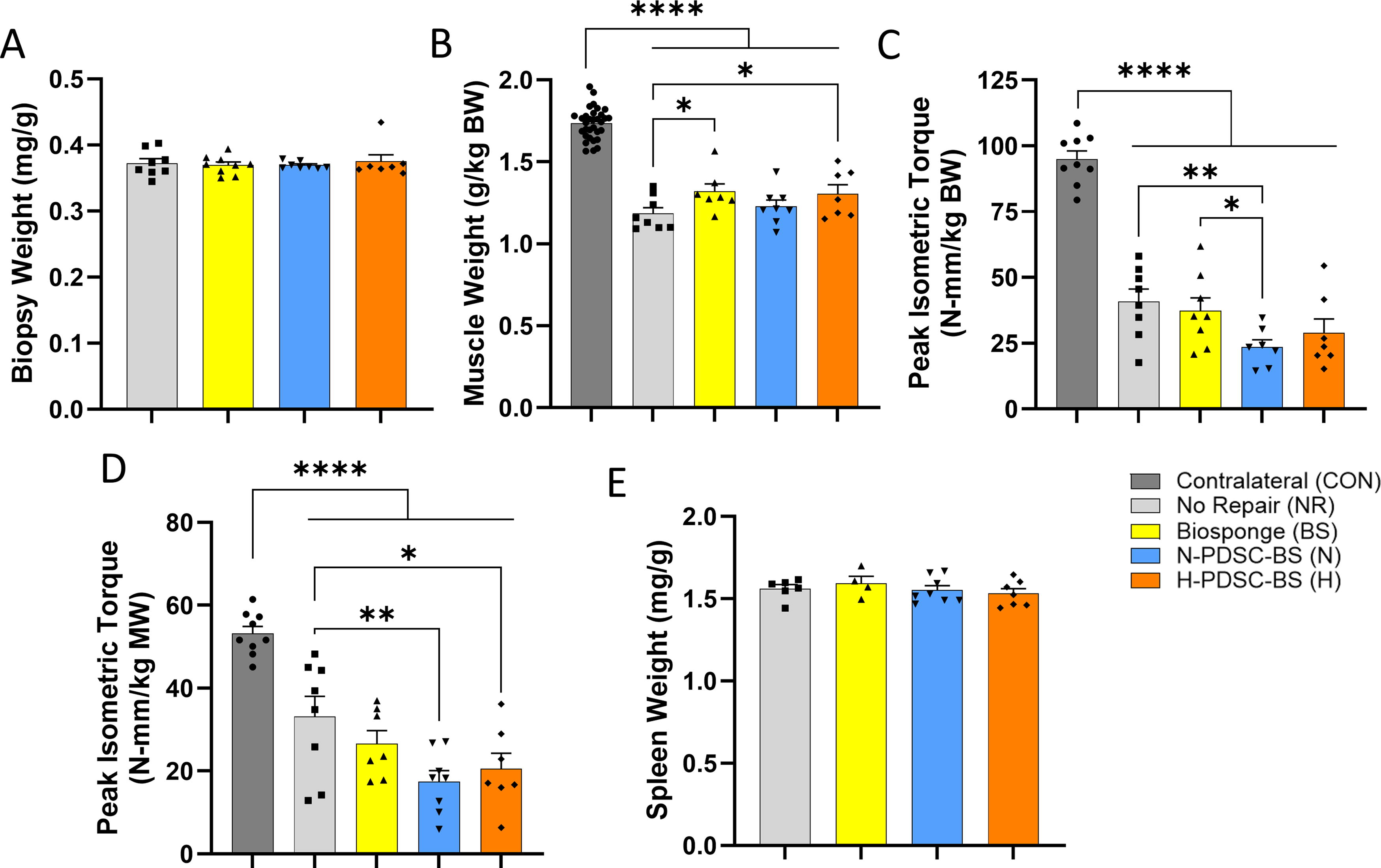
The peak isometric torque normalized to body weight was significantly lower in the VML-affected limbs compared with the CON limbs at 28 days postinjury (Fig. 4C; ANOVA p < 0.0001). No functional improvements were observed in the treated groups relative to the NR group. The N-PDSC-BS group had lower peak isometric torque than the NR (p = 0.0084) and BS (p = 0.0319) groups. The raw peak isometric torque data (Supplementary Fig. S4; ANOVA p < 0.0001) also indicate the N-PDSC-BS having lower torque than the NR (p = 0.0210). The peak isometric torque was also normalized to muscle mass (Fig. 4D; ANOVA p < 0.0001), and results showed that the NR group had higher torque than both N- (p = 0.0019) and H-PDSC-BS groups (p = 0.0139). Spleen weight was similar between groups, indicating no adverse systemic inflammatory reactions (Fig. 4D; ANOVA p = 0.6254).
Myofiber cross-sectional area
Laminin-stained muscle cross-sections used for CSA analysis are shown in Figure 5A. The myofiber fiber size distribution was then investigated (Fig. 5B; Interaction p < 0.0001). The hPDSC groups had higher percentages of small diameter myofibers (<500 µm2) than the other groups (H vs. BS: p = 0.0035; N; H vs. NR; C: p < 0.0001). N-PDSC-BS had significantly higher small myofibers than the H-PDSC-BS group (p = 0.0347). The NR (p = 0.0411), N- (p = 0.0012), and H-PDSC-BS (p = 0.0170) groups had higher percentages of 500–999 µm2 fibers relative to the CON group.
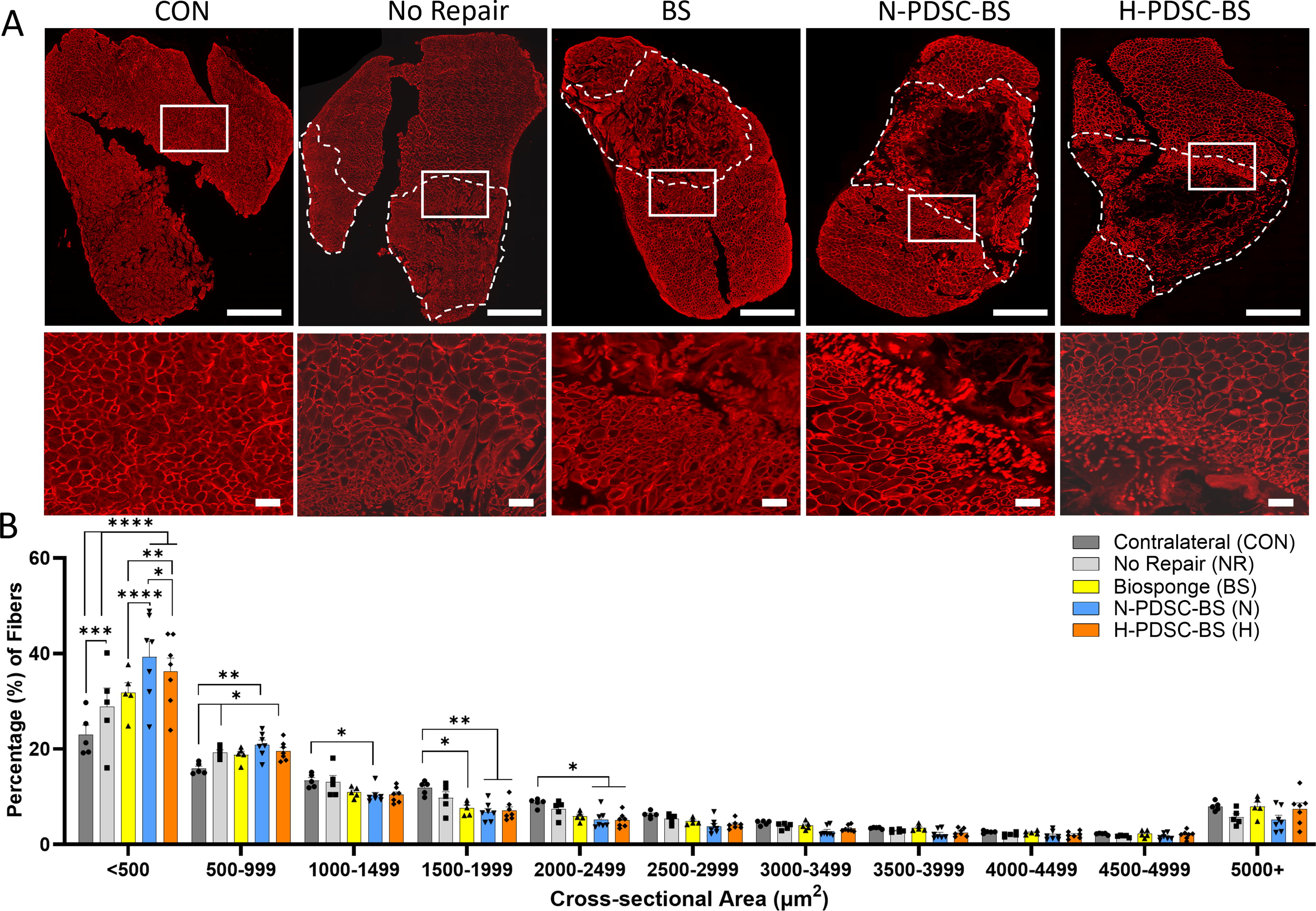
Mean CSA quantification determined that the CON (p = 0.0017) and BS (p = 0.0308) groups had higher mean CSA than the N-PDSC-BS group (Supplementary Fig. S4; ANOVA p = 0.0243). The CON group also had a higher mean than the H-PDSC-BS group (p = 0.0308). The CON group had significantly higher amounts of fibers than the other groups (C vs. NR: p = 0.0095; C vs. BS;N;H: p < 0.0001). The NR group had a greater number of myofibers (Supplementary Fig. S1; ANOVA p < 0.0001) relative to the treatment groups (NR vs. BS: p = 0.0290; NR vs. N: p = 0.0024; NR vs. H p = 0.0042).
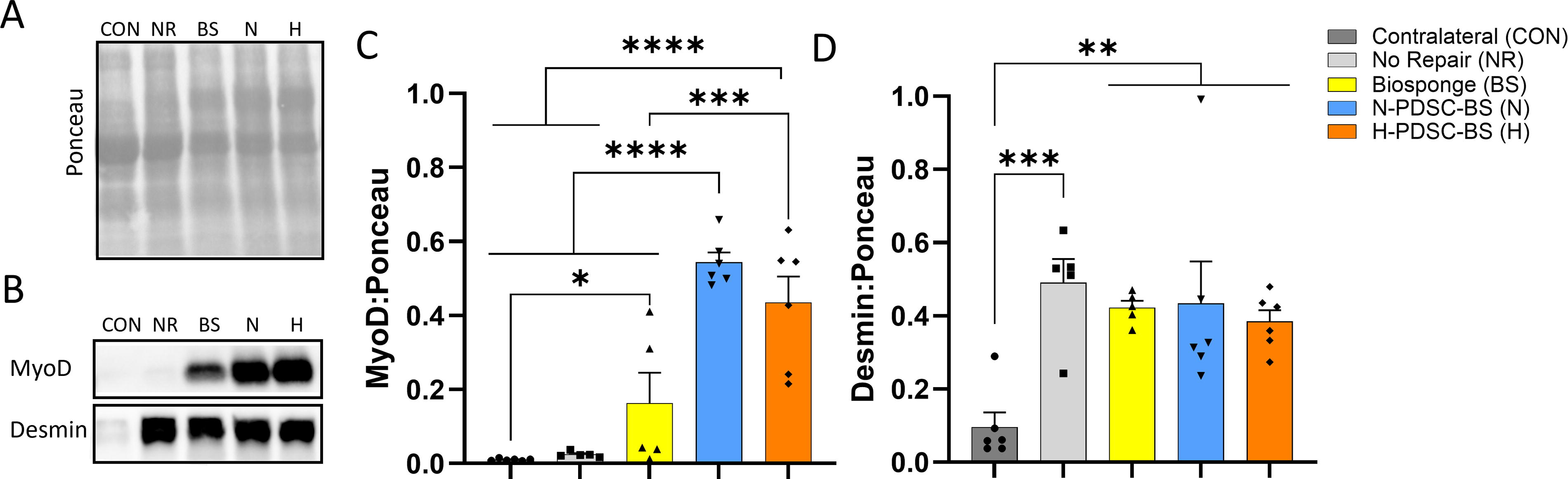
Myogenic response
Protein expression of myogenic markers, MyoD and desmin, was analyzed (Fig. 6A–B). Both N- and H-PDSC-BS groups had higher MyoD expression than the CON (N;H: p < 0.0001), NR (N;H: p < 0.0001), and BS (N p < 0.0001; H p = 0.0007) groups (Fig. 6C; ANOVA p < 0.0001). BS group expressed higher amounts of MyoD than the CON group (p = 0.0395). Desmin expression (Fig. 6D; ANOVA p = 0.0025) was similar among injured groups but significantly higher than the CON group.
Host response
Macrophage (CD68+) infiltration was observed (Fig. 7A) using immunofluorescence staining. Qualitative analysis indicates that the acellular BS had more macrophage (CD68+) infiltration than the N- and H-PDSC-BS groups. However, quantitative analysis revealed that the total percentage area occupied by macrophages (CD68+) in muscle cross-sections was similar between injured groups (Fig. 7B; ANOVA p = 0.0015). As expected, the CON group showed a lower presence of macrophages (C vs. NR: p = 0.0003; BS: p = 0.0004; N: p = 0.0044; H: p = 0.0009).
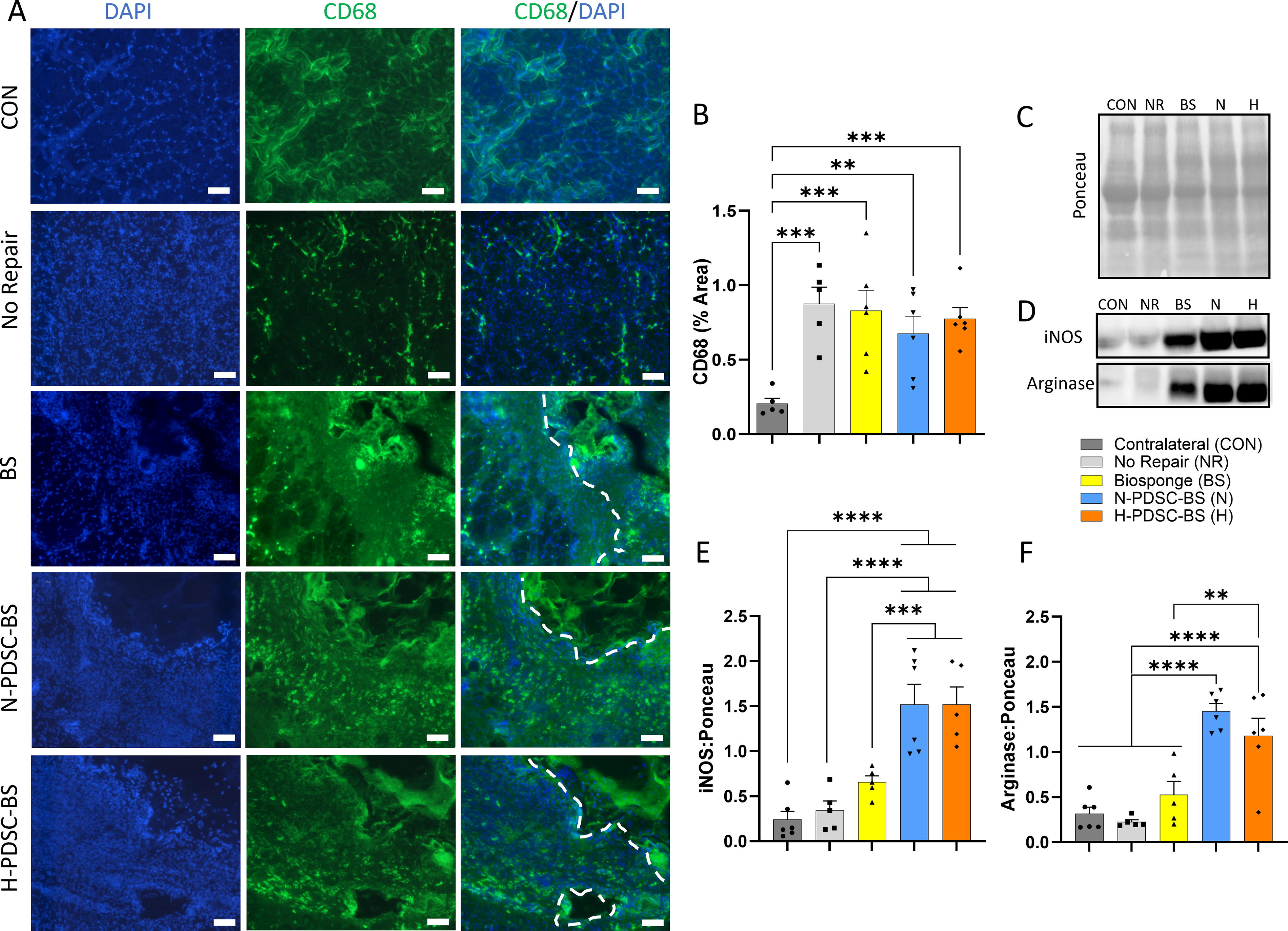
Protein expression of iNOS and arginase, associated with pro- (M1) and anti- (M2) inflammatory macrophages, respectively, was analyzed (Fig. 7C–D). Both N- and H-PDSC-BS groups had higher iNOS expression than the CON (N;H: p < 0.0001), NR (N;H: p < 0.0001), and BS (N: p = 0.0006; H: p = 0.0009) groups (Fig. 7E; ANOVA p < 0.0001). Arginase expression (Fig. 7F; ANOVA p < 0.0001) was also elevated in both hPDSC groups relative to the other groups (N;H vs. C;NR: p < 0.0001; N vs. BS: p < 0.0001; H vs. BS: p = 0.0011).
Capillary density
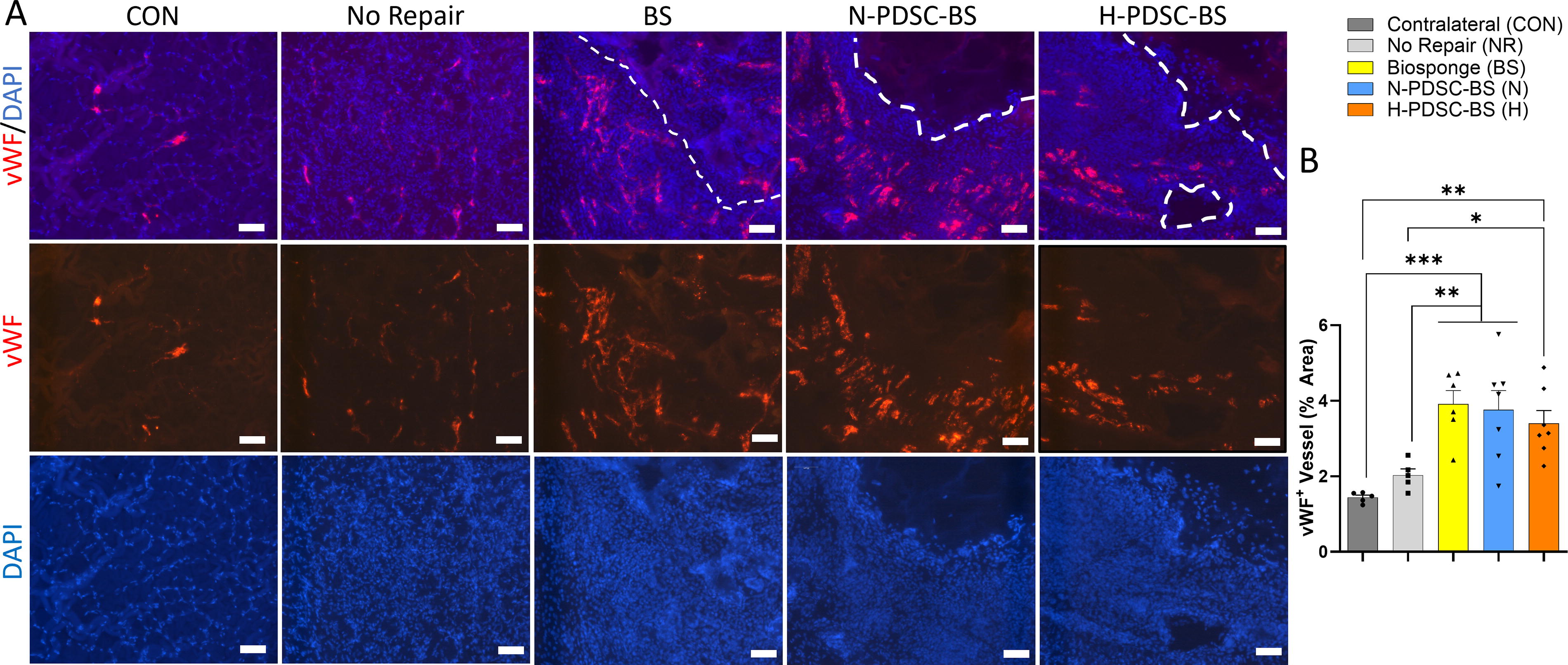
Muscle cross-sections were stained with vWF to identify capillaries (Fig. 8A). Quantitative analysis of vWF+ vessel percentage area (Fig. 8B; ANOVA p = 0.0002) showed that all groups containing BS had higher capillary density than the CON (C versus BS: p = 0.0001; N: p = 0.0002; H: p = 0.0010) and NR group (NR versus BS: p = 0.0020; N: p = 0.0031; H: p = 0.0155).
Fibrotic response
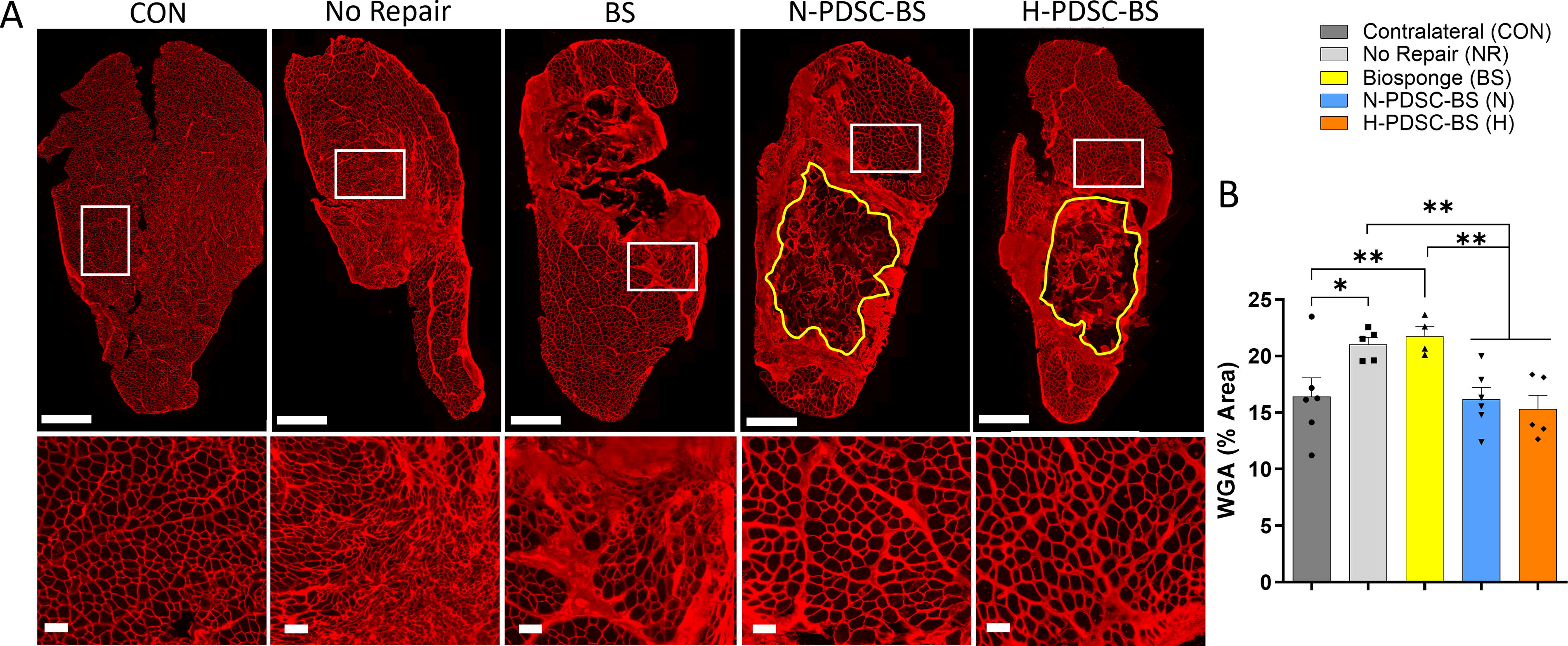
To determine the extent of ECM deposition in injured muscles, the percentage area occupied by WGA was quantified (Fig. 9A). WGA density was lower in hPDSC groups relative to the NR (NR vs. N: p = 0.0086; H: p = 0.0038) and BS (BS vs. N: p = 0.0051; H: p = 0.0024) groups (Fig. 9B; ANOVA p = 0.0031). The NR (p = 0.0120) and BS (p = 0.0070) group had significantly higher WGA density than the CON group.
Neuromuscular junction
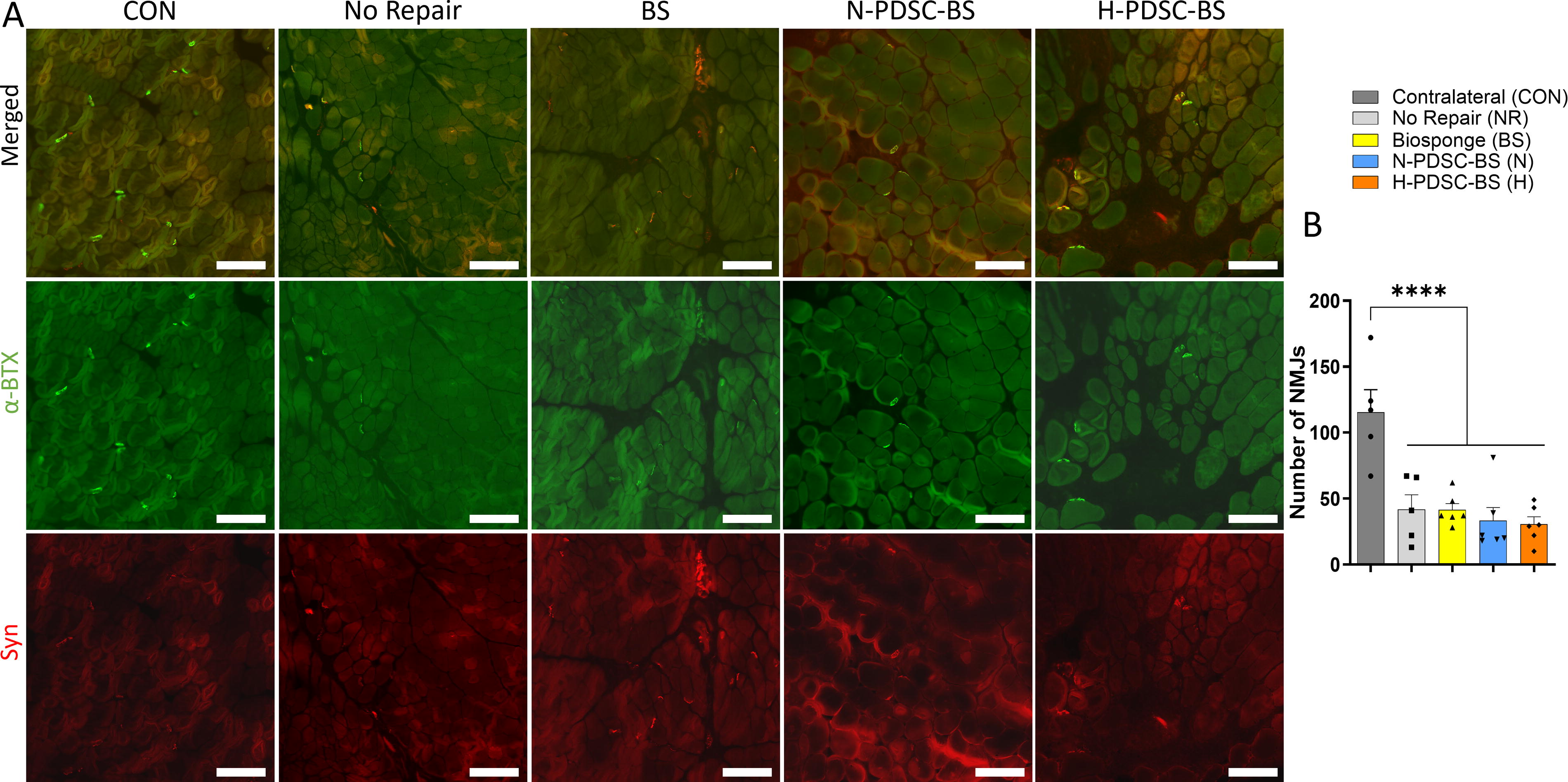
We stained the AchRs on the motor endplate with α-BTX and utilized antibodies against synaptic vesicles (Syn+) to visualize the neuron’s terminal bouton 52 (Fig. 10A). We quantified the AchRs with colocalized synaptophysin, which indicated the presence of an NMJ. The total number of NMJs was significantly higher in the CON group relative to the injured groups (C versus NR; BS;N;H: p < 0.0001). Among the injured groups, the total number of NMJs was similar (Fig. 10B; ANOVA p < 0.0001).
The peak isometric torque significantly correlated with muscle mass, myofiber CSA, total number of myofibers, and NMJs, emphasizing the interdependence of muscle structure and function (Supplementary Fig. S5).
DISCUSSION
The most salient finding of our study is that treatment with BS encapsulated hPDSCs enhanced myogenesis and suppressed fibrotic tissue accumulation in VML injured muscles relative to the No Repair group. Improved myogenesis is evidenced by increased protein expression of an early-stage myogenic marker, MyoD, in VML-injured muscle. The encapsulation of hPDSCs also slowed the remodeling and degradation of the BS scaffolds in the VML defect. However, despite these improvements, an increase in total or regenerating myofibers (eMHC+) and peak isometric torque was not observed. The lack of functional improvement could be attributed to the persistent presence of an intact BS and insufficient contractile tissue deposition. Other contributing factors may also include deficits in reinnervation and immunomodulation. In addition, our findings indicate that the persistent presence of BS likely contributed to enhanced muscle mass observed in the study.
hPDSCs have been evaluated as a treatment for multiple muscle conditions in animal models, such as Duchenne muscular dystrophy 53 and hindlimb ischemia. 54,55 Recent research indicates that the regenerative effects of hPDSCs primarily result from their paracrine activity rather than direct participation in forming new muscle fibers. hPDSCs secrete a diverse array of trophic factors that can promote immunomodulation 56 and vascularization, 55,57 minimize fibrosis, 53,58 and enhance signaling to native cells. In fact, a study showed that live hPDSC transplantation was more effective than an injection of their cellular lysate in restoring blood flow, highlighting the significance of prolonged trophic factor release in repairing injured tissues. 54 The same study also showed that blood blow restoration can be achieved whether cells are injected into the injured limb or the contralateral control limb, suggesting that the repair mechanism is likely to be paracrine in nature.
Our in vitro studies established that BS encapsulated hPDSCs can maintain their viability and functionality in both hypoxic and normoxic culture conditions. hPDSCs encapsulated within BS released follistatin, IL-6, HGF, and amphiregulin. These factors play multifaceted roles in muscle regeneration. For instance, follistatin is a regulatory protein that can block myostatin, a factor that limits muscle growth. 59,60 Transgenic mice overexpressing follistatin in skeletal muscle show increased muscle mass. 61 IL-6, a pro-inflammatory cytokine, is predominantly produced in actively contracting muscle fibers after increased workload. 62 –65 It serves as a chemoattractant for myoblasts and myeloid cells, supporting myoblast proliferation. 66 IL-6 deficiency blunts hypertrophic growth in mice under mechanical overload. 64 HGF can activate quiescent satellite cells and drive them into cell cycle in vitro and in vivo. 67,68 HGF overexpression in skeletal muscle is shown to facilitate muscle regeneration not only by stimulating satellite cell activity but also by enhancing angiogenesis 69 and polarizing macrophages into an anti-inflammatory M2 phenotype. 70 Amphiregulin, a growth factor that signals through the epithelial growth factor receptor, promotes myogenic differentiation in satellite cell cultures. 71 The administration of amphiregulin reduces pro-fibrotic gene expression in injured skeletal muscle 71 and alleviates muscle damage and inflammation caused by Toxoplasma gondii infection. 72
Studies have also demonstrated that hypoxic culture can enhance trophic factor release by hPDSCs. 57,73 Several studies support the fact that cellular response to hypoxia is mediated by the activation of the hypoxia-sensitive transcription factor HIF-1α. 74 Our in vitro studies revealed elevated release of pro-angiogenic growth factors such as HIF-1α, VEGF, and FGF2 on day 3 of hypoxic culture from BS encapsulated hPDSCs. However, by day 6 of hypoxic culture the release of some myogenic growth factors was suppressed. These findings indicate that brief exposure to hypoxia may enhance the release of trophic factors, whereas prolonged exposure could have detrimental effects. Based on these results, we chose to culture the hPDSCs within the BS for 3 days before transplantation. By selecting this time point, we aimed to maximize trophic factor release by the hPDSCs.
We also assessed the effect of hypoxic preconditioning on cellular persistence and retention in the hostile microenvironment of VML injured muscle. A human Ku80 stain of muscle cross-sections visualized the human DNA of hPDSCs within the rodent muscle. 42 While the percentage of human Ku80+ cells was low, quantitative analysis showed higher hPDSC presence in the hypoxic preconditioned group at day 28 postinjury. In support, previous studies have also shown that hypoxic preconditioning can promote higher cell retention after transplantation. 31,32 Furthermore, qualitative analysis of the Ku80+ cells suggested that the cells did not migrate away from the defect site into the remaining muscle mass. Furthermore, we could not identify any myofibers with Ku80+ staining, indicating that hPDSCs did not directly contribute to myofiber regeneration. These results are consistent with previous studies demonstrating that transplanted MSCs do not directly differentiate into myofibers. 75 –77 Previous work also indicates that when injected locally into muscles 53, 55 or tendons, 78 these cells can persist in vivo for 3–4 weeks, with levels gradually decreasing over time. In a rat model of tendinopathy, hPDSC administration improved tendon biomechanical properties at 2 weeks but not at 4 weeks postinjury due to diminishing cell presence. In this work, we did not detect any statistical differences between the N-PDSC-BS and the H-PDSC-BS in terms of muscle mass, function, or myofiber size. However, the H-PDSC-BS group performed slightly better than the N-PDSC-BS group in improving muscle mass relative to the NR group while maintaining peak isometric torque and mean myofiber CSA.
Studies have shown that hPDSCs can successfully engraft in mouse 53,55,57 or rat 58,78 tissues without adverse events 79 and that cells were confined to the injection site with little to no proliferation or migration into adjacent or distant tissues. 55,79 To determine the hPDSC effects on immunomodulation in VML-injured muscles, we quantified macrophage infiltration and phenotypic polarization. At 28 days postinjury, we noted comparable macrophage (CD68+) presence in the injured groups. The protein analysis of macrophage phenotype markers uncovered that BS encapsulated hPDSCs increased both M1-like iNOS and M2-like arginase protein expression in injured muscles. Studies have shown that a complex interplay between both pro- and anti-inflammatory macrophages is needed for successful tissue recovery of VML injured muscles. 12 Pro-inflammatory macrophages support migration and activation of myogenic precursor cells, reduce fibrosis, 80 and promote scaffold remodeling, 81 whereas an anti-inflammatory response fosters the differentiation of the muscle precursor cells and formation of myotubes. 66 These results suggest that BS encapsulated hPDSC group elicits a mixed macrophage phenotype response and amplifies both M1 and M2 activity in the injured muscle. In support, previous studies have shown that PDSCs can be immunogenic 82 and can release cytokines that can recruit and activate immune cells. 83
Excessive noncontractile tissue deposition is common in VML injuries. This excessive noncontractile tissue deposition or fibrosis promotes rapid repair of damaged tissue but compromises functional tissue regeneration. This can hinder regeneration, hypertrophy, and force production. 13,14,84,85 BS encapsulated hPDSCs diminished the ECM accumulation in the VML-injured muscle relative to other groups. In support, transplantation of hPDSCs resulted in reduced fibrosis in animal models of liver 58 and intestinal 86 fibrosis. These studies have attributed these effects to the antifibrotic agents in the secretome of hPDSCs, including HGF, 87 amphiregulin, 71 and follistatin. 88
Histological analysis of vWF+ vessels was performed to determine the extent of vascularization. The angiogenic response was higher in all BS groups, and the incorporation of hPDSCs did not offer any additional benefit by day 28 postinjury. These findings were unexpected, given the higher levels of angiogenic growth factor production by hypoxic preconditioned hPDSCs. This result could be explained by the diminished cellular presence on day 28 postinjury, which likely reduced the production of pro-angiogenic growth factors at the VML site.
The peak isometric torque at day 28 postinjury was similar between the NR and BS groups, with a ∼57% and ∼61% deficit relative to the CON control group. However, the deficits in the N-PDSC-BS and the H-PDSC-BS groups were higher at ∼75% and ∼70% relative to the control group. The higher deficit in torque production could be attributed to the largely intact BS scaffold in hPDSC groups, which might impede force transmission to the tendon and contractile tissue deposition in the defect. The BS scaffold must undergo substantial remodeling and degradation before any considerable myofiber deposition can occur in the defect area. 89 Previous studies have shown that MSCs can deposit extensive amounts of ECM within bioscaffolds and enhance their mechanical properties. 90 –92 We speculate that increased deposition of cell-secreted ECM may have mechanically strengthened the BS and slowed its remodeling and degradation in the VML injured muscle. Future investigations will examine functional recovery at later times when the scaffold has been completely resorbed and replaced by native tissue. In support, another study with MSC seeded bioscaffold repair of VML injury showed deficits in functional recovery at 4 weeks postinjury, but at 12 weeks postinjury, significantly higher recovery in function was observed. 93 In follow-up studies, the mechanical and degradation properties of the BS can be modified by adjusting the concentration of ECM proteins and cross-linkers.
It should be noted that peak isometric torque correlated significantly with muscle mass, myofiber CSA, total number of myofibers, and the number of NMJs in this study (Supplementary Fig. S5). This highlights the interconnectedness of these measurements, indicating their reliability and mutual reinforcement in data interpretation. In addition, this correlation sheds light on the key contributors to force production, elucidating the complex interplay between muscle structure and function.
Intramuscular neural damage from VML disrupts myofiber innervation. 94,95 VML injury resulted in significant deficits in NMJ quantity relative to the contralateral control muscles. However, we did not observe any differences among the experimental groups in NMJ quantity. A limitation of the study is that muscle tissue was only preserved for histological cross-sections where only a two-dimensional analysis could proceed. We could not investigate structural changes in NMJ. Future studies will explore the extent of morphological remodeling in NMJs in longitudinal sections.
Another limitation of this study is that it only tested a single cell dose. A study examined the dose-dependent effects of hPDSCs in restoring blood flow. Interestingly, intramuscular injection of 0.05, 0.1, and 0.2 million cells per mouse resulted in similar improvements in blood flow rate. The minimal effective dose was identified as 10,000 cells per animal in the study, but all tested dosages were found safe. 54 Future studies will explore the dose-dependent effects of hPDSCs in improving muscle regeneration post-VML injury. In addition, only a single time point was chosen for analysis. Future studies will evaluate the regenerative effects of hPDSC administration at 2 weeks postinjury. Another potential area for future investigation is the role of administering additional injections of hPDSCs for sustained therapeutic effects.
CONCLUSION
Overall, our results suggest that treatment with BS encapsulated hPDSCs enhances myogenesis while simultaneously reducing fibrosis and BS remodeling rate in a rat model of VML. These results are encouraging and suggest that hPDSCs are a promising therapeutic option for VML repair that is worthy of further investigation.
KEY FINDINGS
Biosponge encapsulated hPDSCs Enhance myogenesis (based on increased MyoD protein expression) Reduce fibrosis and biosponge remodeling rate Did not increase muscle function, likely due to persistent presence of an intact biosponge and deficits in innervation Biosponges with or without hPDSCs increased capillary density.
Footnotes
ACKNOWLEDGMENTS AND FUNDING SOURCES
The authors thank Jessica Robinson and Saloni Shringarpure for help with BS fabrication and immunofluorescence stain assistance when needed. The authors also thank Caroline Murphy at Kolar Lab histological core for slide scanning assistance and training. This work was supported by the Center of Defense Medicine at
AUTHORS’ CONTRIBUTIONS
K.G. and K.B. designed the study. D.J. performed the animal experiments, collected data, performed statistical analysis, and prepared figures. M.R., R.M., M.C., J.B., J.T., and A.J. also performed experiments and collected data. D.J. and K.G. drafted the article. D.J., K.G., and K.B. revised and finalized the article. All authors approved the final version of the article.
AUTHOR DISCLOSURE AND GHOSTWRITING
GenAssist, Inc. is developing products related to the research described in this article. K.G. has an equity interest in GenAssist, Inc., and serves on the company’s scientific advisory board. The terms of this arrangement have been reviewed and approved by Saint Louis University, in accordance with its conflict-of-interest policies. The authors declare no other competing financial or nonfinancial interests. The content of this article was written by the authors listed. No ghostwriters were used to write this article.
ABOUT THE AUTHORS
D.J. is a 4th year PhD. candidate at Saint Louis University (SLU) in Dr. Koyal Garg’s Musculoskeletal Tissue Engineering lab. M.R., R.M., M.C., and J.B. are undergraduate students, and J.T. and A.J. are PhD. students at SLU in the same laboratory. K.B. is the collaborating PI at Wake Forest Institute of Regenerative Medicine (WFIRM). K.G. is the PI and 7th year Associate Professor at SLU. As a prominent researcher in the field of VML, K.G.’s research interests include stem cells and extracellular matrix-based biomaterials to improve the regenerative and functional capacity of skeletal muscle following trauma.
SUPPLEMENTARY MATERIAL
Supplementary Figure S1
Supplementary Figure S2
Supplementary Figure S3
Supplementary Figure S4
Supplementary Figure S5
Abbreviations and Acronyms
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.