
Abstract
Objective:
Fat grafting is widely applied for various purposes, including volume restoration, improving tissue quality, and promoting wound healing, but it has poor long-term graft survival predictability. Alpha-1-antitrypsin (AAT) administration is hypothesized to improve fat graft outcomes by expediting inflammatory resolution and graft vascularity and reducing necrosis.
Approach:
Mice heterozygote to human AAT was grafted fat under the scalp alongside 400 µg/graft AAT or albumin (ALB) on days 0 and 3. Graft volume was determined by micro-magnetic resonance imaging, and explants were assessed for viability, histology, immunohistochemistry, and expression of selected genes. AAT expression was examined in hypoxia-exposed adipose-derived stem cells (ADSCs).
Results:
After 90 days, AAT-treated grafts maintained higher volumes (70.06% vs. 34.54%, n = 8, p = 0.02) and displayed improved tissue quality. On day 10 after grafting, grafts exhibited more blood vessels (mean 1.94/mm2 vs. 0.33/mm2) and 6.25-fold more adiponectin transcript levels (n = 12, p = 0.02). Although day-3 interleukin (IL)-1β expression was 5-fold greater in AAT-treated grafts (n = 6, p = 0.4), day-10 IL-1β expression was 2-fold lower compared to ALB-treated grafts (n = 22, p = 0.01). In the Methoxynitrosulfophenyl-tetrazolium carboxanilide (XTT) assay, day-3 AAT-treated grafts were 1.56-fold more metabolically functional (n = 6, p = 0.04) and exhibited greater perilipin-positive regions (18.5% versus 3.1%). Hypoxia-exposed ADSC expressed 9-fold higher AAT transcript levels (p = 0.04).
Innovation:
Fat grafting outcomes improved by early AAT treatment, probably by accelerating inflammatory resolution. Due to its marked safety profile, the study’s findings are for adjunct clinical-grade AAT therapy.
Conclusion:
AAT has a promising potential to be utilized as a fat graft outcome enhancer in terms of volume retention predictability and tissue quality.
Dor Halpern

INTRODUCTION
Autologous fat grafting is widely applied in reconstructive and aesthetic surgery performed in various anatomical sites for volume restoration or enhancement, improving the quality of the overlaying tissue, and treating difficult-to-heal wounds.1–3
Every aspect of the multistep fat grafting procedure, from fat collection, graft processing, and graft injection, might affect outcomes.4,5 Factors contributing to fat graft survival include the capacity of the vascular bed in the recipient area to supply the graft and the proportion of passenger adipose-derived stem cells (ADSCs).6,7 ADSCs are crucial for fat grafting outcomes since, unlike mature adipocytes that expire within 24 h, 8 ADSCs survive without direct blood supply for 72 h. 9 In addition, ADSCs protect adipocytes from inflammatory injury. 10 Adiponectin gene expression is low in fibrotic and inflamed fat tissue, whereas collagen-I gene expression is elevated in fibrosis. This distinction may serve to identify a graft that contains intact adipocytes and is less fibrotic.11,12
Many fat graft enrichment techniques have been introduced to improve fat graft retention rates, including ADSC, stromal vascular fraction (SVF), and platelet-rich plasma.13–16
Alpha-1-antitrypsin (AAT) is a serine protease inhibitor with multiple biochemical activities that is induced by inflammatory and hypoxic signals; circulating levels of AAT rise during acute phase responses, as well as during healthy pregnancy, as reviewed elsewhere. 17 In conditions rich in AAT, cell survival is improved, tissue repair is enhanced, bacterial burden is reduced, and immune responses are modulated towards tissue tolerance. 17 AAT supports tissue-repair—associated M2-like macrophages low in inducible nitric oxide synthase (iNOS) and high in arginase-1 transcripts, 18 a phenotype associated with improved long-term volume retention, as well as improved graft vascularization. 19 AAT also protects endothelial cells from injury 20 and promotes revascularization during ischemic conditions. 21 Interestingly, ADSCs produce AAT under inflammatory conditions. 22 Patients with genetic AAT deficiency receive life-long weekly infusions of plasma-derived AAT to restore circulating AAT to a normal range. 17 Clinical-grade AAT has a remarkable safety profile, 23 and is presently evaluated as a tissue-protective agent for tissue repair during myocardial infarction and cerebrovascular injury, as well as for organ preconditioning for transplantation24–26 and for improving outcomes of nonvascularized cell transplants.27,28
Clinical problem addressed
There is currently no consensus regarding fat graft enrichment, and the resulting fat graft volume retention rates vary between 10% and 90%; in addition, grafts display unpredictable surface irregularities, fibrosis, and calcifications, 29 all representing significant hurdles towards the desired outcomes in the context of reconstructive and aesthetic surgery. 30 We hypothesized that introduction of AAT into fat grafts will improve graft retention rates and desired tissue profile.
MATERIALS AND METHODS
An electronic laboratory notebook was not used
Animals
The study focused on therapeutic translatability by examining the applicability of a clinically safe formulation of AAT using a straightforward animal model for fat graft transplantation. Experimental protocols were approved by the Institutional Animal Care and Use Committee of Ben Gurion University (authorization No. IL-09-03-2023D), and follows the ARRIVE guidelines (Animals in Research: Reporting In Vivo Experiments). 31 Heterozygote mice transgenic for human AAT on a C57BL/6 background were generated by breeding homozygote transgenic mice with wild-type C57BL/6 mice (Envigo, Israel) and routinely genotyped as described elsewhere;32,33 heterozygote mice were used so as to allow treatment with human AAT without mounting antihuman murine antibodies. Eight-week-old females were used as both fat donors and fat graft recipients so as to allow subsequent follow-up as the mice mature, and minimize changes that might arise from sex and age differences. Mice were kept in standard care conditions at the animal facility, that is, 25°C ambient temperature, 2–4 animals per cage, with constant replacement of bedding, food and water. To reduce pain after procedures, analgesia was provided in drinking water (Dipyrone, 15 mg/kg).
Syngeneic fat graft mouse model
Fat grafts were prepared as described elsewhere with minor modifications.6,34 Briefly, as illustrated in Figure 1, mice were anesthetized by 2.5% isoflurane inhalation, and fat tissue was excised from the inguinal fat pads. The excised tissue was immediately placed on a sterile dish on ice and minced into <1 mm3 pieces using sterile scissors. The minced tissue was transferred to perpendicular 3 cc syringes and allowed to gravitate downwards towards the tip. When there was a visible separation of the top oily layer, middle adipose tissue layer, and lower SVF, the oily layer was absorbed using sterile gauze. Then, the stromal vascular and adipose tissue fractions were mixed for immediate injection into the recipient site. Fat was injected underneath the scalp beneath the subcutaneous layer, above the pericranium through an 18 G needle (300 µL), followed by placing a 5–0 absorbable suture to prevent leakage. Based on preliminary dose-response studies (unpublished data), 400 µg/graft of either human AAT (Glassia®, Kamada, Israel) or human serum-purified albumin (ALB) (ENCO, Israel) were introduced to the graft site in a nonblinded manner (n = 50). Another dose of AAT/ALB was injected subcutaneously to grafts 72 h after grafting. Under anesthesia with 2.5% inhaled isoflurane, mice were placed in a micro-MRI apparatus (Aspect M7, 1Tesla) set at high-resolution, T2 sequence, head-only, sagittal plane. Photos of the MRI sequences per mouse were acquired on days 3, 10, 30, 45, 60, and 90 from grafting and assessed for signs of necrosis and formation of a capsule around the graft; graft volume was analyzed using the VivoQuant program (ver.2.5, Invicro, MA). Two independent examiners visually demarcated the fat grafts. On days 1, 3, 10, 60, and 90 after grafting, randomly assigned graft recipients were anesthetized and euthanized and their grafts were extracted. Isolated fat grafts were immediately photographed next to a measuring scale, then fixed in formalin (Sigma-Aldrich) for histology and immunohistochemistry or RNA save (BI, Israel) for gene expression analysis.

Murine syngeneic fat graft model. A schematic diagram depicting the stages of the used model for fat grafting. (
Histology
Paraffin-embedded graft explants were cut in a microtome, and slides were stained with Hematoxylin and Eosin (H&E) following standard protocols. Tissue was assessed by a digital microscopy expert under optical magnification ×40 or ×200 for overall fatty tissue structure and adipocyte viability, inflammatory cell infiltration, blood vessels, and necrosis, using the following methods:
Image acquisition and analysis
Images were obtained by Panoramic Midi II slide scanner (Histotech, Hungary). Slide images were processed by QuPath v.0.4.2 software by a custom-made pipeline. The thresholding algorithm extracted tissue regions with pixel size 2 μm and threshold level 250. Extracted regions were further segmented into adipocytes, stroma, and vessels. Adipocytes and stroma were distinguished according to the automatically reconstructed eosin channel intensity using a threshold algorithm. Regions of interest (ROI) with intensity below 0.01 were assigned as adipocytes and above the threshold as stroma. The adipocyte ROI was automatically split on individual cell territories, and a standard set of shape features was measured automatically. Analyses were performed using R Statistical Software (v4.1.2; R Core Team 2021). Area and circularity were used to describe the heterogeneity of adipocytes. Cells with an area below 200 μm were filtered out from the initial dataset to exclude nonadipocyte cells. Circularity represents the characteristics of cells, calculated via the following equation:

According to control sample characteristics, normal adipocytes have a circularity range of 0.3–0.8. Objects with circularity below 0.3 were assigned as abnormal adipocytes. For each sample, the total adipocyte and stromal area fraction, the normal and abnormal adipocyte area fraction, was calculated as a sum of corresponding ROI areas to the entire sample area. To compare individual adipocyte areas between control and AAT groups, counts of adipocytes were normalized according to counts in smaller groups by random sampling from larger groups. Vessels were identified by size, circularity, and the content of red blood cells.
Immunofluorescent staining
Tissue sections were deparaffinized in serial dilution of xylene and isopropanol (Sigma-Aldrich). Dehydrated samples were boiled for 40 min at 90°C in antigen-retrieval buffer (25 mM Tris-HCl, 1 mM Ethylenediaminetetraacetic acid [EDTA], 0.05% Sodium Dodecyl Sulfate [SDS] pH 8.5). Sections were blocked with phosphate-buffered saline (PBS) with 0.5% goat serum for 20 min and incubated with a rabbit monoclonal primary antiperilipin antibody (Cat # A4758, ABclonal) overnight, at 4°C. After extensive washing with PBS, sections were incubated with goat-antirabbit Alexa Fluor 647 secondary antibody (Invitrogen, USA) and DAPI (1 μg/mL). Mounted sections were imaged on a Zeiss Cell Discoverer microscope with ×20 Plan Apochromat 0.95 objective and appropriate filters. The resulting images were processed using ImageJ and QuPath ver 0.4.3. For intensity estimation, the adipocyte area was segmented by thresholding and divided into small ROIs of 50 × 50 μm. Metrics for perilipin intensity were measured for each ROI.
Gene expression profile
Tissue samples were submerged in RNA save before processing (Sartorius AG). RNA was isolated using Genezol® reagent (Invitrogen) at 1 μL/well following the manufacturer’s instructions. Then, 200 μg of RNA were reverse-transcribed into cDNA using the Prime Script RT Reagent kit (Takara, China). Quantitative polymerase chain reaction (PCR) was performed using StepOnePlus™ Real-Time PCR System (Thermo Fisher Scientific corporation, USA) and SYBR Premix Ex Taq II (Takara Biomedical Technology, Beijing, Co) at a 20 μL volume reaction. CFX Manager™ Software #1845000, (Bio-Rad Laboratories Ltd. Hercules CA USA) was used to determine threshold cycle values. Specific gene amplification was obtained using the following murine-specific primers (Hy-Labs, Israel): IL-1β; vascular endothelial growth factor (VEGF); adiponectin; collagen-I; CD14; iNOS; arginase-1; AAT. Specific primer sequences are detailed in the Supplementary Data. Analysis was performed using the double-delta method, 35 and fold from sham is presented for relative gene expression.
In vitro cell metabolic function viability assays
Adipocytes and ADSCs were isolated following specific protocols.36,37 Briefly, extracted inguinal fat pads were minced and enzymatically digested with collagenase I or II (Worthington, NJ) and centrifuged until the floating adipocyte phase separated from the SVF. A red-cell-lysis buffer (Thermo Fisher Scientific corporation, USA) was used for ADSC isolation. Cells were filtered through a sterile mesh (420 μm for adipocytes, 100 μm for ADSC, Lapidot, Israel) and incubated in DMEM/F-12 medium (BI, Israel). Cells were cultured in 12-well plates (Bar-Naor, Israel) at 2
LDH cytotoxicity assay
A cytotoxicity assay was performed using Cytotox 96 nonradioactive assay (Promega, WI). According to the manufacturer’s instructions, 50 μL supernatant was transferred to a 96-well assay plate, and 50 μL assay buffer was added. After 30 min at room temperature, OD490nm was determined by a spectrophotometer.
XTT metabolic function assay
Fat samples from 3 days after grafting were minced into <1 mm-sized pieces and placed in a 96-well plate with 100 μL PBS. As per the manufacturer’s instructions (Sigma-Aldrich), 50 μL XTT reagent and activator mix were added to each well. After 4 h at room temperature, OD490/630 nm was determined by a spectrophotometer. The assay was validated for assessing adipocytes in 3D structures.38,39
Statistical analysis
Comparisons were performed using Prism (Ver. 9.5.1, GraphPad Software, LLC). Results are presented as mean ± SEM. t test was used for univariate analysis, and Wilcoxon test was used for sequential comparisons of two groups. For comparison of more than two groups, the Kruskal–Wallis test was performed. Chi-square was used to compare categorical variables. A p value of ≤0.05 was considered statistically significant.
RESULTS
Volume retention rates of AAT-treated fat grafts
To explore the impact of AAT treatment on fat grafts, female mice weighing an average of 20 g underwent syngeneic fat grafting under the scalp (Fig. 1). Conditions included ALB-treated and AAT-treated graft sites during the procedure, and again 3 days later (n = 25/group). According to micro-MRI of 14 animals per group, once positioned under the scalp, graft volumes measured 344.26 ± 27.73 and 343.93 ± 12.03 mm³ (mean ± SEM, ALB-treated and AAT-treated mice, respectively). There were no signs of distress or events of mortality in the subsequent 90-day follow-up.
As shown (Fig. 2A), AAT-treated grafts preserved higher volumes (mean, mm³) throughout the follow-up period, albeit reaching statistical significance on days 10, 60, and 90: day 3, AAT-treated mice 322.38 (93.99%) and control mice 294 (85.6%); day 10, 341.31 (99.51%) and 263 (76.63%) (p = 0.001); day 30, 255.05 (74.36%) and 214.17 (62.26%); day 45, 258.24 (75.29%) and 191.57 (55.69%); day 60, 245.45 (71.56%) and 132.81 (38.61%) (p = 0.02), and day 90, 240.30 (70.06%) and 118.81 (34.54%) (p = 0.02).
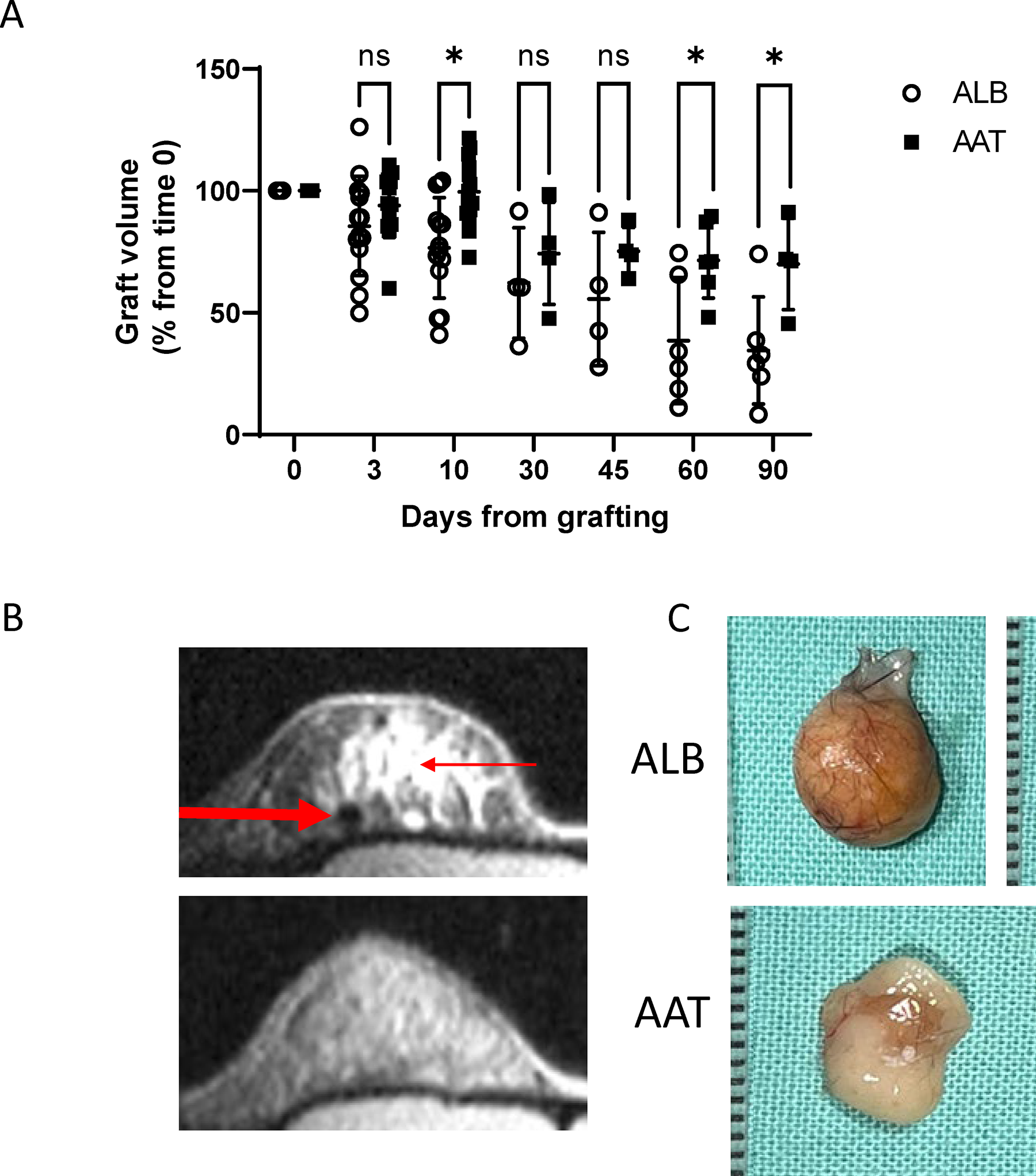
Graft volume and macroscopic appearance 90-day follow-up.
Fat graft characteristics
Figure 2B presents representative day-60 micro-MRI images. Representative ex vivo photographs of fat grafts of similar sizes harvested on day 90 are depicted in Figure 2C.
Explants were obtained for histological examination on days 1, 3, 10, and 60 (n = 6, 6, 22, and 10, respectively). Representative images are presented in Figure 3A. According to a designated pipeline method for objective analysis of digital microscopy, vessels were denser per mm2 in the AAT group compared with the control group (day 3, 4/mm2 and 0.28/mm2; day 10, 1.94/mm2 and 0.33/mm2), and, although not statistically significant, less dense on day 60 (5.5/mm2 and 7.15/mm2) (Fig. 3B).

Fat graft characteristics. C57BL/6 mice were treated with ALB as control, or AAT, at 0 and 3 days of grafting (400 µg/graft directly into graft site).
Relative expression of VEGF was higher in AAT-treated grafts compared with the control on both day 10 (69.57 ± 14.11 and 112.11 ± 8.68, n = 12, p = 0.03) and day 60 (100 ± 5.85 and 57.74 ± 4.67, n = 6, p < 0.01) (Fig. 3C). The AAT group exhibited a higher percentage of adipocytes than the control group (day 10, 49.89% and 40%; day 60, 35.29% and 10.71%, respectively). The portion of morphologically normal adipocytes from that percentage was lower on day 10 in the AAT group compared with the control (38.6% and 51.24%) but higher on day 60 (69.36% and 9.11%) (Supplementary Fig. S1 in Supplementary Data). Adiponectin gene expression on day 10 was higher in the AAT group compared with the control group (0.005 ± 0.002 and 0.0008 ± 0.0002, n = 12, p = 0.02, respectively). Relatively lower collagen-I gene expression was found in the AAT-treated group (5.17 ± 1.3 and 6.46 ± 1.04, n = 12, p = 0.4). Because these findings are limited, being representative only of gene expression and not protein levels, a subsequent analysis was undertaken for the relationship between adiponectin and collagen-I gene expression levels per graft (Fig. 3D). Outcomes were defined as adiponectinhigh/collagen-Ilow or adiponectinlow/collagen-Ihigh (set by the higher value of the two). According to chi-square analysis, compared with the control group, AAT-treated grafts formed a cluster of primarily adiponectinhigh/collagen-Ilow transcripts (72.72% and 16.67%, respectively, n = 11 AAT group, n = 6 control group, p = 0.02).
Inflammatory response-related gene expression profile of treated fat grafts
Relative IL-1β levels were determined to represent upstream inflammatory responses, reaching statistical significance between AAT and control groups on days 10 and 60 (day 1, 14.6 ± 3.4 and 10.87 ± 6.93, n = 6; day 3, 15.04 ± 13.39 and 3.4 ± 1.4, n = 6; day 10, 62.56 ± 15.47 and 120.5 ± 7.7, n = 22, p = 0.01; day 60, 10.2 ± 6.37 and 118.59 ± 11.59, n = 10, p < 0.01) (Fig. 4A). Relative CD14 transcript levels were assessed, representing a marker of immune cell infiltration, but no significant differences were found between the groups (day 3, 2.192 ± 0.27 and 1.97 ± 0.39, n = 6; day 10, 3.53 ± 0.52 and 4.72 ± 1.82, n = 6). The relative degree of expression of iNOS and arginase-1 was evaluated as per their relation to M1 and M2-like gene expression profiles, respectively. As shown in Figure 4B, on day 3, arginase-1 expression exhibited no significant change on transcript levels between the groups (5.95 ± 0.47 in the AAT group, 7.01 ± 0.96 in the control group, n = 6). On day 10, AAT group arginase-1 levels were higher (17.36 ± 1.23 compared with the control 9.77 ± 0.9, n = 6, p = 0.03). In contrast, iNOS transcript levels were higher in the AAT group on day 3 (0.08 ± 0.005 compared with the control 0.04 ± 0.007, n = 6, p = 0.02) and lower on day 10 (0.4 ± 0.005 compared with the control 0.69 ± 0.007, n = 6, p = 0.02) (Fig. 4C). The ratio between iNOS and arginase-1 gene expression levels per graft are presented in Figure 4D.
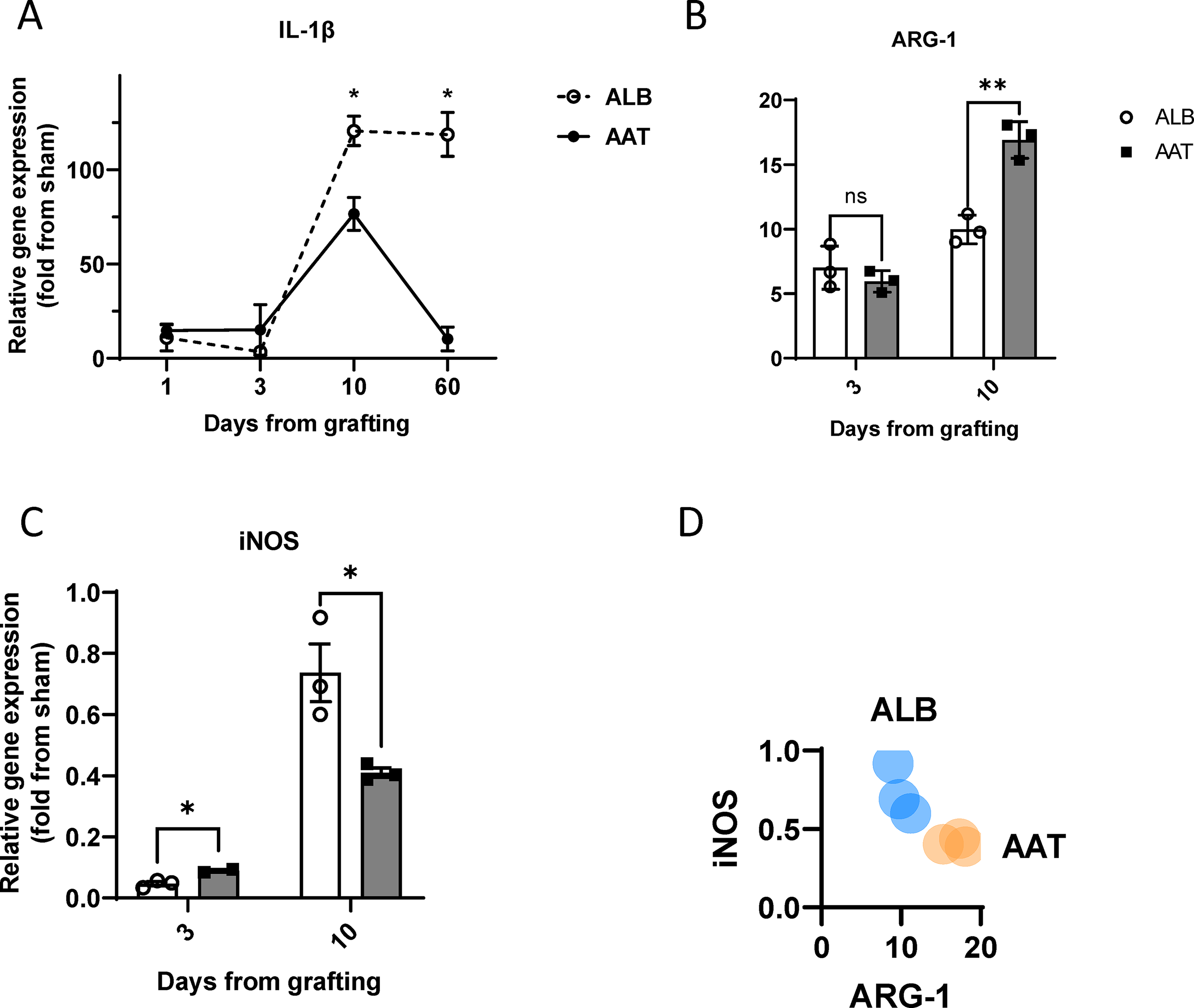
Inflammatory response in fat grafts. C57BL/6 mice were treated with ALB or AAT at 0 and 3 days after grafting (400 µg/graft directly into the graft site). Graft explants underwent RNA isolation and RT-PCR for indicated gene products.
Metabolic function assays of fat grafts, adipocytes, and ADSC under AAT treatment
The possibility of AAT affecting cell viability in the early phase after grafting was addressed. Freshly explanted grafts (n = 6) were evaluated for perilipin content (Fig. 5A) and for metabolic function by XTT assay (Fig. 5B). AAT-treated grafts exhibited a mean OD450/630 nm of 0.67, whereas the control group mean value was 0.43 (p = 0.04), suggesting more viable cells in the AAT-treated group. The area of viable cells, indicated by perilipin intensity >100 (Fig. 5C), demonstrated no apparent difference between the groups at 24 h, yet exhibited more viable cells in the AAT-treated group compared with the control after 72 h (18.5% and 3.1%, respectively) (Fig. 5D).

Ex vivo and in vitro cell viability. Harvested grafts (n = 6), cultured adipocytes, and adipose-derived stem cells (ADSC) were assessed for cell viability via perilipin staining, LDH cytotoxicity assay, and XTT viability assay.
Adipocytes cultured under hypoxic conditions for 24 h exhibited less cytotoxicity in the presence of AAT, as LDH released from injured cells was two-fold lower than LDH released from cells not added AAT. However, according to the XTT assay, under these conditions, there was no significant change in metabolic function with added AAT. ADSCs were cultured under hypoxic conditions for 72 h. According to the LDH release assay, cell injury was increased in control conditions and absent in the presence of AAT. However, according to the XTT assay, metabolic function decreased in both groups under hypoxia (Supplementary Fig. S2 in Supplementary Data).
Interestingly, endogenous murine AAT gene expression was significantly increased in ADSC cultured under hypoxic conditions compared to ADSC cultured under normoxic conditions (2.44 ± 0.7 and 0.27 ± 0.07, respectively, six-plicates, p = 0.04) (Fig. 5E).
DISCUSSION
Multiple approaches are tested to enhance fat graft survival and outcome predictability. To this end, there is no agreement on a working protocol that can provide predictable outcomes, gravely limiting the application of fat grafting.40,41
In this current study, aiming to exhibit and characterize the outcomes of fat graft treated with AAT, the clinically available plasma protein was administered directly to the graft site in a mouse model of fat grafting. Ninety days later, graft volumes remained stable and retained greater volumes in the AAT-treated group, along with less capsule formation, and necrotic centers, either cystic or fibrotic. The results may be attributed to an effective inflammatory response that appears to have accelerated inflammatory resolution, signified, in part, by the early rise and early fall in inflammatory agents such as IL-1β and iNOS, bringing about rapid revascularization and tissue repair in the context of a hypo-oxygenated microenvironment. The relative degree of expression of iNOS and arginase-1 was evaluated in relation to M1 and M2-like gene expression profiles to serve as an initial evaluation and set the ground for further exploration in future studies.
Both in vivo and in vitro aspects of fat grafts were examined. Interestingly, AAT improved cell survival in vivo but had a relatively limited effect in vitro. This observation suggests that the mechanism by which AAT promotes graft survival may involve multiple local factors rather than targeting responses of adipocyte or ADSC under hypoxic stress. The early nonvascularized phase of a fat graft is particularly interesting, as AAT treatment provided benefits to graft viability even before angiogenesis was observable. These findings are in agreement with pancreatic islet grafts, which are limited by blood supply early upon transplantation and have demonstrated improved early viability and function in the presence of AAT treatment. 42
The potential role of ADSCs in fat graft survival has gained recent attention. Similar to AAT, ADSCs modify macrophages to adopt the M2-like phenotype. 43 In general, local mesenchymal stem cells (MSCs) bring about similar outcomes to those achieved by AAT augmentation with respect to diverting inflammation toward resolution. 44 MSCs release AAT in response to inflammatory triggers 22 and synergize with AAT to mitigate acute inflammation, 44 and MSCs respond to AAT by proliferation. 45 Here, we describe the hypoxia-induced endogenous expression of AAT by ADSCs, suggesting that a protective role for local AAT is inherently present in fat tissue repair processes. The relationship between AAT and ADSCs is an intriguing field to explore in future studies, particularly that which relates to underlying hypoxic conditions.
Although overexpression of AAT represents potential interventional treatment in humans, one would assume that AAT knockout would indicate its role as an endogenous agent. However, mice have several copies of AAT across their genome, all expressed during acute phase and hypoxia, and all inhibit elastase, rendering the feat of a complete knockout problematic. 46 Humans, in contrast, have only one gene for AAT, and in its deficiency, it is both the lack of AAT in serum that causes pathology and the accumulation of erroneously coded protein inside liver cells. Mice that express such mutated AAT do indeed represent lower serum levels of endogenous circulating AAT, but they also carry the liver condition and associated morbidity.47–49 As the normal levels of AAT in the circulation rise 4–6-fold during acute phase and hypoxia, the addition of exogenous AAT represents the difference between steady-state and inducible physiological concentrations of AAT. Since the gene for AAT is responsive to hypoxia, it is unsurprising that AAT is tissue-protective in conditions of ischemia-reperfusion injury and that tissue irrigation is improved by treatment with AAT in skin flap models and equivalent conditions. 21 Although promoting angiogenesis and cell survival may raise concerns in postoncologic reconstructive procedures, it is noted that AAT was found to delay tumor development and metastatic spread in a model of melanoma cell inoculation; 50 in that study, AAT significantly enhanced antitumor NK cell activities. In addition, the present protocol is reserved for the first 3 days of grafting. Interestingly, 60 days after grafting, AAT-treated grafts had fewer blood vessels per mm2 than in control grafts, suggesting that effective blood supply had been achieved, diminishing the drive for further angiogenesis.
INNOVATION
Together with its availability and safety, the present study suggests that AAT is a potential agent for improving fat graft survival and enhancing procedure predictability. Additional research may reveal the underlying mechanisms of those results. Advancements of this kind may improve fat graft use for clinical applications and be utilized to treat wounds and scars and reconstruct damaged tissue.
KEY FINDINGS
AAT improved fat graft retention rates and tissue quality in a murine model
AAT treatment shows an improved immune response that favors tissue repair
Treating fat grafts with local AAT yielded faster and time-limited angiogenesis
Footnotes
ACKNOWLEDGMENTS AND FUNDING SOURCES
No funding was received to conduct this study.
AUTHOR DISCLOSURE AND GHOSTWRITING
The authors declare no competing interests forming this research.
ABOUT THE AUTHORS
Supplemental Material
Abbreviations and Acronyms
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.