
Abstract
Objective:
Oral mucosal wound healing is not completely understood, and effective therapies are lacking. This study explores the potential of an adipose-derived stem cell (ADSC) exosome sheet in enhancing intraoral wound healing in rats.
Approach:
An ADSC exosome sheet derived from Tisseel and rat adipose tissue (ADSC-exo) was applied to 16 rats with 6 mm full-thickness mucosal hard palate wounds. Eight wounds received ADSC-exo with a superficial occlusive dressing (ADSC-exo group), and eight received only an occlusive dressing (control group). Wound closure was monitored on days 0, 2, 4, 7, and 10, with dressings changed every 2 days. On day 10, rats were sacrificed, and wounds (n = 8 per group) were collected for immunohistochemical analysis. In vitro, four ADSC-exosome concentrations (0, 4.5 × 1011, 9 × 1011, and 18 × 1011 exosomes/mL; n = 4 per group) were applied to rat oral mucosal fibroblasts to assess migration speed.
Results:
ADSC-exo accelerated wound closure (18% ± 5% vs. 35% ± 9% of initial wound area; p = 0.002) and fibroblast migration (for 18 × 1011 exosomes/mL at 24 h: 29.7% ± 3% vs. 62.2% ± 4% of initial gap area; p < 0.0001) compared with the control. ADSC-exo promoted reepithelialization (87% ± 14% vs. 21% ± 6%; p < 0.0001), proliferation (34 ± 12 vs. 18 ± 7 Ki67+/high-power field [HPF]; p = 0.004), and neovascularization (28 ± 9 vs. 11 ± 5 CD31+/HPF; p = 0.0002) while reducing inflammation (4 ± 1 vs. 13 ± 9 CD68+/HPF; p < 0.0001) and increasing M2 macrophages (9.2 ± 2 vs. 4.2 ± 3 CD163+/HPF; p = 0.0008). ADSC-exo increased Transforming Growth Factor beta 1 (TGF-β1) (1.3 ± 0.3 vs. 0.9 ± 0.2; p = 0.006), Smad3 (0.9 ± 0.02 vs. 0.7 ± 0.1; p = 0.006), and collagen I (1.5 ± 0.9 vs. 0.5 ± 0.3; p = 0.005) while downregulating caspase-3 (0.7 ± 0.3 vs. 1.1 ± 0.2; p = 0.003) and Bax (0.9 ± 0.2 vs. 1.4 ± 0.1; p < 0.0001).
Innovation:
This is the first study to demonstrate the pro-wound healing effects of an ADSC exosome sheet on intraoral wounds. This paves the way for future research and clinical applications of ADSC exosomes in mucosal wound healing.
Conclusions:
Application of an ADSC-exo to rat mucosal wounds significantly improved wound healing. Mechanistically, these effects may be linked to upregulated activity of the TGF-β/Smad pathway.
Huang-Kai Kao, MD
Keywords
INTRODUCTION
Oral mucosal wounds resulting from surgical procedures, trauma, and certain medical conditions present unique challenges in wound healing.1,2 The oral cavity’s constant physical trauma and bacteria-rich environment can lead to chronic and poorly healing wounds that can significantly interfere with essential daily activities such as eating and speaking.1–3 Many patients experience recurrent nonspecific stomatitis on the oral mucosa, which often presents as aphthous ulcers. Recurrent aphthous stomatitis is the most common form of oral ulceration, with its prevalence in the general population ranging from 5% to 60%.4,5 Characterized by recurrent oral mucosal ulcers in otherwise healthy individuals, recurrent aphthous stomatitis can continue into adulthood and persist throughout life. Oral mucositis is also a significant concern for patients undergoing radiation therapy. A cohort study published by Iovoli et al. in 2023 found that 62% of patients (360 out of 576) undergoing radiation therapy for head and neck cancer developed severe oral mucositis. 6
These issues are further compounded in specific patient populations, such as those with diabetes, where wound healing is significantly impaired.1,3,6 While cutaneous wound healing is well-understood, there is a notable lack of knowledge regarding intraoral mucosal healing, which has hindered the development and translation of targeted treatments.1,6
Several regenerative approaches have recently demonstrated promise in addressing the challenges of oral mucosal wound healing. Current therapeutic approaches being investigated include synthetic polymers,7,8 biological grafts,9–11 gel-like ointments,12–14 hybrid scaffolds, 15 and ultrasound-based therapy.16,17 However, there is still an absence of proper, comprehensive analyses that effectively evaluate the success of treatment methods using both qualitative and quantitative evidence. Ironically, very few of the studies investigated changes in immune response following treatment, although chronic wound healing is often attributed to prolonged or impaired immunoregulation. 18
Adipose-derived stem cells (ADSCs) are considered the most advantageous therapy in modern regenerative medicine due to the abundance of adipose tissue sources, their remarkable proliferative capacity, and ease of isolation. ADSC exosomes, which are the primary mediators of the biological effects, are small, single-membrane nanovesicles released through paracrine secretion, containing a rich cargo of proteins, lipids, and nucleic acids. ADSC exosomes do not pose the same risk of tumorigenesis as ADSC tissue19,20 and have demonstrated therapeutic potential in wound healing, organ diseases, neurodegenerative conditions, and cancer. 21 In the context of regeneration and wound healing, ADSC exosomes regulate persistent inflammation, angiogenesis, and extracellular matrix (ECM) reconstruction. They exhibit functions akin to their parent stem cells but offer enhanced safety and efficacy in clinical settings, thereby reducing the risks associated with cell transplantation. 22 However, studies on the effects of ADSC exosomes on oral mucosal wound healing have not been reported.
CLINICAL PROBLEM ADDRESSED
The unique environment of the oral cavity, characterized by constant physical trauma and a bacteria-rich milieu, presents significant challenges for wound healing. The limited availability of effective therapies for oral mucosal wound healing has contributed to a high prevalence of persistent inflammatory oral conditions, significantly diminishing quality of life. Currently, there is a lack of treatments that effectively promote oral mucosal wound healing. This underscores the critical need for developing novel, effective therapies to address this clinical challenge.
MATERIALS AND METHODS
Animals
A total of sixteen 10-week-old male Sprague–Dawley (SD) rats (average weight: 230 ± 22.5 g; National Laboratory Animal Center, Taiwan) were purchased and acclimatized for 2 weeks in the Chang Gung Memorial Hospital vivarium. Animal experiments were carried out after 2 weeks according to, and as approved by, the Institutional Animal Care and Use Committee at Chang Gung Memorial Hospital (approval number IACUC2020061904).
ADSC exosome sheet preparation
This study follows all the Minimal Information for Studies of Extracellular Vesicles (MISEV) 2023 reporting standards. 23
Adipose tissue (1.0 g) was harvested and weighed from the groin region of a rat and then minced into small pieces and washed with phosphate-buffered saline (PBS). The tissue was digested at 37°C for 30 min with continuous shaking in PBS containing 0.075% collagenase (Wako Pure Chemical Industries, Osaka, Japan). Following digestion, mature adipocytes and connective tissue were separated from the pellet by centrifugation at 800g for 10 min. The pellet, enriched with stromal vascular fraction cells, was suspended and immediately plated in T225 culture flasks. The culture medium used was 50 mL of M-199 supplemented with 10% fetal bovine serum (FBS), 100 IU/mL penicillin, 100 µg/mL streptomycin, 5 µg/mL heparin, and 2 ng/mL acidic fibroblast growth factor. For culturing of ADSCs, exosome-free Dulbecco’s modified Eagle’s medium (DMEM; Gibco Invitrogen/Life Technologies, Carlsbad, CA, USA) and 10% FBS (Gibco Invitrogen/Life Technologies) were added.
After 3 days of incubation at 37°C in a 5% CO2 incubator, culture supernatants from conditioned media were collected and mixed with 5% exosome-depleted FBS (Thermo Fisher Scientific, Waltham, MA, USA). After centrifugation at 300g for 10 min to remove cells and large debris, extracellular vesicles (EVs) were precipitated from the resulting supernatant using reagents from the Exo-spin exosome purification kit (Cell Guidance Systems, St. Louis, MO, USA). After incubation at 4°C for 12 h, the solution was centrifuged at 2,000g using a microcentrifuge collection tube, and the final purified EV fraction was obtained and lyophilized.
Three Tisseel-ADSC exosome sheets (ADSC-exo) were prepared using the ARTISS Kit (Baxter International, Deerfield, IL, USA) following the standard protocol, as shown in Fig. 1. First, 200 µg, 100 µg, and 50 µg aliquots of the lyophilized ADSC EV fraction were dissolved in 200 µL of PBS to create three different concentrations of ADSC EV solutions: 1, 0.5, and 0.25 µg/µL. Next, 400 µL of each ADSC EV solution was mixed with 600 µL of a fibrinolysis inhibitor solution and a sealer protein concentrate. Separately, 3,000 µL of calcium chloride solution was mixed with thrombin. Finally, three distinct ADSC-exo sheets were created by combining each one of the ADSC EV solutions (0.25, 0.5, or 1 µg/µL) with 1,000 µL of the calcium chloride and thrombin mixture. This resulted in the formation of three different Tisseel sheets (ADSC-exo), each embedded with different concentrations of EVs.
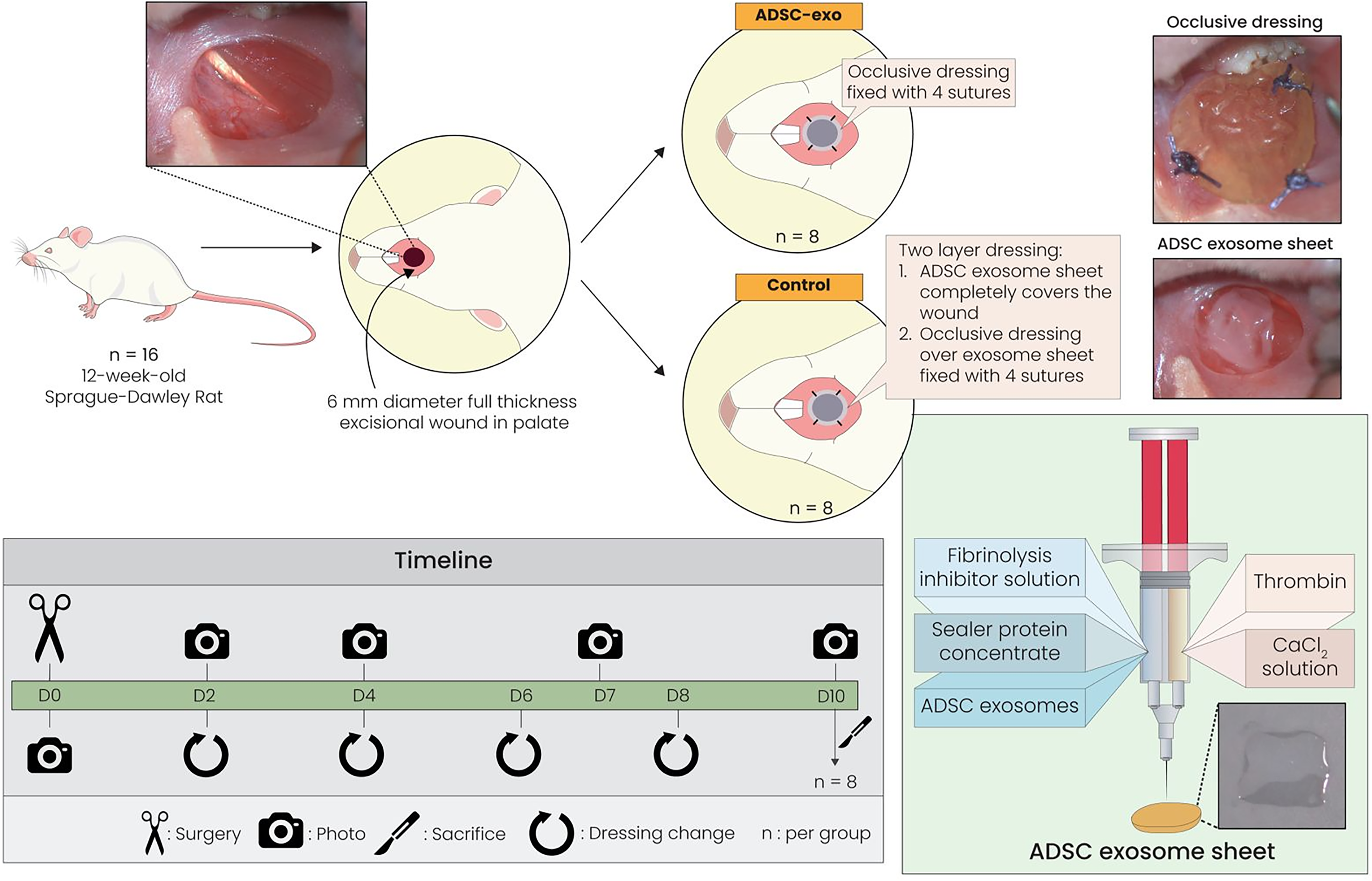
Study design. Full-thickness wounds (6 mm diameter) were induced on the oral hard palate of 16 12-week-old Sprague–Dawley (SD) rats. The wounds of eight rats were covered with an occlusive dressing, while those of the remaining eight were completely covered by a two-layer dressing of adipose-derived stem cell exosome sheet (ADSC-exo), followed by an occlusive dressing. Dressings were changed on days 2, 4, 6, and 8 postsurgery. Wounds were photographed on days 0, 2, 4, 7, and 10 postsurgery. On day 10 postsurgery, the rats were sacrificed for tissue harvesting and histological analysis. The preparation of ADSC-exo used in the rat experiments involved suspending 200 µg of adipose-derived stem cell-extracellular vesicles (ADSC EVs) in 200 µL of phosphate-buffered saline (PBS) to achieve a 100% solution, followed by mixing with a fibrinolysis inhibitor solution and sealer protein concentrate to create fibrinogen. Tisseel hemostasis solution was made by adding calcium chloride solution to thrombin. These two solutions were then mixed to complete the preparation of the ADSC-exo used in all animal experiments.
Fibroblast migration assay
A 6-mm-diameter full-thickness sample of intraoral palatal mucosa was harvested from a healthy SD rat under anesthesia using 2–4% isoflurane vapor in an induction chamber. The tissue was then rinsed in sterile PBS and minced into small fragments (approximately 1 mm³) using sterile scissors. Enzymatic digestion was performed by incubating the tissue in a collagenase solution (0.1% collagenase type I in DMEM) at 37°C for 2–4 h with gentle agitation. The digested tissue was filtered through a 70 µm cell strainer to remove undigested fragments, followed by centrifugation at 300g for 5 min. The supernatant was discarded, and the resulting cell pellet was resuspended in complete culture medium (DMEM supplemented with 10% FBS). The cell suspension was then plated in tissue culture flasks and incubated at 37°C in a humidified atmosphere with 5% CO2, with medium changes every 2 days to facilitate cell adherence and proliferation. Within 4 days, the culture predominantly consisted of fibroblasts due to their rapid adherence and growth. To further purify the fibroblasts, differential trypsinization was applied. Finally, fibroblast identity was confirmed through morphological assessment under a light microscope, where the cells exhibited a characteristic spindle-shaped appearance. This method resulted in a population of rat fibroblasts suitable for in vitro cell migration studies.
Fibroblast migratory velocity from healthy SD rats was assessed in vitro using a culture insert migration assay (ibidi, Gräfelfing, Germany) as per the manufacturer’s instructions. A physical gap was created in a monolayer of rat fibroblasts suspended in PBS, and cell migration into the gap was imaged at 0, 12, and 24 h. The gap closure rate, a standard metric, was analyzed using ImageJ software (V1.52a; Media Cybernetics) by measuring the empty space at each time point relative to the initial empty area. The procedure was repeated for four rats (n = 4) per group, with results averaged for each of the following groups: a Tisseel control supernatant (Tisseel incubated with PBS for 2 days) and three treatment groups. The three treatment groups consisted of supernatants from ADSC-exo (made with 0.25, 0.5, and 1 µg ADSC EVs/µL) collected after 2 days of incubation in PBS. The ADSC-exo with the highest closure rate in the in vitro analysis was selected for exosome characterization and the rat experiments.
Exosome characterization
The assessment of EV release ratio from the ADSC-exo was done with a NanoSight System (NS300; Malvern Instruments, Malvern, United Kingdom). First, a 20% ADSC-exo solution was prepared by mixing a 6-mm-diameter punch-out of the ADSC-exo (made with 1 µg ADSC EVs/µL) with 3 mL of PBS. This mixture was incubated in a 15 mL tube for 2 days to create the ADSC-exo supernatant. After 2 days of incubation at 37°C, the supernatant was collected and transferred to a separate 6 mL tube for analysis. Nanoparticle tracking analysis (V3.20; Malvern Instruments) was then performed to measure the concentration of particles and check if the size distribution of the EVs in the ADSC-exo supernatant matched the expected size distribution for exosomes. 23
The morphological characterization of EVs within the ADSC-exo was conducted using transmission electron microscope (TEM; Hitachi-7000FA; Tokyo, Japan) to confirm the identity of embedded EVs as exosomes. 23 To quantify the protein expression of exosome-associated tetraspanins, including CD9, CD63, and CD81 (Abcam, Cambridge, United Kingdom), 23 Western blot analysis was performed on the ADSC-exo supernatant. The potential presence of cellular contaminants was evaluated by probing for calnexin (Abcam) via Western blot. 23 A supernatant taken from a Tisseel control sheet served as a comparative reference for all Western blots for exosome characterization.
Surgical procedure and monitoring
Sixteen 12-week-old SD rats were utilized in this study, as illustrated in Fig. 1. The rats were anesthetized using 2–4% isoflurane vapor in an induction chamber. Subsequently, all rats underwent a 6-mm-diameter full-thickness oral wound in the mucosal hard palate. The location of the wound on the palate was standardized by centering the excision medially along the palate, just below the first rugae. The use of oral spacers, prepared from pipette tips, helped to keep the mouth open and provided clear visibility of the palate during the procedure. A sterile 6 mm KRUUSE biopsy punch tool was used to create a circular excisional wound. The punch was rotated in a circular motion until friction was felt, indicating complete excision of the epithelial tissue. The excisional wound extended through the epithelium but left the periosteum intact.
The 16 rats were then divided into two equal groups: control and treatment. The control group (n = 8) received a standard occlusive dressing (Tegaderm HP Transparent Dressing; 3M Health Care, St. Paul, MN, USA). The treatment group (n = 8) received a two-layer dressing: a 6-mm-diameter circular punch-out of the ADSC-exo (made using 1 µg ADSC EV/µL) was applied directly to the wound bed, followed by a standard occlusive dressing superficially to secure the ADSC-exo. The occlusive dressing for both groups was secured at four corners with 4–0 Prolene (Ethicon, Inc., Somerville, NJ, USA) to prevent rats from removing it. Buprenorphine (subcutaneous; 0.05 mg/kg) was administered to all rats for analgesia immediately after surgery and every 12 h for 48 h. ADSC-exo and occlusive dressings were replaced every 2 days (days 0, 2, 4, 6, 8) postsurgery for both wound inspection and to maintain a consistent secretion of exosomes from ADSC-exo into the surrounding tissue. Photographs of wounds were taken on days 2, 4, 7, and 10 postsurgery. On day 10, all rats were euthanized using carbon dioxide asphyxiation, and the entire wound was collected.
For tissue harvesting, curved scissors and dissector scissors were used to separate and remove the palate along with underlying bone tissue. Forceps were used to carefully handle and extract the wound bed and 2 mm of wound border. The collected specimens were halved at the midline, with one half of each wound fixed in formalin for histological analysis, and the other half cryopreserved with liquid nitrogen for Western blotting.
Wound area assessment
The wounds, including a ruler, were photographed for all 16 rats (Canon PowerShot G7X) on days 0, 2, 4, 7, and 10. The images were imported into ImageJ (version 1.52a; Media Cybernetics, Rockville, MD, USA), and the area was measured by outlining the wound border under double-blinded conditions. Healing was assessed with the following equation:

Histology
Formalin-fixed tissues were embedded in paraffin, sectioned into 5 µm slices, and analyzed on day 10 postsurgery. All sections used in this study were obtained starting from the center of the halved wound ensuring to include both the wound bed and the wound border. One section from each rat wound (n = 16) was first stained with standard hematoxylin and eosin (HE). 24 An image of the HE stained was taken per sample (n = 8 per group) at 10× magnification using an Olympus BX53 microscope (UCMAD3, T7; Tokyo, Japan). Wound reepithelialization was assessed for each rat using ImageJ software (V1.52a; Media Cybernetics) by measuring the epithelial distance (the distance between original wound edge and leading edge of epithelialization) divided by the wound bed diameter on D0. Two independent, blinded researchers (D.M. and H.K.K.) performed the analysis.
For immunofluorescent staining, one deparaffinized and rehydrated wound cross section was probed for each rat (n = 16) for each of the following antibodies: anti-Ki-67 (1:200; cellular proliferation marker; Lab Vision, Freemont, CA, USA), anti-K5/6 (1:200; cytokeratin 5/6 marker; Thermo Fisher Scientific, MA, USA), anti-CD31 (1:200; neovascularization marker; Pharmingen, San Jose, CA, USA), anti-CD68 (1:200; pan-macrophage marker; Abcam, Cambridge, United Kingdom), and anti-CD163 (1:100; M2-macrophage marker; Bio-Rad Laboratories Inc, CA, USA). All slides were coated with ProLong Diamond Antifade Mountant (DAPI, P36971; Invitrogen, Carlsbad, CA, USA) to probe for all cellular nuclei. Three representative high-power field (HPF) photographs of the wound bed cross section were taken and averaged per animal (n = 8 per group) using an Olympus microscope at 20× magnification (UCMAD3; T7).
The color deconvolution plugin on ImageJ (V1.52a; Media Cybernetics) was used to quantify the number of Ki-67+ and CD31+ cells per HPF. To measure inflammation, the CD68+ and CD163+ cells were manually counted to estimate the average number of macrophages per HPF. For any background staining, we carefully and consistently adjusted the brightness threshold to reduce random noise on an image-by-image basis.
Western blotting
For ADSC-exo characterization, the supernatant from ADSC-exo prepared with 1 µg ADSC/µL over 2 days was analyzed using primary antibodies against CD9, CD63, CD81, and calnexin (Abcam, Cambridge, United Kingdom). For Western blot characterization of rat wound tissue, one half of the wound tissue was analyzed per rat (n = 16) using the following primary antibodies: anti-Bax (pro-apoptosis marker, Abcam, Cambridge, United Kingdom), anti-caspase-3 (pro-apoptosis marker, Thermo Fisher Scientific, MA, USA), anti-collagen I (fibrosis marker; Abcam, Cambridge, United Kingdom), anti-collagen III (fibrosis marker; Abcam, Cambridge, United Kingdom), anti-Smad3 (signaling protein; Abcam, Cambridge, United Kingdom), and anti- Transforming Growth Factor beta 1 (TGF-β1); fibrosis marker; Abcam, Cambridge, United Kingdom).
All samples were lysed in ice-cold lysis buffer containing a cocktail of protease and phosphatase inhibitors. Equal amounts of protein extracts were separated using sodium dodecyl sulfate–polyacrylamide gel electrophoresis and subsequently subjected to Western blotting. Chemiluminescent signals (LumiLight, Roche Diagnostics, Basel, Switzerland) were detected with a Lumi-Imager or X-ray films and quantified using the LumiAnalyst program (Roche Diagnostics, Rotkreuz, Switzerland). Densitometry was analyzed with ImageJ software (V1.52a; Media Cybernetics); β-actin and β-tubulin served as the housekeeping references.
Statistical analysis
Data collection was performed using Microsoft Excel 2020 (Microsoft, Redmond, WA, USA). All results presented in this article are denoted as mean ± standard deviation. For the cell migration assay, a one-way analysis of variance was conducted to identify statistically significant differences in mean gap area covered between the ADSC-exo-treated fibroblasts and the Tisseel-treated fibroblasts at each time point (0, 12, and 24 h). For all other analyses—wound area assessment, staining, and western blotting—comparisons were done between the average measurements of the ADSC-exo-treated wounds and the control wounds using a simple Student’s t-test. For multiple comparisons, p-values were adjusted using the Bonferroni correction method. Statistical analyses and graphical representation of the results were carried out using GraphPad Prism (V8.00, MacOS; GraphPad Software, Boston, MA, USA). Statistical significance was set at a p < 0.05 for all analyses.
RESULTS
ADSC-exo characterization and effects on in vitro fibroblast migration
Nanosight concentration analysis revealed that after a 2-day incubation with PBS, the exosome concentrations in the supernatant were as follows: 4.5 × 1011 exosomes/mL for the ADSC-exo prepared with 0.25 µg ADSC EVs/µL, 9 × 1011 exosomes/mL for the 0.5 µg ADSC EVs/µL preparation, and 18 × 1011 exosomes/mL for the 1 µg ADSC EVs/µL preparation.
Fibroblasts from healthy SD rats had substantially enhanced migration when treated with any concentration of ADSC-exo compared with the control (Fig. 2). At 12 h, the control group had covered 18.0% ± 4.2% of the original gap area, while groups with 4.5 × 1011 exosomes/mL, 9 × 1011 exosomes/mL, and 18 × 1011 exosomes/mL covered 40.1% ± 5%, 34.3% ± 0.9%, and 25.0% ± 4% of the gap, respectively (n = 4 per group; p < 0.0001). At 24 h, the control group covered 29.7% ± 3% of the original area, while the 4.5 × 1011 exosomes/mL treatment group covered 54.2% ± 4%, the 9 × 1011 exosomes/mL group covered 53.5% ± 2%, and the 18 × 1011 exosomes/mL group covered 62.2% ± 4% of the original area (n = 4; p < 0.0001) (Fig. 2). Since the 18 × 1011 exosomes/mL ADSC-exo group showed the highest gap closure rate, it was selected as the ADSC-exo treatment for all exosome characterization and rat experiments.
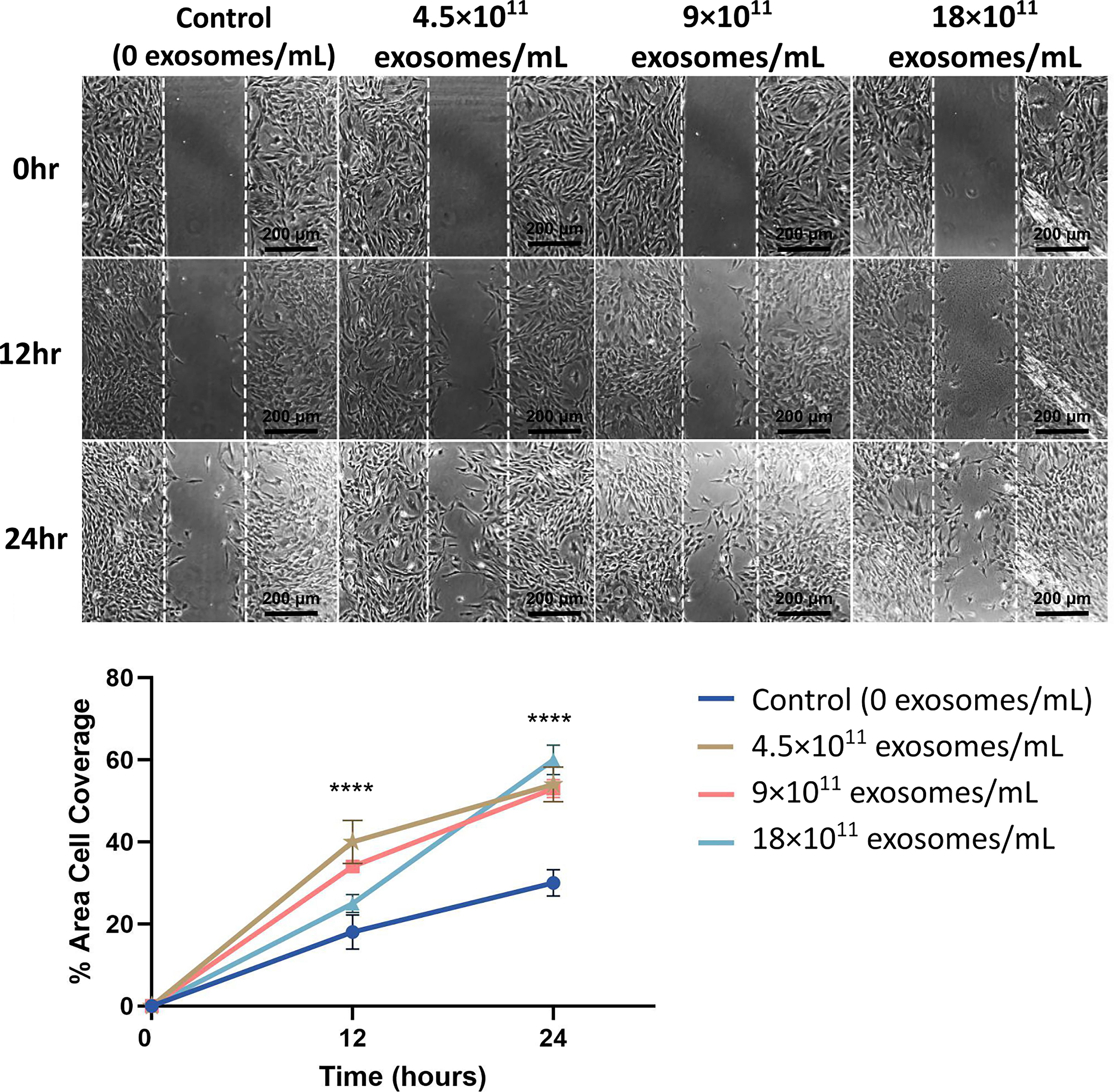
Representative images of the fibroblast cell migration assay along with quantitative analysis of % area cell coverage. Scale is 200 µm. Fibroblasts from healthy SD rats displayed markedly higher % area coverage of the initially empty space when treated with various concentrations of ADSC-exo supernatant (n = 4; 50 µg/mL (4.5 × 1011 exosomes/mL), n = 4; 100 µg/mL (9 × 1011 exosomes/mL), and n = 4; 200 µg/mL (18 × 1011 exosomes/mL) compared with the Tisseel control (n = 4 for each group) at both 12-h and 24-h time points.
Analysis of TEM micrographs revealed that embedded ADSC EVs exhibited characteristic features of exosomes, including hollow vesicle spherical structures with a denser core and less dense periphery (Fig. 3A). Supernatants taken from ADSC-exo in PBS for 2 days were extracted and analyzed for size distribution of EVs, which ranged from 10% to 90% approximately between 80 and 212 nm. The distribution displayed a right-skewed pattern, with a mean diameter of approximately 134 nm and a mode of 104 nm (Fig. 3B), consistent with the exosome size range of 40–160 nm reported in the literature. 23 In terms of protein expression, the levels of exosome-associated CD9, CD63, and CD81 were notably elevated in the ADSC-exo supernatant compared with a control Tisseel supernatant (Fig. 3C). The levels of calnexin were absent in both groups (Fig. 3C).
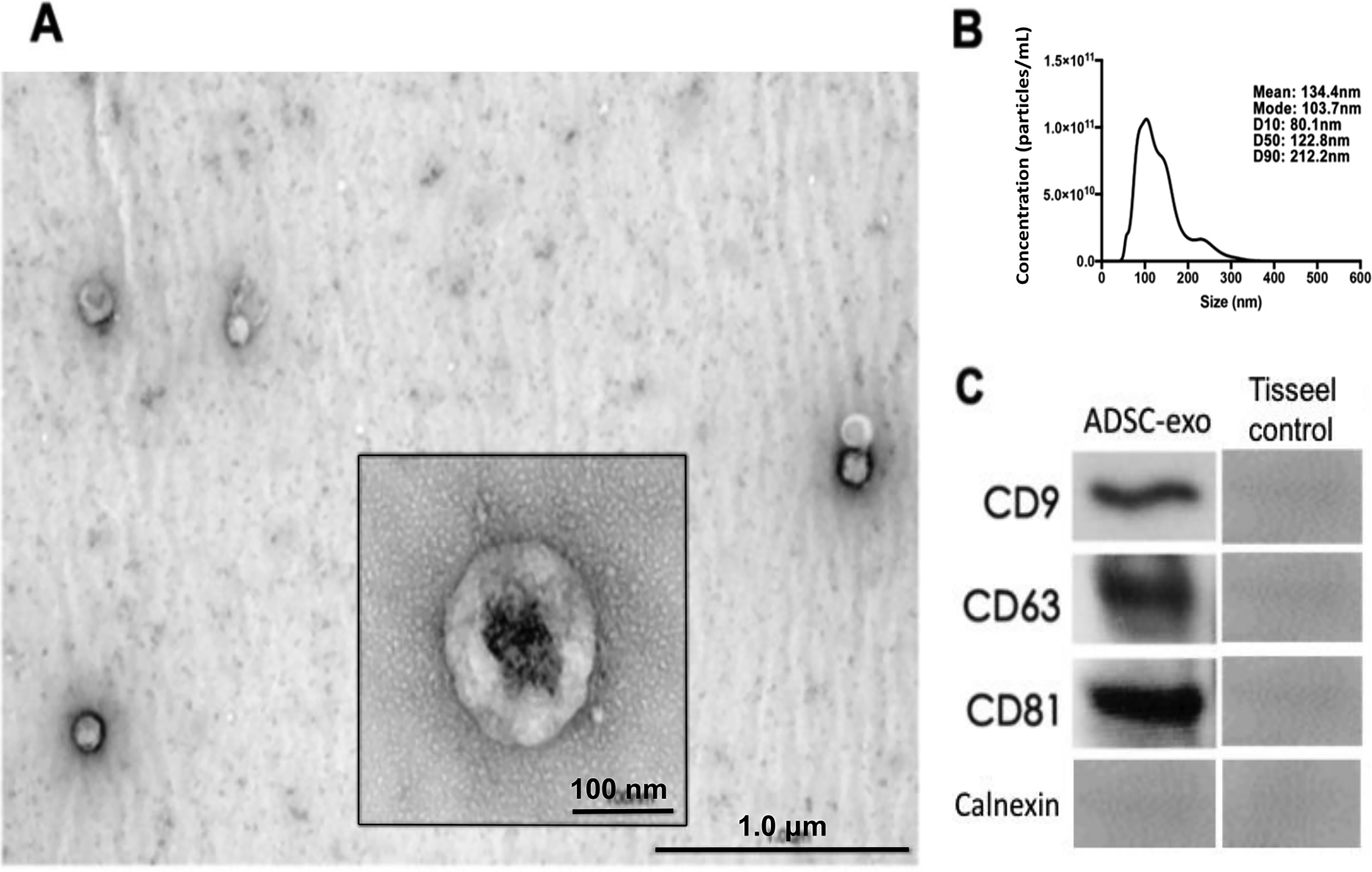
Characterization of ADSC exosomes.
ADSC-exo promotes oral mucosal wound closure
On day 10 following surgery, the wounds in the ADSC-exo group exhibited a significantly smaller wound (18% ± 5% of the initial wound area) compared with the control group (35% ± 9% of the initial wound area; p = 0.002), indicating that treatment of ADSC-exo accelerated the rate of wound healing (Fig. 4).

Representative images of wounds at each time point and quantitative analysis of wound healing kinetics. Scale is 3.0 mm. By day 10 postsurgery, wounds in the ADSC group displayed a significantly reduced area (n = 8; 18.1% ± 5% of initial wound area) compared with the control group (n = 8; 34.7% ± 9% of initial wound area; p = 0.002), suggesting accelerated wound healing with ADSC exosome sheet treatment.
ADSC-exo enhances reepithelialization, cell proliferation, angiogenesis, while regulating inflammation
Epithelialization exhibited a notable increase in the ADSC-exo group (87% ± 14% reepithelialization of the initial wound cross section diameter) compared with the control group (21% ± 6% reepithelialization of the initial wound cross section diameter; p < 0.0001) on day 10, suggesting that ADSC-exo promotes keratinocyte remigration across the wound surface (Fig. 5). Furthermore, cellular proliferation was significantly elevated in the ADSC group (33.5 ± 11.9 Ki67+/HPF) compared with the control (18.3 ± 7.4 Ki67+/HPF; p = 0.004). Regarding neovascularization, the overall blood vessel density was notably higher in the ADSC-exo group (28.2 ± 8.8 CD31+/HPF) than in the control group (11.2 ± 5.1 CD31+/HPF; p = 0.0002).
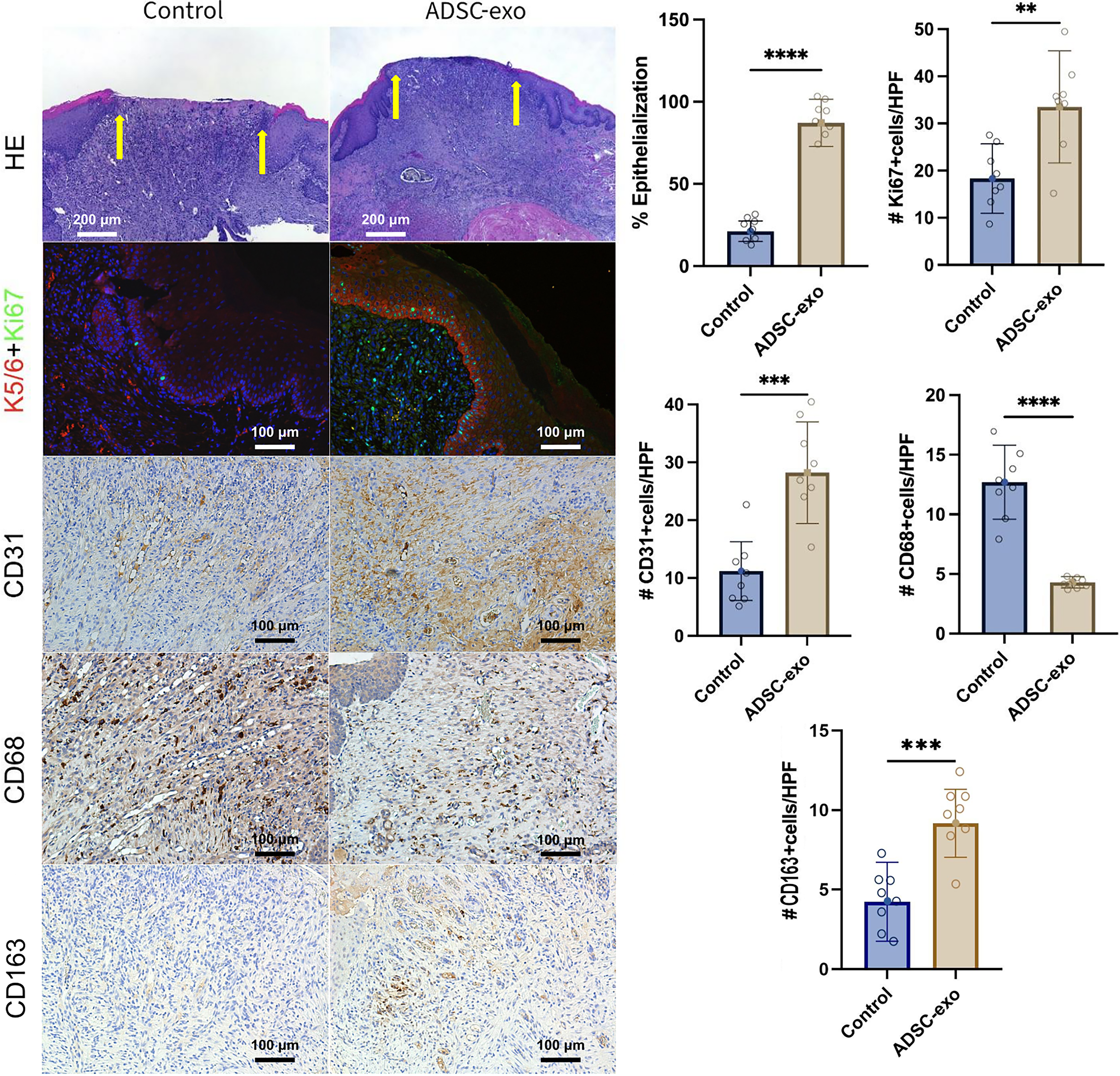
Histological analysis. Presentation of representative images and quantitative findings. Enhanced reepithelialization of the wound bed (margins marked by yellow arrows), measured from 10× hematoxylin and eosin (HE) images (scale is 200 µm), was evident in the ADSC-exo group (n = 8; 87% ± 14% reepithelialization of initial wound diameter) compared with controls (n = 8; 21.2% ± 6%; p < 0.0001), indicating ADSC-exo promotes keratinocyte remigration. Cellular proliferation, assessed via anti-Ki67 probing of 20× images (scale is 100 µm; green fluorescence for Ki67, red for K5/6), was significantly higher in the ADSC group (n = 8; 33.5 ± 11.9 Ki67+/high-power field [HPF]) compared with controls (n = 8; 18.3 ± 7.4 Ki67+/HPF; p = 0.004). Neovascularization, evaluated by CD31 probing of 20× images (scale is 100 µm; light brown color), was notably higher in the ADSC-exo group (n = 8; 28.2 ± 8.8 CD31+/HPF) versus controls (n = 8; 11.2 ± 5.1 CD31+/HPF; p = 0.0002). Conversely, pan macrophage infiltration, indicated by anti-CD68 probing of 20× images (scale is 100 µm; dark brown color), was significantly decreased in the ADSC-exo group (n = 8; 4.3 ± 1.3 CD68+/HPF) compared with controls (n = 8; 12.7 ± 8.8 CD68+/HPF; p < 0.0001). However, M2 macrophage presence, indicated by anti-CD163 probing on the 20× images (scale is 100 µm; dark brown color), was significantly higher in the ADSC-exo group (n = 8; 9.2 ± 2.4 CD163+/HPF) compared with controls (n = 8; 4.2 ± 2.5 CD163+/HPF; p = 0.0008). **p < 0.01, ***p < 0.001, ****p < 0.0001.
Pan-macrophage infiltration into the wound environment was significantly reduced in the ADSC-exo group (4.3 ± 1.3 CD68+/HPF) compared with the control group (12.7 ± 8.8 CD68+/HPF; p < 0.0001). However, M2 macrophage infiltration was significantly higher in the ADSC-exo group (9.2 ± 2.4 CD163+/HPF) compared with controls (4.2 ± 2.5 CD163+/HPF; p = 0.0008) (Fig. 5).
ADSC-exo increases TGF-β1, Smad3, and collagen I deposition, while reducing apoptosis
Western blot analysis revealed significant differences in the ratio of TGF-β1 to β-tubulin, indicating a markedly higher ratio in the ADSC-exo group (1.3 ± 0.3) compared with the control group (0.9 ± 0.2; p = 0.006) on day 10. In addition, Smad3/β-actin densitometry ratio was significantly higher in the ADSC-exo group versus control (0.9 ± 0.02 vs. 0.7 ± 0.1; p = 0.006). Similarly, the ratio of collagen I to β-tubulin was significantly elevated in the ADSC-exo group (1.5 ± 0.9) compared with the control group (0.5 ± 0.3; p = 0.005). However, no significant differences were observed in terms of collagen III production. The ratio of apoptotic caspase-3 to β-actin was lower in the ADSC-exo group (0.7 ± 0.3) compared with the control group (1.1 ± 0.2; p = 0.003). Similarly, the ratio of Bax to β-actin was lower in the ADSC-exo group (0.9 ± 0.2) compared with the control group (1.4 ± 0.1; p < 0.0001) (Fig. 6).
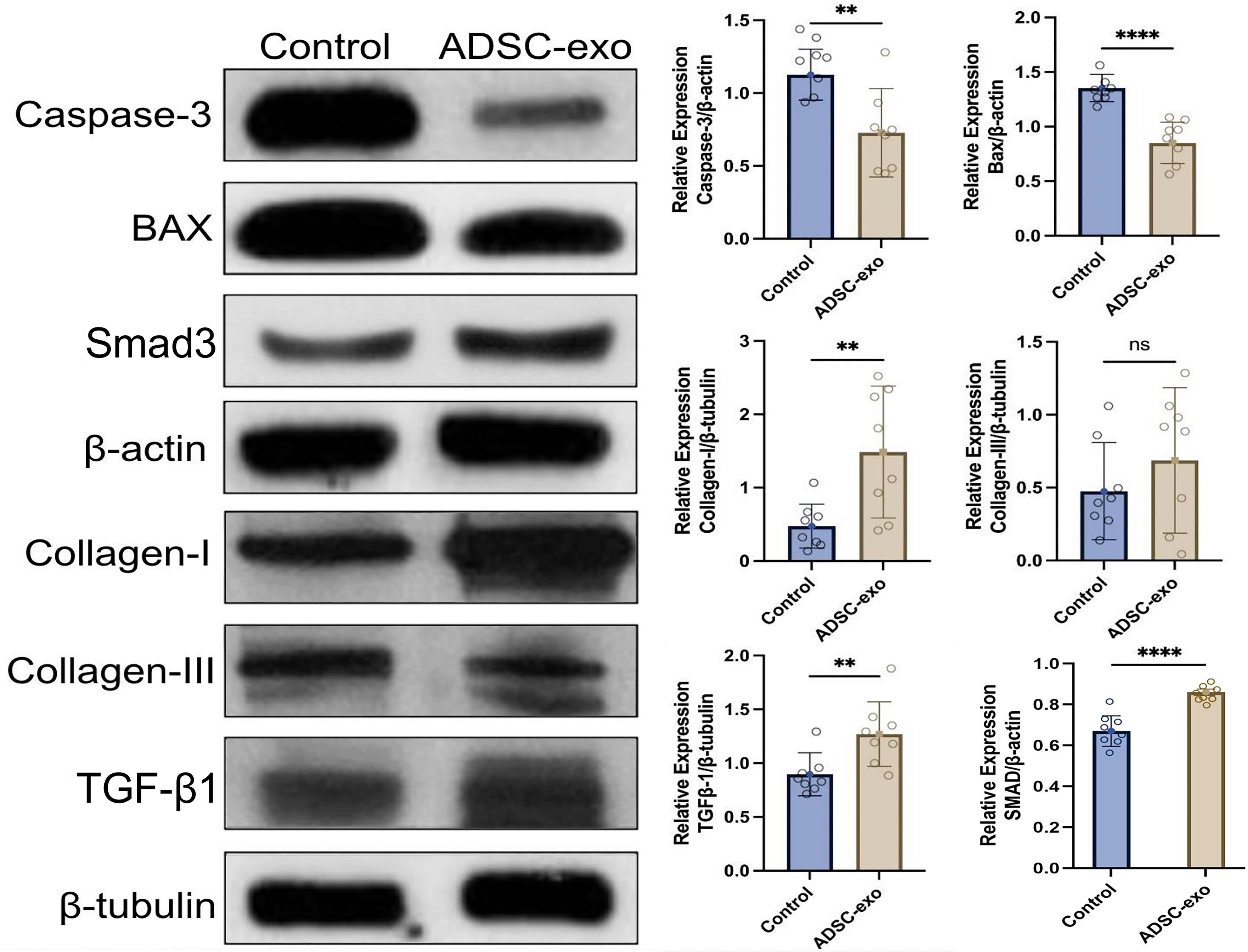
Protein expression analysis. Presentation of representative images and quantitative findings. Western blot analysis revealed significant differences in the average densitometry ratio of pro-apoptotic markers; caspase-3/β-actin was lower in the ADSC-exo group compared with the control (n = 8; 0.7 ± 0.3 vs. n = 8; 1.1 ± 0.2; p = 0.003), and the ratio for Bax/β-actin was lower in the ADSC-exo group compared with the control (n = 8; 0.9 ± 0.2 vs. n = 8; 1.4 ± 0.1; p < 0.0001). Collagen I/β-tubulin densitometry ratio was significantly elevated in the ADSC-exo group compared with control (n = 8; 1.5 ± 0.9 vs. n = 8; 0.5 ± 0.3; p = 0.005). No significant differences were observed in collagen III expression. TGF-β1/β-tubulin densitometry ratio was markedly higher in the ADSC-exo group compared with control (n = 8; 1.3 ± 0.3 vs. n = 8; 0.9 ± 0.2; p = 0.006). In addition, Smad3/β-actin densitometry ratio was significantly higher in the ADSC-exo group versus control (n = 8; 0.9 ± 0.02 vs. n = 8; 0.7 ± 0.1; p = 0.006). **p < 0.01, ****p < 0.0001.
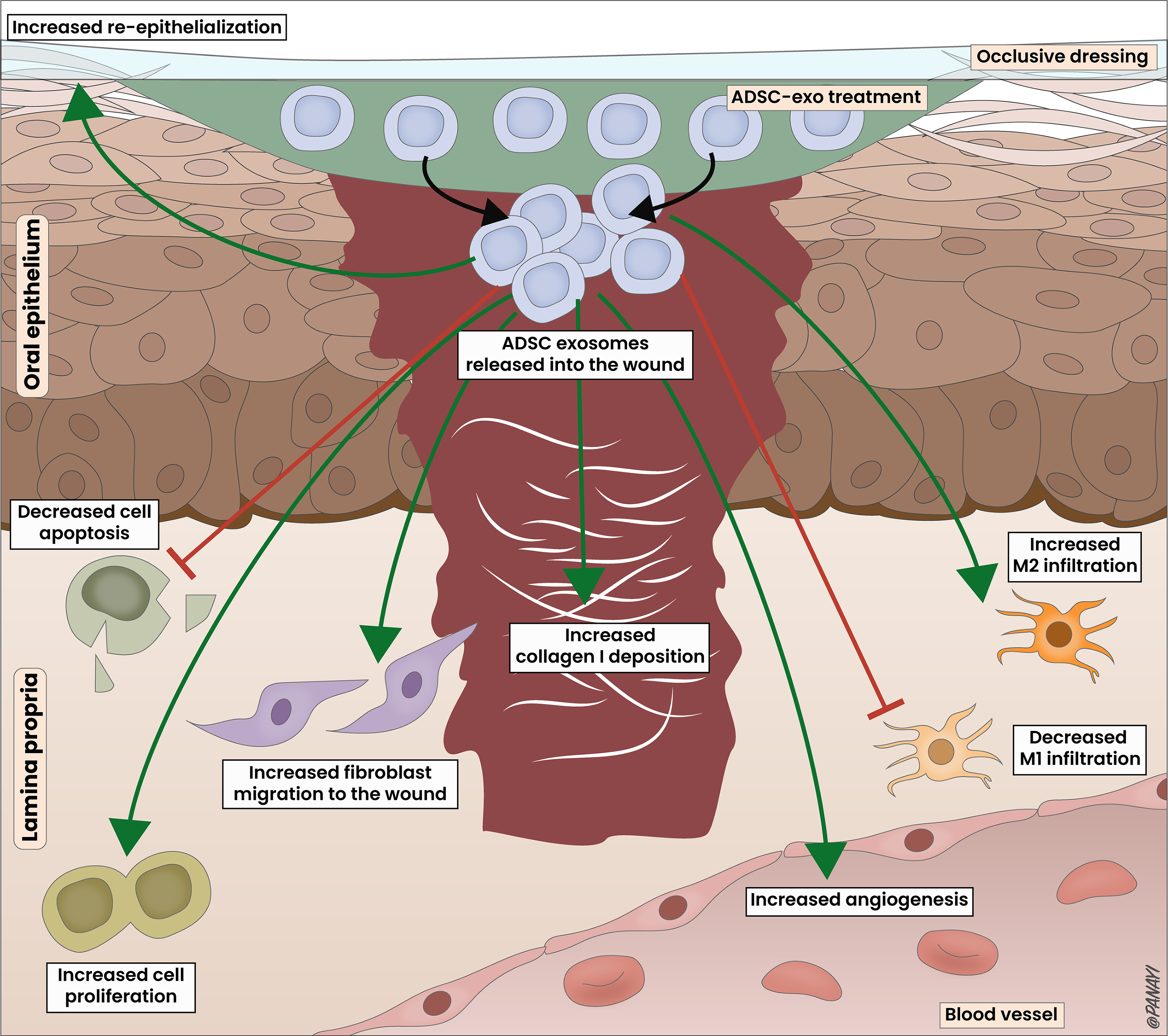
Proposed mechanism of action of ADSC-exo in oral mucosal wound healing. The pro-wound healing effects of ADSC-exo are multifaceted, involving the uptake of exosomes released by the ADSC-exo sheet into the wound. This uptake results in the upregulation of various factors, including TGF-β1 and Smad3, which subsequently enhances epithelialization, cellular proliferation, fibroblast migration, angiogenesis, and collagen I deposition. Simultaneously, it decreases apoptosis and influences macrophage polarization. This orchestrated response improves wound perfusion, promotes closure, facilitates extracellular matrix (ECM) remodeling, regulates inflammation, and ensures proper transition through the wound healing process.
DISCUSSION
To date, no research has been published investigating the effect of ADSC exosomes on oral mucosal wound healing. All prior investigations into ADSC exosomes have predominantly centered on cutaneous wound healing, revealing a myriad of pro-regenerative properties. 21 This study showed that using a Tisseel sheet containing ADSC exosomes (ADSC-exo) covered with a standard occlusive dressing on full-thickness hard palate mucosal wounds in rats improves the wound healing process, compared with using an occlusive dressing alone. Notably, ADSC-exo treatment expedited in vitro fibroblast migration, augmented cellular proliferation, promoted angiogenesis, enhanced collagen deposition and epithelialization, while concurrently mitigating apoptosis and regulating inflammation, within the wound microenvironment. This concerted response culminated in substantially accelerated wound closure rates in rat oral mucosal wounds, thus highlighting the potential of ADSC-exo in fostering effective and robust oral mucosal wound healing.
In accordance with the MISEV 2023 guidelines, 23 we used a range of techniques to confirm the presence of exosomes secreted from the ADSC-exo. The use of autologous ADSC exosomes in this study presents a remarkable advancement in mucosal wound healing therapies. Particularly noteworthy is that when contrasted with ADSCs, ADSC exosomes offer several advantages, such as circumventing ethical concerns, facile storage, and heightened stability, which includes improved traceability and a minimal risk of adverse reactions.19–21 Consequently, this strategy harbors substantial potential for clinical application, signifying a significant milestone in the endeavor to develop effective therapies for the management of mucosal injuries in human patients.
At the cellular level, our study revealed increased expression of the angiogenic microvessel endothelial marker CD31 in the group treated with ADSC-exo. This finding suggests a heightened density of blood vessels and perfusion of oxygen and nutrients within the wound environment.25–27 Moreover, enhanced oxygen perfusion has been associated with the reduction of reactive oxygen species activity, the promotion of leukocyte function, and improved wound healing.26–28
Our findings demonstrated that ADSC-exo is associated with a significant increase in fibroblast migration in the in vitro studies. Fibroblasts play a pivotal role in wound closure by synthesizing and depositing ECM components such as collagen, elastin, and fibronectin, which are essential for tissue integrity and strength.25,29 ADSC-exo may have also enhanced fibroblast migration in the in vivo studies, potentially contributing to the higher wound closure rate observed in that group.
ADSC-exo-treated wounds exhibited higher collagen I expression but not collagen III. The distinction between collagen I and collagen III expression in wound healing processes is significant and relates to the structural and functional properties of these two collagen types. Collagen I is the primary collagen found in mature scar tissue and provides tensile strength to the healed wound. In contrast, collagen III is prevalent in the early stages of wound repair and is gradually replaced by collagen I as the wound matures. 30 This suggests that ADSC-exo treatment may have specifically targeted the synthesis or deposition of collagen I without significantly impacting collagen III levels, or in other words, that it influenced the later stages of wound healing more than the early stages.
Previous research describes the inflammatory response to epithelial injury in three stages as follows: initial, peak, and declining. In the initial stage, M1 macrophages infiltrate the injury site, triggering an inflammatory cascade. 31 During the peak phase, M2 macrophages dominate, promoting anti-inflammatory processes and angiogenesis. 4 In the final stage, macrophage presence declines, and tissue remodeling occurs. 4 M1 macrophages tend to be linked to proinflammatory processes, while M2 macrophages suppress inflammation and promote healing. Wounds treated with ADSC-exo exhibited reduced pan-macrophage presence, but slightly increased M2 macrophage infiltration. Chronic and inadequate wound healing can partly stem from macrophages failing to transition into a pro-healing reparative phenotype, instead remaining in a pro-inflammatory and tissue-destructive state that hinders healing. 25 The diminished macrophage presence, coupled with increased M2 macrophage infiltration and accelerated wound closure in the ADSC-exo-treated group, implies that ADSC-exo may mitigate the pro-inflammatory phenotype, potentially fostering faster resolution of the wound.
ADSC-exo influenced cellular turnover by increasing proliferating cell activity, as indicated by elevated levels of the mitosis marker Ki-67, and suppressing the apoptotic expression of caspase-3 and Bax proteins. The balance between cellular proliferation and apoptosis directly impacts wound healing. Proliferating cells rapidly divide and replace damaged tissue, aiding in granulation tissue formation and wound closure. 25 Conversely, excessive apoptosis can hinder healing by depleting necessary cell populations. 32 Thus, maintaining a favorable ratio of proliferation to apoptosis is crucial for effective wound repair, ensuring the generation of new tissue and preserving essential cell types such as fibroblasts and keratinocytes.
Higher levels of TGF-β1 and Smad3 in ADSC-exo-treated wounds suggest that the therapeutic effects of ADSC-exo on oral mucosal wound healing may be intricately linked to its modulation of the TGF-β/Smad signaling pathway. TGF-β1 is released by macrophages, platelets, and fibroblasts during injury. Prior studies in cutaneous wound healing have underscored the significant contribution of TGF-β1 in the wound healing process.33–37 TGF-β1 activates receptor-regulated Smads (R-Smads), particularly Smad2 and Smad3, which play divergent roles in angiogenesis and tissue repair.33,34 Smad3 expression is associated with increased angiogenesis and enhanced microvascular density,38,39 as indicated by elevated CD31 levels in this study. Other studies have shown that Smad3 is crucial for fibroblast activation, driving α-smooth muscle actin expression and myofibroblast differentiation for wound contracture.25,33,34 Thus, the differences in magnitude for wound closure and reepithelialization in this study may be attributed to the differential effects of ADSC-exo on keratinocytes and fibroblasts, which are both crucial for the reformation of the protective epithelial barrier over the wound surface.
TGF-β1 has also been found to skew macrophage polarization toward an anti-inflammatory M2 phenotype through SNAIL upregulation,36–39 which, in turn, reduces pro-inflammatory cytokines and supports inflammation resolution in ADSC-exo-treated wounds. Furthermore, TGF-β1/Smad3 signaling is a key regulator of ECM remodeling, enhancing collagen I synthesis while suppressing collagen III production.25,36,37 Taken together, our results help outline a plausible mechanism of ADSC-exo in amplifying the TGF-β1/Smad pathway, a master regulatory pathway that facilitates cross talk among various cell types and induces pro-healing effects in a mucosal oral wound (Fig. 7).
Tisseel, a fibrin sealant composed of human fibrinogen and thrombin produced by Baxter Healthcare, is widely used in surgical procedures for hemostasis and has demonstrated the capacity to encapsulate therapeutic agents. 41 We selected Tisseel as the carrier for ADSC exosomes in this study due to its established efficacy in prior research. Recent studies have shown that a 20% TISSEEL-plasma-derived exosome product formulation can sustain EV release for up to 2 weeks. 42 Tisseel has also been shown to effectively deliver mesenchymal stem cells to accelerate wound healing, 43 and its combination with exosomes enhances cell migration and tissue remodeling.44,45 Notably, one study found that Tisseel has been shown to enhance vascularization and cellular proliferation in tendon healing on its own, however, its effects were significantly stronger when combined with platelet-rich plasma-derived exosome products. 46 This suggests that ADSC-exo in this study—composed of Tisseel and ADSC exosomes—may possibly work together synergistically. The use of Tisseel and ADSC exosomes in our study offers key clinical advantages, including FDA approval, cost-efficiency, and simplified production, eliminating the need for complex cell culture and manipulation.23,42
The study’s limitations include a small sample size, which may restrict the generalizability of findings and statistical power, and a lack of long-term follow-up beyond day 10 postsurgery, limiting insights into treatment durability and potential complications. Future studies should incorporate larger samples and longer time points (e.g., 30 days, 3 months) to assess the long-term scarring and stability of the healed wound. This would offer a more comprehensive understanding of the treatment’s clinical potential, particularly regarding final wound closure and tissue remodeling. In addition, while SD rats are commonly used, extrapolating findings to humans has inherent limitations, necessitating further research in larger animals or humans. The omission of wound marker measurements in normal palatal tissue was made to reduce animal use, in line with ethical guidelines. Investigations into specific cellular and molecular mechanisms and pathways underlying the effects of ADSC-exo were limited, highlighting the need for replication of this study and further investigation. While the Tegaderm-only control group served as an internal control in rat experiments, comparisons with undressed wounds could provide additional insights. Furthermore, the rapid regeneration of normal oral mucosa, especially with the occlusive dressing applied in both the treatment and control groups, may have reduced the ability to detect the effects of ADSC-exo in impaired healing conditions. Future research should focus on developing and applying ADSC-exo in models that simulate impaired oral mucosal healing, such as those mimicking diabetes or radiation-induced damage, as these models are currently lacking. Evaluating the broader applicability of ADSC-exo is crucial, as varying wound environments (e.g., mucosal, rectal, cutaneous excisional, incisional, and burn wounds) differ in dryness, fibrosis, and inflammation.
INNOVATION
The role of ADSC exosomes in oral mucosal wound healing is underexplored. In this study, we developed an innovative ADSC exosome sheet and assessed its therapeutic effects in a rat model. At the molecular, cellular, and functional levels, our results demonstrated that ADSC-exo significantly improved wound healing by accelerating tissue regeneration and enhancing wound closure. This work advances the field by providing a novel, clinically relevant therapeutic approach for oral mucosal injuries, potentially offering a new avenue for enhancing recovery in patients with oral wounds.
KEY FINDINGS
The mechanisms of action of an ADSC-exo in rat oral mucosa are elucidated. The ADSC-exo significantly accelerated wound closure in rat oral mucosal wounds. Application of ADSC-exo enhanced fibroblast migration speed. ADSC-exo promoted reepithelialization, cellular proliferation, and neovascularization while reducing apoptosis and regulating inflammation in the wound environment.
AUTHORS’ CONTRIBUTIONS
H.-K.K., C.-J.K., and D.Y.M. conceptualized and designed the experiments, performed the animal experiments, staining, molecular, and histology analysis, and acquired data. D.Y.M. carried out data analysis, interpretation of findings, and article writing. D.Y.M. and A.C.P. created the figures. H.-K.K., C.-J.K., D.P.O., and A.C.P. edited the article.
Footnotes
ACKNOWLEDGMENTS AND FUNDING SOURCES
This work was supported by grants from the
DATA AND RESOURCE SHARING AND AVAILABILITY
Detailed information on the resources used in this study, including the rat model, drugs, antibodies, and software, can be found throughout the article. Datasets generated and analyzed during the study are available from the corresponding author upon request.
AUTHOR DISCLOSURE AND GHOSTWRITING
The content of this study was exclusively contributed and written by the authors listed, and no ghostwriters were involved in the writing of this article. All authors confirmed no conflicts of interest, reviewed the article, and approved for submission.