
Abstract
Abstract
Zebrafish are increasingly used to study neurodegenerative conditions such as Parkinson's disease (PD). In rodents, the influence of the genetic background on important experimental parameters in PD research such as susceptibility to toxin exposure or motor behavior is well established. In contrast, little is known about the impact of the genetic background in commonly used zebrafish wild-type strains on these important experimental parameters. We determined the effect of the genetic background in five commonly used zebrafish wild-type strains on crucial, PD-related aspects, in particular the number of ascending dopaminergic neurons, their susceptibility to PD-related neurotoxins, and the expression levels of five genes involved in oxidative stress defense, protein degradation, cell death, and apoptosis. We also investigated whether the susceptibility to morpholino-mediated knockdown of the PD gene DJ-1 may have a varying effect on neuronal cell loss depending on the genetic background. Finally, we determined the influence of the genetic background on spontaneous motor behavior. There was remarkably little variation between the different wild-type strains for most parameters investigated. However, the susceptibility to the neurotoxin 1-methyl-4-phenylpyridinium differed between the five investigated strains and so did their spontaneous motor behavior.
Introduction
The role of genetic factors in both familial and sporadic PD is increasingly recognized. 2 The identification and subsequent molecular analyses of these genes have greatly improved our understanding of the pathogenesis of this disorder. Epidemiological studies also provide good evidence for an important role of genetic susceptibility factors in the pathogenesis of the considerably more common sporadic form of PD. 3 The most favored hypothesis is that distinct genetic variants confer increased susceptibility to exogenous or endogenous neurotoxins. Animal modeling is considered to be increasingly important in attempting to elucidate gene–environment interactions. 4
The zebrafish has recently emerged as a new and powerful vertebrate animal model to study human diseases. 5 Its usefulness for the study of neurodegenerative diseases is particularly noteworthy. 6 We and others have, therefore, adopted the zebrafish as a new model for PD.7–11 The dopaminergic system is well characterized in both embryonic and adult zebrafish. 12 Dopaminergic neurons are first detected between 18 and 19 hours postfertilization (hpf) in a cluster of cells in the posterior tuberculum of the ventral diencephalon. 13 Retrograde labeling studies suggest that the diencephalic neurons in groups 1, 2, 4, and 5 represent the dopaminergic system ascending to the striatum, comparable to the nigrostriatal system in humans.14,15 A more recent study (published after completion of our study) has cast some doubt on this. 16 However, all studies using zebrafish as an animal for PD have reported specific vulnerability of these dopaminergic cell clusters to PD neurotoxins (see below) or PD gene knockdown regardless of the precise projection of these dopaminergic neurons.8–10
The number of dopaminergic neurons and their susceptibility to neurotoxins in mice is strongly influenced by their genetic background. 17 Hydrogen peroxide, the proteasome inhibitor MG132, and 1-methyl-4-phenyl-1,2,3,6-tetrahydropyridine (MPTP) are frequently used neurotoxins for the investigation of the different mechanisms leading to neuronal cell death in PD. Hamre and co-workers investigated seven different mice strains for varying susceptibility to MPTP and found a remarkable range of neuronal cell loss from 63% cell loss in C57BL/6J mice to 14% cell loss in Swiss Webster mice, after standardized MPTP exposure. 18 Intercrossing of two different mice strains with high and low MPTP susceptibility, with subsequent assessment of dopaminergic cell loss in the SN accompanied by genome-wide linkage analysis, allowed the mapping of quantitative trait loci (QTL) for MPTP sensitivity.19,20 Previous studies have suggested that there may be a sufficient degree of genetic variability between some zebrafish strains to undertake similar studies. 21 Strain-dependent susceptibility to MPTP or indeed any other neurotoxin implicated in the pathogenesis of neurodegenerative diseases such as hydrogen peroxide or proteasome inhibitors have, however, never been investigated in this model organism.
The effect of the genetic background on the behavioral phenotype in mammals and rodents has long been recognized as well.22–25 Marked differences between different mice strains have been reported in spontaneous locomotor activity, coordination ability, and also swim speed as well as the number of fore- and hind-paw paddles required to swim the length of a tank.26,27
In contrast, all studies investigating the effect of PD neurotoxins or PD gene knockdown in zebrafish have ignored the potential impact of variation in genetic background on spontaneous motor behavior, gene expression levels, number of dopaminergic neurons, susceptibility to neurotoxins, and the effect of gene knockdown.
The aim of our study was therefore to systematically determine the effect of variation in the genetic background between five commonly used zebrafish wild-type strains on number of dopaminergic neurons, susceptibility to PD neurotoxins, expression levels of crucial genes frequently altered in neurodegenerative diseases, and spontaneous motor behavior. We also determined whether genetic background influences the effect of morpholino-mediated antisense knockdown of the zebrafish homologue of DJ-1 on the number of dopaminergic neurons. Autosomal recessively inherited mutations in DJ-1 lead to early onset PD in human patients, but the precise mechanisms leading to neuronal cell death are still poorly understood. 2
Materials and Methods
Zebrafish maintenance
LWT, AB, EKK, WIK, and TL zebrafish were maintained using standard procedures in accordance with UK Home Office animal welfare guidelines. Embryos were obtained by natural mating and synchronously raised at 28.5°C.
Cell count of dopaminergic neurons
Digoxygenin-labeled antisense RNA probes were synthesized from pBS-TH plasmid linearised by XhoI and transcribed using T3 RNA polymerase. Whole-mount in situ hybridization of zebrafish embryos was performed at 72 hpf as previously described. 7 The ascending TH positive diencephalic neurons in the groups 1, 2, 4, and 5 according to the classification by Rink and Wullimann were counted only.14,15 For each strain the results for the toxin exposure experiments are expressed in percentage of the neurons present in unexposed zebrafish embryos belonging to the same wild-type strain.
Toxin exposure
Embryos at 24 hpf were transferred in groups of 20 into 12-well plates containing 2.5 mL of the neurotoxin at the following concentrations: H2O2 2.5 mM; MG132 50 μM; and 1-methyl-4-phenylpyridinium (MPP+) 6 mM (all from Sigma). They were exposed for 24 h to H2O2 and for 48 h to MPP+ and MG132. The dose of MG132 was based on previously published experiments. 7 The dose of MPP+ was based on the results of unpublished pilot studies using a range of different MPP+ concentrations. MG132 was dissolved in 0.2% dimethyl sulfoxide, which was used as control vehicle, and E3 was used as the control vehicle for H2O2 and MPP+. 1-Phenyl 2-thiourea (SIGMA) was used at the concentration of 0.21 mM to prevent the development of pigmentation in all experiments. Embryos were fixed overnight at 4°C in 4% paraformaldehyde and stored at −20°C in methanol.
Quantitative gene expression analysis
Quantitative RT-polymerase chain reaction (PCR) was undertaken using SYBR Green PCR Master Mix (Applied Biosystems) and the MX3000P Real-time PCR System (Stratagene), following a previously described protocol. 28 Expression of SOD1, catalase, Hsp70, Bax, and p53 was analyzed at 3 days postfertilization (dpf) as previously described. 7 Gene expression levels were normalized to β-actin gene expression. The expression levels for the five different genes relative to those detected in LWT embryos are expressed as percentages.
Morpholino injections
The following two MOs (Genetools, LLC) were designed against the splice-donor sites of exons 4 and 5 of DJ-1: 5′-CCAAATGAAGATT
Measuring locomotor activity
Movement analysis of untreated zebrafish (5 dpf) from the five WT strains was undertaken using the semiautomatic video-tracking and motion analysis system Etho Vision Pro 3.1 (Noldus). The larvae were raised in Petri dishes until 5 dpf and then transferred individually to 12-well plates and allowed to adjust to the conditions for 5 min. Their spontaneous locomotor activity was recorded for 5 min under infrared light at a defined light setting and analyzed for the total distance swum (cm). There were between 10 and 15 zebrafish larvae for each experiment in each of the tested groups/wild-type strains, and experiments were repeated three to five times. The behavioral analysis was always carried out between 9.00 and 11.00 AM. The experiments were undertaken in a quiet, darkened room without any other extraneous stimuli.
Statistical analysis
A minimum of 10 zebrafish larvae per group was analyzed in each experiment. All experiments were performed in triplicate unless otherwise stated. Data represent the mean ± standard error of the mean. For multiple comparisons, significant differences were tested first using single-factor analysis of variance (ANOVA) followed by Tukey post-hoc test (GraphPad Prism).
Results
The number of dopaminergic neurons was somewhat lower in TL and WIK (WIK 33.97 ± 1.64, TL 33.53 ± 2.20) than in the other investigated strains (LWT 37.74 ± 1.74, AB 36.83 ± 1.12, EKK 36.47 ± 1.98), but this difference did not reach statistical significance (p > 0.05). There was no significant decrease in the number of the dopaminergic neurons in any of the investigated wild-type strains after MO-mediated knockdown of the PD gene DJ-1 (p > 0.05; Fig. 1).
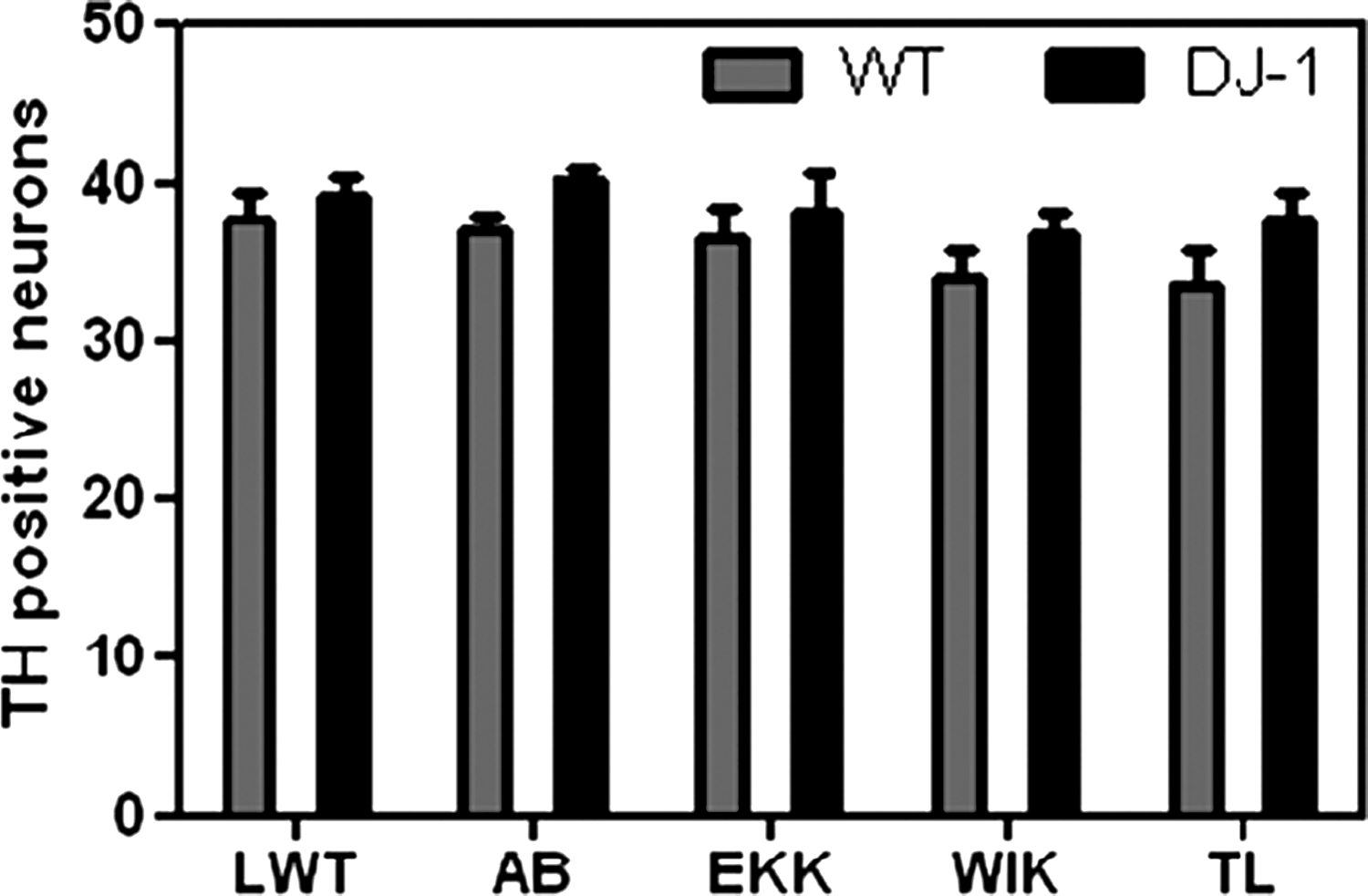
Number of dopaminergic neurons in the posterior tuberculum with and without Morpholino-mediated knockdown of the Parkinson's disease gene DJ-1 in LWT, AB, EKK, WIK, and TL. Dopaminergic neuronal cell count was neither significantly different between the different untreated wild-type strains (gray bars) nor in the Morpholino-injected embryos (black bars). The number of dopaminergic neurons was counted manually after whole mount in situ hybridization with an RNA probe for tyrosine hydroxylase.
Exposure to the complex I inhibitor MPP+ at a concentration of 6 mM induced a significant decrease of DA neurons in all 5 strains (23%–37%). TL zebrafish larvae had a significantly increased susceptibility to MPP+ compared to LWT with a mean TH neuronal cell count in TL of 34.70 ± 0.62 before and 22.20 ± 1.27 (63.95% ± 3.19%) after MPP+ exposure compared to 35.23 ± 0.44 before and 27.10 ± 0.68 (76.9% ± 2.22%) after MPP+ exposure in LWT (p < 0.01, using one-way ANOVA and Tukey post-hoc testing) (see Fig. 2a).

Susceptibility of the five investigated wild-type strains (LWT, AB, EKK, WIK, and TL) to neurotoxins at 3 days postfertilization (dpf). Embryos were exposed for 48 h to the complex I inhibitor 1-methyl-4-phenylpyridinium (MPP+, ** for p < 0.01)
There was no significant difference between the different wild-type strains in their susceptibility to the proteasome inhibitor MG132 and the pro-oxidant H2O2 (see Fig. 2b, c).
Gene expression levels were very similar for Catalase, SOD-1, Bax, p53, and Hsp-70 in the five different wild-type strains at 3 dpf (Fig. 3a–e).

TL and EKK larvae showed less spontaneous motility than WIK (distance swum for TL: 49.26 ± 4.59 cm, distance swum for EKK: 47.39 ± 5.70 cm compared to WIK 75.42 ± 1.63 cm, p < 0.05, Fig. 4).
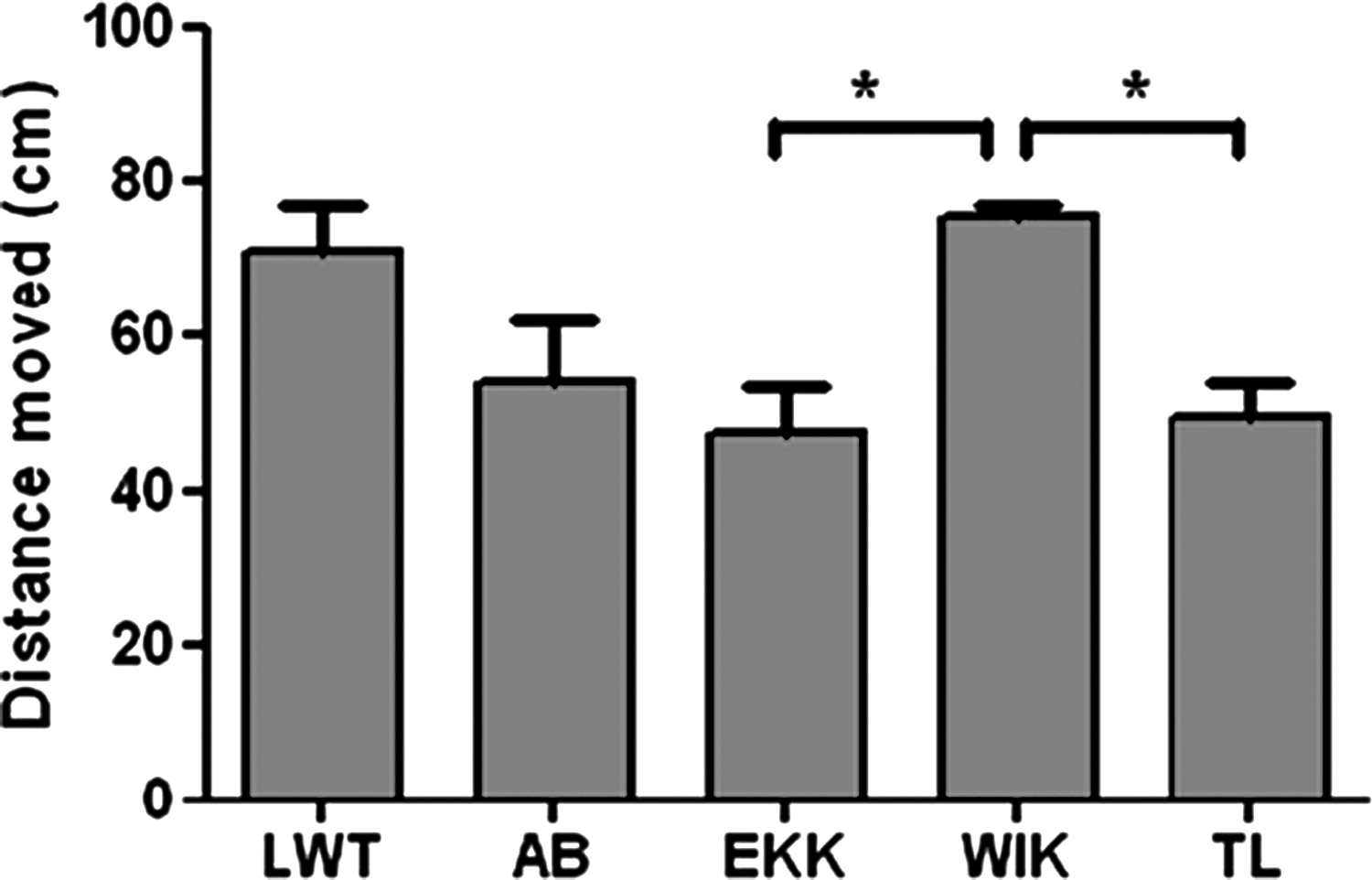
Spontaneous motor activity for LWT, AB, EKK, WIK, and TL. Locomotor activity was recorded in 5 dpf larvae for 5 min. Experiments were repeated at least three times, each group contained 12–24 larvae per experiment. The distance swum by WIK was significantly longer (75.42 ± 1.63 cm) cm than the distance covered by EKK (47.39 ± 5.70 cm) or TL (49.26 ± 4.59 cm). *for p < 0.05.
Discussion
We have investigated the influence of the genetic background on DA neuronal cell count, susceptibility to neurotoxins, gene expression, effect of PD gene knockdown, and spontaneous motor behavior in five different, commonly used zebrafish wild-type strains. We observed an influence of the genetic background on the susceptibility to one particular neurotoxin, MPP+, but not for the other toxins tested. Our data also suggest a difference in spontaneous motor activity between the different wild-type strains.
Our previous work showed the lack of an effect of DJ-1 knockdown on the loss of dopaminergic neurons in one particular zebrafish wild-type strain, namely, LWT. 7 We therefore hypothesized that it may be the particular genetic background of LWT that would protect the dopaminergic neurons against any deleterious effect of DJ-1 deficiency. However, the data presented in this study suggest that the lack of an effect of DJ-1 deficiency on the number of dopaminergic neurons is not limited to the LWT strain, but rather a shared feature amongst all the different wild-type strains analyzed.
Strain-dependent susceptibility to ethanol in zebrafish has previously been reported by several different groups.29–31 We have previously demonstrated that DJ-1 knockdown in LWT results in increased susceptibility to the proteasome inhibitor MG132. 7 We therefore hypothesized that there may be genetic variants in at least some zebrafish wild-type strains that would confer a similar increase in susceptibility to MG132. This might have allowed subsequent QTL mapping leading to the possible identification of a previously unidentified genetic susceptibility factor for PD neurotoxins. However, susceptibility to MG132 was largely similar in all the different wild-type strains tested. Higher doses of MG132 lead to high, nonspecific mortality (data not shown).
Our previous study revealed a distinct pattern of gene expression changes in DJ-1 knockdown zebrafish embryos for superoxide dismutase, catalase, p53, Bax, and Hsp70. 7 We therefore hypothesized that any change to the previously observed gene expression pattern might explain a difference in the sensitivity to DJ-1 knockdown across the different strains. However, as mentioned above, the effect of DJ-1 knockdown was largely the same in all wild-type strains tested. Further, the susceptibility to two of the three neurotoxins tested was also very similar. The lack of a difference in gene expression levels is thus in keeping with the similar susceptibility to PD gene knockdown and toxin susceptibility (other than to MPP+).
An influence of the genetic background on complex behavior such as aversion, learning and memory is also well established. 32 Life span varies between outbred zebrafish and at least one particular inbred strain, golden sparse, suggesting that longevity in zebrafish is also influenced by the genetic background. 33 We were thus surprised to see that the overall influence of the genetic background appears to be considerably smaller for any of the paradigms tested in our study compared to the marked differences between different strains in other animal models such as rodents (see Introduction section). The average nucleotide diversity in zebrafish dramatically exceeds that observed in the human genome by an order of magnitude and is four times higher that that in commonly used rat strains or between two mouse subspecies. 34 There is, however, also considerable intrastrain variation. 34 This may at least partially be due to the fact that most zebrafish originate as outbred stocks and at least some of them are also maintained in this way. Thus, the relatively weak effects of genetic background on the experimental parameters tested in our study may be due to the fact that it is less stable in zebrafish than in other animal models. QTL analysis might therefore be possible for certain behavioral characteristics, but is unlikely to reveal loci with increased susceptibility to PD neurotoxins or any of the other, PD-relevant parameters. 35
Footnotes
Acknowledgments
Financial support from Parkinson's UK (G-0608; G-0901), the Sheila McKenzie Fund (University of Sheffield, UK), and Medical Research Council (MRC Centre Grant to P.W. Ingham) is gratefully acknowledged.
Disclosure Statement
No competing financial interests exist.