
Abstract
Abstract
Zebrafish (Danio rerio), is one of the most extensively studied chordate model animal in the field of neuroscience and development biology. Understanding the proteome profile of various tissues and organs of Danio rerio has become increasingly important for inching it as an alternate translational model animal. This study aimed in understanding the proteome profile of D. rerio brain's olfactory bulb exclusively. The proteome pattern of olfactory bulb of D. rerio was determined using the two-dimensional gel electrophoresis approach involving both full range and three various narrow pH range immobilized pH gradient strips, followed by mass spectrometry and tandem mass spectrometry analysis. 221 various proteins were characterized based on matrix-assisted laser desorption/ionization mass spectrometry and tandem mass spectrometry analysis as olfactory bulb–specific proteins. The identified proteins represented various pI, mass, functions, and localization. The different proteins acknowledged in this study were found to be associated with diverse metabolic pathways, enzymatic activities, and neurological functions. Glycolysis, gluconeogenesis, and oxidative phosphorylation are the major metabolic pathways found associated with the identified proteins. The identified protein catalogue was also found to be participating in various network pathways such as GABAergic neurophysiological neurotransmission, remodeling of cytoskeleton, neurofilaments, and cell adhesion. This study provides the D. rerio olfactory bulb two-dimensional gel electrophoresis proteome map and the details of 221 olfactory bulb–specific proteins.
Introduction
The central nervous system (CNS) is the most complex system among vertebrates for its intricate structure and function. The CNS of D. rerio includes a spinal cord and compound brain. The compound brain is made of forebrain (telencephalon including olfactory bulb), midbrain (mesencephalon), and hindbrain (metencephalon including cerebellum).8,9 Understanding the complexity of the CNS based on embryonic growth has given extensive clues regarding the development of the complex structure.10–12 Recent studies on understanding the various proteins expressed in the brain tissue in basal conditions based on two-dimensional gel electrophoresis matrix-assisted laser desorption/ionization MS/MS analysis and liquid chromatography mass spectrometer analysis13,14 have given a better view of the atlas of the proteome expression pattern and can be used as a reference tool for biomarker discovery and understanding the protein level changes for various neurological studies.
Olfactory bulb is one of the vital parts of the brain having dynamic neurogenesis and neuronal migration processes. Danio rerio olfactory bulbs are found in the limbic region of the forebrain and are mostly involved in olfaction and perception of odors through neurogenesis and neuronal migration. Main and accessory olfactory bulb are the two discrete structures of D. rerio olfactory bulbs associated in relaying the sensory signals and sense of smell to the olfactory tract. Understanding the proteome map of olfactory bulbs has a high impact in estimating the major biological functions associated with olfactory bulb in D. rerio. The proteome profile of mature olfactory bulb of rat, 15 differential expression of proteins between neural progenitor cells from the adult rat brain subventricular zone and olfactory bulb, 16 and redox proteomics and cellular localization of oxidative stress in the aging murine olfactory bulb 17 are the notable studies performed for understanding the proteome of olfactory bulbs. This study aims specifically to identify and map all the proteins found in the olfactory bulb of the adult D. rerio brain in its native state.
Materials and Methods
Animal and sample preparation
Adult male and female D. rerio (Danio rerio) were collected from breeding colonies 18 and were anesthetized for 10 min in 0.1% Tricaine (Sigma US; www.sigmaaldrich.com). Mature olfactory bulbs were carefully dissected from 50 animals and were washed twice in Locke-Ringer's solution (0.9% NaCl, 0.042% KCl, 0.024% CaCl2, 0.02% MgCl2, 0.05% NaHCO3, and 0.05% dextrose (w/v) in Milli-Q). The tissues were pooled and homogenized in liquid nitrogen for the extraction of total olfactory bulb proteins using dissolving buffer (7 M urea, 2 M thiourea, 4% 3-[(3-cholamidopropyl)dimethylammonio]-1-propanesulfonate, 18 mM Tris-HCl pH 8, 14 mM trizmabase, 2 tablets EDTA-free protease inhibitor, 0.2%Trition X, 50 mM dithiothreitol). The concentration of the total protein extract was quantified using an amido black protein assay protocol with bovine serum albumin as standard. 18
Isoelectric focusing (IEF) and second-dimensional gel electrophoresis
First-dimensional isoelectric focusing (IEF) of the olfactory bulb protein was performed using 600 μg of total protein on both broad-range immobilized pH gradient (IPG) strips (pH 3–10 NL, 13 cm) (GE Healthcare; www.gehealthcare.com) and three various narrow range IPG strips (pH 3–6, 5–8, and 7–10, 11 cm) (BioRad; www.bio-rad.com). After overnight passive rehydration of the protein in IPG strips, the IEF for broad and narrow range IPG strips was performed at 20°C with 50 μA current using the following two different IEF programs respectively,100 Volts for 90 min; 300 Volts for 2 h; 500 Volts for 2 h; 1000 Volts for 2 h; 8000 Volts for 120,000 Volt hours; 500 Volts for 10 h, and 250 Volts for 30 min; 1000 Volts for 90 min; 4000 Volts for 40,000 Volt-hours; 500 Volts for 10 h. Equilibration of the strips were performed in buffer I (6 M urea 0.375 M Tris-HCl (pH 8.8), 2% sodium dodecyl sulfate, 20% glycerol, 2% dithiothreitol), and buffer II (buffer I with 2.5% iodoacetamide instead of dithiothreitol). The second-dimensional gel electrophoresis was performed in a 12% sodium dodecyl sulfate-polyacrylamide gel overnight at 15 mA. The gels were stained for fast Coomassie staining by boiling for 2 min and 4 h at room temperature incubation in a series of three solutions, solution A (50 mg Coomassie brilliant blue R250, 25 ml isopropanol, 10 ml acetic acid, and 65 ml water), solution B (50 mg Coomassie brilliant blue R250, 10 ml isopropanol, 10 ml acetic acid, 80 ml water), and solution C (2 mg Coomassie brilliant blue R250, 10 ml acetic acid, 90 ml water). Following this, gels were de-stained in 10% acetic acid for 2 min (boiling state) and 24 h (room temperature). Further, gels were scanned and documented using BioRad versadoc photo imager (www.bio-rad.com) and analyzed using IMP (GE Healthcare; www.gehealthcare.com) software. The experiments were repeated thrice for both broad and narrow ranges separately to confirm the spot pattern.
Matrix-assisted laser desorption/ionization (MALDI) analysis
Prominent protein spots having more than 0.01% spot volume were manually excised from the gel and processed for the MALDI–tandem mass spectrometry (MS/MS) analysis. The excised gel pieces were destained twice for 1 h with 50% acetonitrile (ACN)/25 mM ammonium bicarbonate, washed twice for 30 min in sterile water, dehydrated (100% ACN wash for 15 min), and dried. The dried gel pieces were digested using 120 ng trypsin (Promega) reconstituted in 40 mM ammonium bicarbonate and 10% ACN for 16 h at 37°C. The digested samples were vortexed for 1 h in 100 μl of 5% trifluoroacetic acid (TFA)/50% ACN twice, and the supernatant was collected in a fresh microfuge tube and lyophilized. The lyophilized contents were reconstituted in 5 μl of 0.1% TFA. The peptide mass fingerprinting (PMF) data for the MALDI-TOF-MS (matrix-assisted laser desorption/ionization time-of-flight mass spectrometry) assay was performed by mixing 1 μl of peptide mix and the matrix solution, α-cyano-4-hydroxycinnamic acid (CHCA, 5 mg/ml) in 4800 plus MALDI TOF/TOF system (ABI; www.appliedbiosystems.com). Standard peptides spanning a mass range of 800–4000 m/z were used to calibrate the instrument for less than 10 ppm accuracy. Further, the PMF data were acquired in the reflector mode with an accelerating voltage of 25 kV. The preliminary protein IDs along with carbamidomethylation of cysteine modification, methionine oxidation, and up to one missed cleavage were screened for the obtained PMF data using GPS software version 3.6 (ABI; www.appliedbiosystems.com) against the Danio rerio database (2008) and Swiss-Prot 2009 (MASCOT version 2.0 search engine; www.matrixscience.com). The identified proteins were further analyzed for similarity between the theoretical pI and mass against the protein spots as seen in the gel. The MS/MS of each protein were further confirmed from a minimum of three peptide precursors for each protein from the PMF list and fragmented in the CID tube by high laser. The MS/MS were also analyzed as mentioned for MS spectra in the MASCOT database.
Data analysis
The protein identities obtained from the D. rerio olfactory bulbs were analyzed and classified for its pI, molecular weight and localization. Based on DAVID (Data base for Annotation Visualization and integrated discovery; www.david.abcc.ncifcrf.gov) analysis, the protein details were annotated and classified 20 for function, process, and cellular component localization. The proteins identified from the D. rerio olfactory bulbs were also analyzed for their association in various processes, networks, pathways maps, and diseases network using GeneGo software (www.genego.com). The various pathway maps and the associated pathways were obtained from the GeneGo portal upon analysis against the protein identities. Also the obtained 2DE gel images and the details of all the protein spots identified in the study were submitted to the world-2DE PAGE portal (http://world-2dpage.expasy.org/repository/) and a repository accession number was obtained.
Results and Discussion
Two-dimensional gel electrophoresis (2-DE), MALDI and data analysis
A total of 289 spots having more than 0.01% spot volume were picked from all the three narrow ranges 2-DE gel reproducibly and unambiguously. Based on MALDI analysis, a total of 221 proteins were identified and mapped as olfactory bulb proteome, which includes 153 proteins characterized both with MS and MS/MS analysis (Supplementary Table S1; supplementary data are available online at www.liebertonline.com/zeb). All the spots selected for the analyses are highly resolved and are single protein spots. 30% of the proteins were identified from 2-DE gel of pH range 3–6, followed by 65% from pH 5–8 and 5% from pH 7–10 (Fig. 1). The cumulative pattern of all the protein spots identified from the three different narrow ranges were also reproduced and confirmed in the broad range pattern of pI 3–10 (Fig. 1). The identified proteins embody a wide range of pI ranging from 4.09 to 9.25; 66% of the identified olfactory bulb proteins were found in the pH range 5 to 7 (Fig. 2a). The largest and smallest proteins identified from the olfactory bulbs are spectrin alpha 2 (285 KDa) and transcriptional regulator, XRE family protein (8 KDa) respectively (Supplementary Table 1), with 75% of the identified proteins representing the mass range of 20 to 60 KDa (Fig. 2b). The proteins identified from the olfactory bulb were found localized evenly in the cell cytoplasm (39%), nucleus (26%), mitochondria (9%), and membranes (8%) (Fig. 2c). Based on the functional distribution, all the identified proteins were mostly associated with binding and enzymatic activity. Thirty-six percent of the proteins were involved in binding activities followed by 34% enzymatic and 12% with transporting activities (Fig. 2d). The dataset of the proteins identified from the 2DE gels are accessible via the World-2D PAGE repository portal (Accession No. 0039).
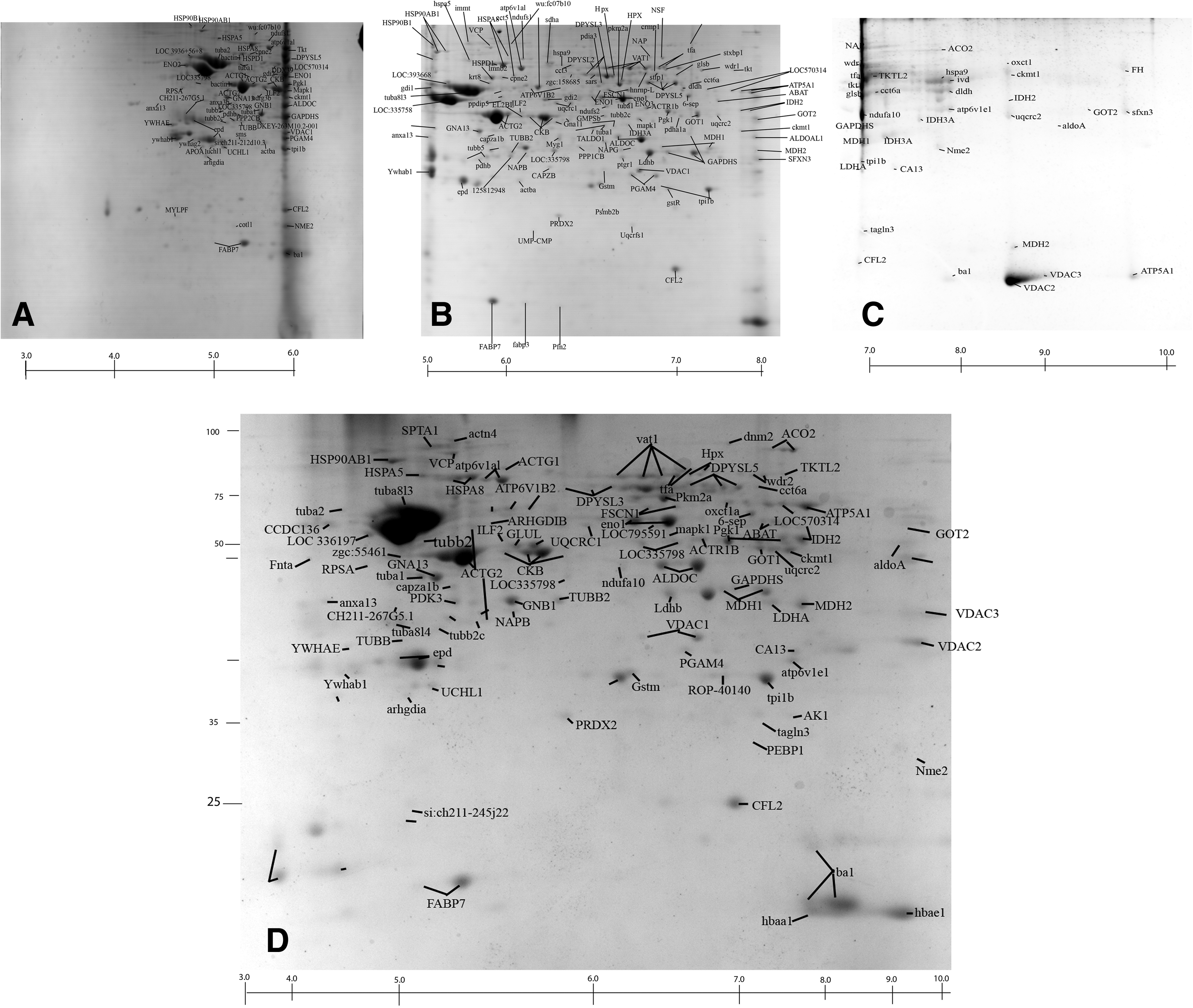
Two-dimensional gel electrophoresis profile of zebrafish olfactory bulb protein stained with fast Coomassie staining. Proteins were separated according to the pI gradient (x-axis) in the first dimension and based on molecular weight in the second dimension (y-axis). 221 spots were identified and analyzed by tandem mass spectrometry (MS/MS).
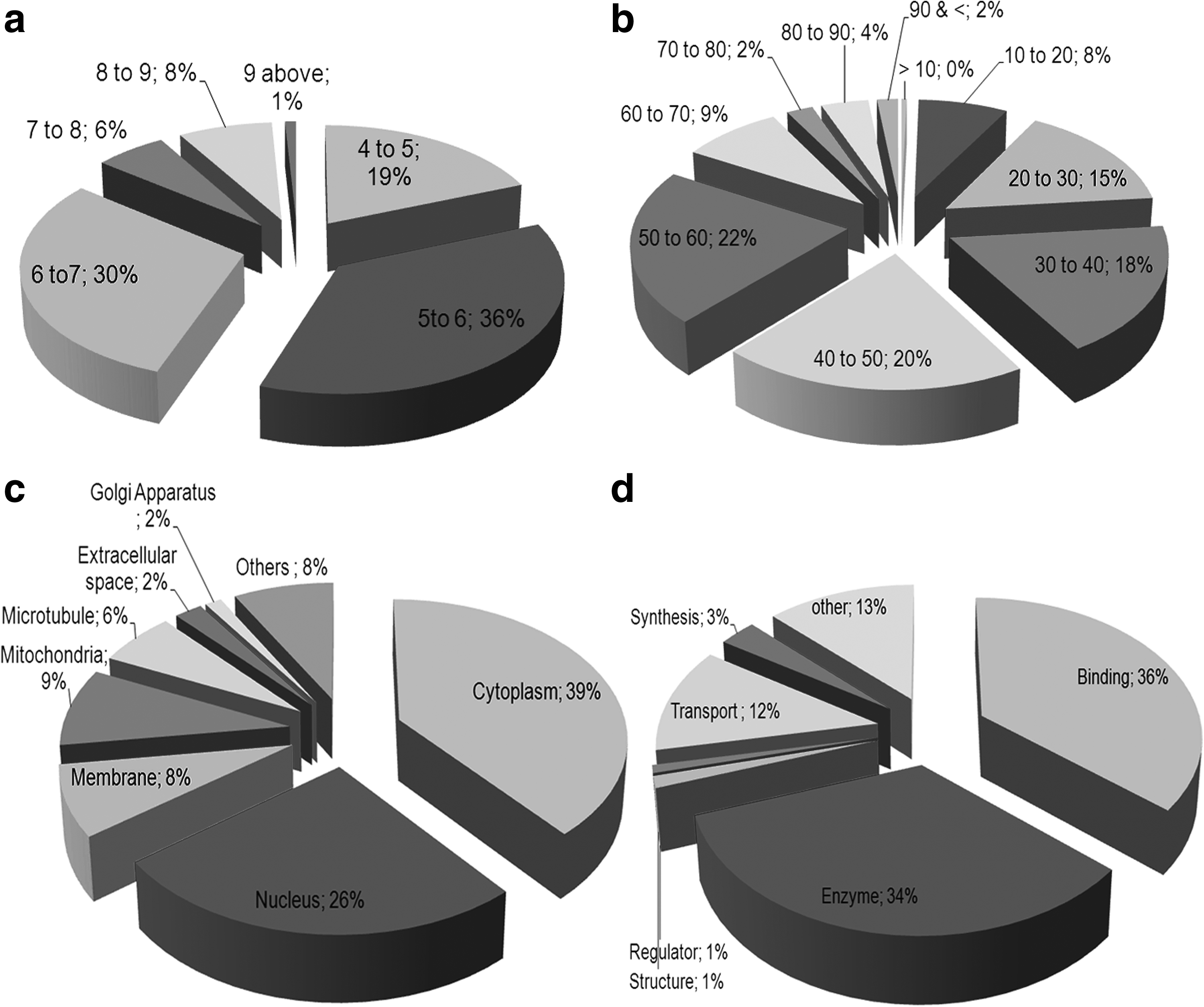
Pie chart distribution of protein identities based on
Dataset classification and function
The D. rerio olfactory bulb proteome identified from this study was found to be associated with a wide range of biological activities such as metabolism, neurophysiological activity, cellular processes, and signaling pathways. The various functions associated with the identified proteins as per DAVID analysis and Gene Ontology molecular function search are nucleotide binding (21 proteins), catalytic activity (19 proteins), protein binding (17 proteins), pyrophosphatase activity (14 proteins), hydrolase (13 proteins), and GTPase activity (12 proteins) (Fig. 3).

Functional classification of the zebrafish olfactory bulb proteins. The bar indicates the association of proteins to different functions (log bar). The classification is based on GeneGo (www.genego.com) process classification. X-axis represents the log p value of calculated hypergeometric distribution statistics of protein association (based on GeneGo analysis).
Pathway network analysis for process based on GeneGo analysis selected 176 D. rerio olfactory bulb proteome identities for analysis. The major functional pathway maps associated with the D. rerio olfactory bulb proteome are energy metabolism and its regulation (36 proteins), degradation and catabolism (16 proteins), disease association (25 proteins), neurotransmission (20 proteins), and calcium signaling (12 proteins). The major metabolic pathway maps identified from the dataset based on GeneGo analyses are glycolysis (Fig. 4), gluconeogenesis (Fig. 4), and oxidative phosphorylation. Aldolase A, aldolase C, aldehyde dehygrogenase 5 family member A1, phosphoglycerate kinase 1, biphospho glyceromutase-1, 4, enolase-1, 2, 3, glyceraldehyde-3-phosphate dehydrogenase, malate dehydrogenase cytoplasmic, NADH dehydrogenase, NADH dehydrogenase NADH dehydrogenase succinate dehydrogenase, lactate dehydrogenase A4, lactate dehydrogenase B4, and MDH mitochondrial are few of the major identified proteins involved in the different metabolic pathways. The glycogenolysis (Fig. 4) metabolic pathway is the most significant process obtained from the dataset involving 11 major proteins (enzymes) of the proteome dataset. The important biological pathways obtained from the proteome list are tricarboxylic acid cycle involving FUMH, ACON, MDH1, IDH3A, AATM, IDH2, IDH2, MDH2, SDHA, and AATC, and glutamate signaling pathway involving KCRU and VDAC1 proteins. The various biomarkers associated diseases found allied with the D. rerio olfactory bulb proteome are schizophrenia, mental disorders, psychiatry, dementia, Alzheimer's disease. 55 olfactory bulb proteins identified in this study were found to be associated as network objects in the biomarker map of schizophrenia disease, a severe emotional disorder of psychotic depth characteristically marked by a retreat from reality with delusion formation.

Process network association for glycogenolysis pathway. 11 proteins (enzymes) were represented from the study for the glycolysis and gluconeogenesis directly. The associated proteins in the pathway are represented with thermometer cartoon.
The neurophysiological process involving GABAergic neurotransmission (Fig. 5) is the major neurotransmission pathway obtained as neural process from the protein dataset. CALM2, MUNC18, Dynamin, GABT, MDH1, MDH2, AATC, and PP1-cat are few of the 12 important proteins of the D. rerio olfactory bulbs that were found to be associated in the process of GABAergic neurotransmission, the most abundant inhibitory neurotransmitter found in the mammalian brain. 21 Remodeling of cytoskeleton, neurofilament remodeling, and cell adhesion for histamine receptor are other major functional pathway maps associated with the identified proteins. ARP1 actin-related protein 1 homolog B and syntaxin binding protein 1, along with actin and tubulin complex, are found to be involved in the cytoskeletal remodeling of neurofilaments, the primary intermediate filament type expressed by neurons.
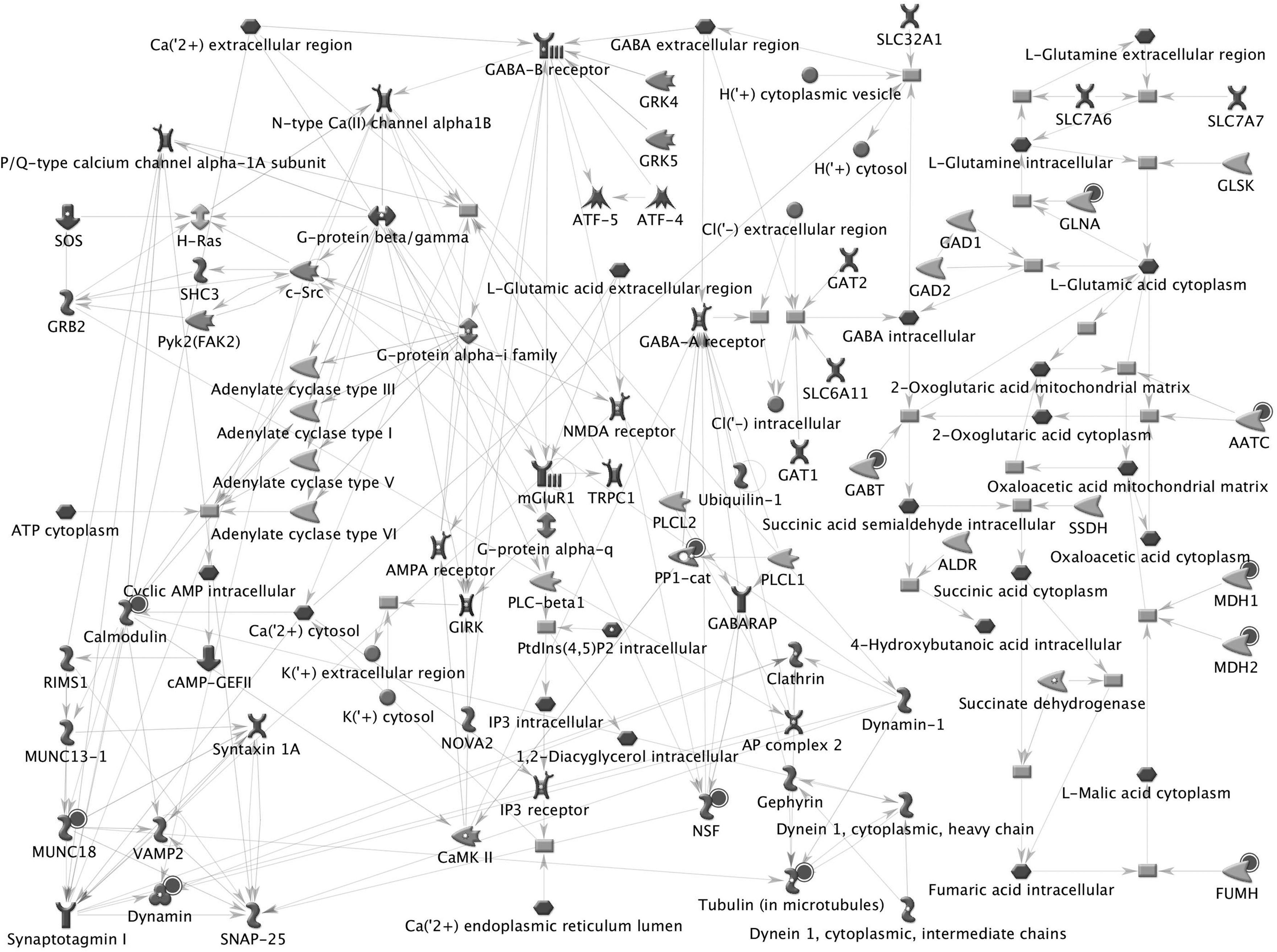
Neurophysiological process involving GABAergic neurotransmission, one of the major networks obtained from the olfactory bulb proteome. Twelve proteins of the olfactory bulb dataset were found mapped for the function. The proteins of the olfactory dataset were shown with black-colored spot.
Conclusion and Perspectives
This study reports the 2-DE proteome profile of normal adult D. rerio olfactory bulb tissue in its normal resting state and identifies 221 various proteins responsible for various functions and activities. This noncomprehensive study of the various proteins expressed in the olfactory bulb during basal conditions identified a wide range of proteins of pI, mass, localization, types, and functions. All the identified proteins from the olfactory bulbs are found to be concerned in metabolic and catabolic processes. Comparison of this study with our earlier study on D. rerio total brain proteome based on 1DE LC MS/MS and 2-DE MS/MS,13,14 found 90% proteome similarity between the datasets, whereas on comparison against the mature rat olfactory bulb proteome 15 identifies only 40% proteome similarity between the rat and D. rerio olfactory proteome dataset.
As D. rerio genome is similar to human by 70%–80%, this proteome profile of D. rerio olfactory bulb can direct better understanding of its function and role in various brain-derived activities. This study of understanding the different proteins expressed in the olfactory bulb during basal conditions would not only lead to understand various biomarkers underlying for various functions and diseases involving the olfactory bulb but also may be used as a marker array tool to understand the olfactory bulb involvement in various stress associated diseases and its development. The association of the D. rerio olfactory bulb proteome and schizophrenia disease might lead to a strong association of the disease and the olfactory tissue. Also understanding the tissue profile by both broad range and narrow range focusing will definitely lead to a better understanding of the proteome map of the tissue or organ. 22 This study complements the proteins identified both in the 3–10 complete pH range and also in three various narrow range IPG strips. This work also signifies the importance of understanding the proteome map of D. rerio towards establishing the animal as a potential model for translational studies.
Footnotes
Acknowledgments
The authors thank Dr. C. S. Sundaram for all the technical help involved in MALDI analysis. This work is supported by the Council of Scientific and Industrial Research (CSIR) and Department of Biotechnology (DBT).
Disclosure Statement
No competing financial interests exist.
References
Supplementary Material
Please find the following supplemental material available below.
For Open Access articles published under a Creative Commons License, all supplemental material carries the same license as the article it is associated with.
For non-Open Access articles published, all supplemental material carries a non-exclusive license, and permission requests for re-use of supplemental material or any part of supplemental material shall be sent directly to the copyright owner as specified in the copyright notice associated with the article.