
Abstract
Abstract
The present study was carried out to examine the use of zebrafish (Danio rerio) as a preliminary screening model for testing the effect of potential immunostimulant substances on the innate immune system. β-Glucan, a polysaccharide used widely as an immunostimulant, was used as a representative molecule and tested on zebrafish embryos and larvae. The efficacy of the molecule was evaluated by determining the differential expression of some selected genes related to the immune system by RT-qPCR. Larvae from 72 hours post fertilization were found at the optimal developmental stage for assessing the expression of the selected genes.
To verify if the β-glucan entered the larvae and therefore was responsible for the effects produced, the molecule was labeled fluorescently to check its localization by using microscopy. For estimating the effects of β-glucan on gene expression, zebrafish embryos and larvae were immersed in three different concentrations of β-glucan (50, 100, and 150 μg/mL) using five different exposure times. A stronger gene induction was observed when longer times of exposure and older larvae were used. The most evident effects of β-glucan were the overexpression of the genes TNFα, MPO, TRF, and LYZ. Moreover, slight changes in MPO expression were detected using a transgenic line of zebrafish (MPO::GFP), and a temporal increase in resistance against Vibrio anguillarum was found after β-glucan immersion. The assay used in this study permits the testing potential of immunostimulants in a simple and cost-effective way.
Introduction
The innate immune system is the ancestral immune mechanism; it is nonspecific and, therefore, not dependent upon previous recognition of any structures of the invader; moreover, it reacts in a very short time-scale. 6 The immune system of zebrafish is poorly developed during the earliest development stages of the organism, and larvae are consequently dependent upon the innate immune parameters for their defense against a wide variety of environmental microorganisms. 4
A wide range of substances can stimulate the immune system by promoting its functionality through the activation of pattern recognition receptors (PRRs), for example, toll-like receptors (TLRs). These receptors, located on the surface of innate immune cells, enhance the overall resistance to various diseases by binding to highly conserved molecules present on the surface of pathogens, known as Pathogen Associated Molecular Patterns (PAMPs). Some examples of PAMPs are the polysaccharides present in the cell wall of yeasts, algae, and fungi (such as β-glucan and chitin), peptidoglycans and lipopolysaccharides (LPS) present in the bacterial cell wall, and the viral or bacterial nucleic acids.2,7–10 The binding of a PAMP to its receptor transmits a signal that activates the response of the innate immune system.2,11 The stimulation of TLRs by microbial products leads to the activation of signalling pathways, resulting in the expression of proteins with antimicrobial activity (such as transferrin and lysozyme) and of inflammatory cytokines (i.e., interleukin 1β and tumor necrosis factor α) protecting fish against infections from pathogens. 10
Thus, PAMPs can be considered immunostimulants, that is, molecules capable of modulating the immune system, by increasing the host's resistance against diseases that in most circumstances are caused by pathogens. 8
The present study explores the use of zebrafish larvae, as a preliminary screening system, to evaluate the effects of potential immunostimulants. In order to test this system, β-glucan was used as a standard molecule. This compound is a well-known immunostimulant that has proven to be very effective on different fish species.8,10 The design of the method was accomplished by defining the optimal exposure protocol, through the selection of the larval developmental stage and a variety of genes affected specifically by stimulation of the innate immune system. The uptake of β-glucan was verified by labeling the compound with fluorescein and checking its presence in the larvae digestive system after 4 and 5 days of exposure. Then, the effects of the molecule were evaluated, by testing the expression level of the selected genes by RT-qPCR. In addition, a transgenic line, MPO::GFP, was used to visualize the inflammation response of zebrafish larvae after β-glucan immersion. Finally, the immunomodulatory effect of β-glucan on zebrafish was confirmed by detection of a protective effect of the compound against the presence of a fish pathogen (Vibrio anguillarum).12–15
Materials and Methods
Animals
Adult zebrafish (Danio rerio, Hamilton 1822) were maintained at 27°C in 60 L tanks, with aerated freshwater. Each tank contained an external filtration system (biological, chemical, and physical filtration), supplemented by an ultraviolet lamp. Zebrafish were maintained according to standard protocols. 16
Fish were fed with commercial flakes twice a day and were reared on a 12-h light/12-h dark cycle. Embryos were collected, rinsed, and then maintained in embryo water (EW) (CaCl2 66.2 mM, MgSO47H2O 13.4 mM, NaHCO3 21 mM, KCl 3.8 mM); they were transferred to Petri dishes, and then maintained at 27°C until the beginning of the experiments (Fig. 1).
Different stages of zebrafish development tested to evaluate the level of expression of some selected genes.
Labeling of β-glucan
β-Glucan was labeled with fluorescein, using fluorescein dichlorotriazine (DTAF, Sigma) following the protocol of McCann et al., with some modifications. 17 Ten mg of DTAF dissolved in 0.1 M borate buffer (pH 7.0) were added to 25 mg of β-glucan particles suspended in 0.1 M borate buffer. The mixture was allowed to react overnight, at room temperature, with gentle stirring. Uncoupled DTAF was removed by extensive washing with EW (approximately six times) and precipitated with ethanol, twice. β-Glucan particles were collected by centrifugation, dried, and dissolved in EW.
To visualize the presence of the polysaccharide inside the larvae, embryos of 4 hpf were immersed continuously to 100 and 150 μg/mL of the labeled β-glucan during 4 and 5 days. Fluorescence was then checked in the exposed larvae under a stereomicroscope (Leica MZFL III). Images were captured using a Leica DFC 360FX camera.
Exposure protocols
To identify the optimal zebrafish larval developmental stage for analyzing the immune genes expression, the expression level of a number of selected genes was evaluated during the first 144 hpf of untreated embryos. Embryos maintained in EW at 27°C were sampled at 4, 24, 48, 72, 96, 120, and 144 hpf (see Fig. 1), RNA was extracted, and gene expression evaluated.
Five different exposure protocols were tested; these are described and summarized in Figure 2 (protocols E1 to E5). In protocols E1, E2, and E3, embryos of 4 hpf were immersed continuously in β-glucan during 6, 5, and 3 days, respectively. In protocols E4 and E5, larvae of 72 hpf were exposed during 3 days and 1 hour, respectively. Zebrafish embryos and larvae were exposed directly in EW to 50, 100, and 150 μg/mL of β-glucan. All experiments were undertaken using triplicates of 30 embryos/larvae and they were repeated twice.

Summary of the different experiment exposure protocols used.
RNA preparation and cDNA synthesis
At the end of each exposure experiment, zebrafish embryos and larvae were frozen rapidly in liquid nitrogen. Total RNA was extracted using Trizol Reagent (Invitrogen) according to the manufacturer's instructions.
The quantity and quality of RNA samples were determined by capillary electrophoresis, using an Agilent 2100 Bioanalyzer (Agilent Technologies). The Bioanalyzer provides a RNA integrity number (RIN), with 0 corresponding to fully degraded RNA and 10 corresponding to intact RNA. 18 In all of the experiments, only RNA samples with a RIN of at least 8.5 were used. These values fulfill one of the requirements of a satisfactory qPCR experiment. 19 cDNAs were synthesized from the RNA samples in a reverse transcription reaction (RT) containing 40 ng of RNA per assay. RT was performed in a mix containing 1x TaqMan RT buffer, 5.5 mM MgCl2, 500 μM dNTPs, 2.5 μM oligo-dT, RNase inhibitor (0.4 U/μL) and 1.25 U/μL MultiScribe reverse transcriptase (Applied Biosystems). The mixture was incubated at 25°C for 10 min and at 48°C for 30 min, and the enzyme was inactivated at 95°C for 10 min.
Changes in mRNA expression of a collection of genes, related to the innate immune system, were monitored by real-time qPCR. The following genes were tested: interleukin 1β (IL1β); transferrin (TRF); myeloperoxidase (MPO); lysozyme (LYZ); toll like receptor 22 (TLR22); tumor necrosis factor α (TNFα); and heat shock protein 70 (HSP70). Primer sequences are listed in Table 1. 12
Quantitative PCR was carried out with an ABI PRISM 7000 sequence detection system (Applied Biosystems). Each reaction was performed in a 25 μL solution, containing 0.5 μL of the optimal concentration for each pair of primers (Table 1), 12.5 μL 2x SYBR Green PCR master mix (Roche), and 40 ng of cDNA template.
Reaction conditions were as follows: 50°C for 2 min and 95°C for 10 min, followed by 40 cycles of 95°C for 15 sec and 60°C for 1 min. A dissociation step was performed at the end of the PCR: 95°C for 15 sec, 60°C for 20 sec, and 95°C for 15 sec.
We used β-actin as a house keeping gene to normalize all the samples and each PCR reaction was performed in triplicate. The threshold cycles and copy number for each reaction were calculated using the 7000 system software; and expression levels of RNA were calculated using the 2-ΔΔCt method. 20 Results were expressed as mean±standard error. Data were analyzed using Student's t test and differences were considered statistically significant at p<0.05.
Exposure of the MPO transgenic line
Batches of 20 embryos of 4 hpf of the MPO::GFP line 21 were immersed in 100 μg/mL β-glucan. The control group was immersed in EW. Differences in gene expression were observed under a stereoscope (Leica MZFL III) after 5 days of exposure. The exposure was carried out in duplicate and repeated twice.
Bacterial culture conditions and infection procedure
The effect of Vibrio anguillarum serotype O1, strains NB10 GFP-labeled 15 was tested on zebrafish larvae. The bacteria were grown at 25°C in TSB (trypticase soy broth, Pronadisa: enzymatic digest of casein 1.7%, enzymatic digest of soybean meal 0.3%, sodium chloride 0.5%, dipotassium phosphate 2.5%, dextrose 2.5%) for 16 h. Ten μg/mL chloramphenicol and 0.5 mM IPTG (isopropyl-β-D-thiogalactopyranoside) were used as described by O'Toole et al. 15 Cells were spun down at 3500 rpm for 15 min and washed twice with sterilized EW with salinity 5% (EW5). Finally, bacteria were resuspended EW5.
Determination of CFU was performed by spreading 100 μL of resuspended bacteria on TSA (Tripticase soy agar, Pronadisa) with chloramphenicol.
Challenge test
Groups of 30 embryos at 4 hpf were immersed continuously in 100 μg/mL β-glucan for 5 days at 27°C. After this time, larvae were immersed in a final concentration of bacteria of 108 UFCs/mL, control groups were immersed in EW5. Mortality was analyzed at 2, 24, 48, and 72 hpi (hours post infection/immersion). This test was carried out in triplicate and repeated three times.
The presence of bacteria inside the larvae was checked after 2 hpi under a stereomicroscope (Leica MZFL III). Images were captured using a Leica DFC 360FX camera.
Results
Definition of the optimal developmental stage for testing effects on immune gene expression in zebrafish
We evaluated the expression of the immune genes selected during the normal embryonic development in untreated embryos. Figure 3 shows the expression level of each of the genes tested during the first 144 hours of embryos development. As shown in the figure, genes IL1β, LYZ, and HSP70 were expressed from the very early developmental stages (4 hpf); however, the expression of MPO, TNFα, TLR22, and TRF was not detected until 24 hpf.
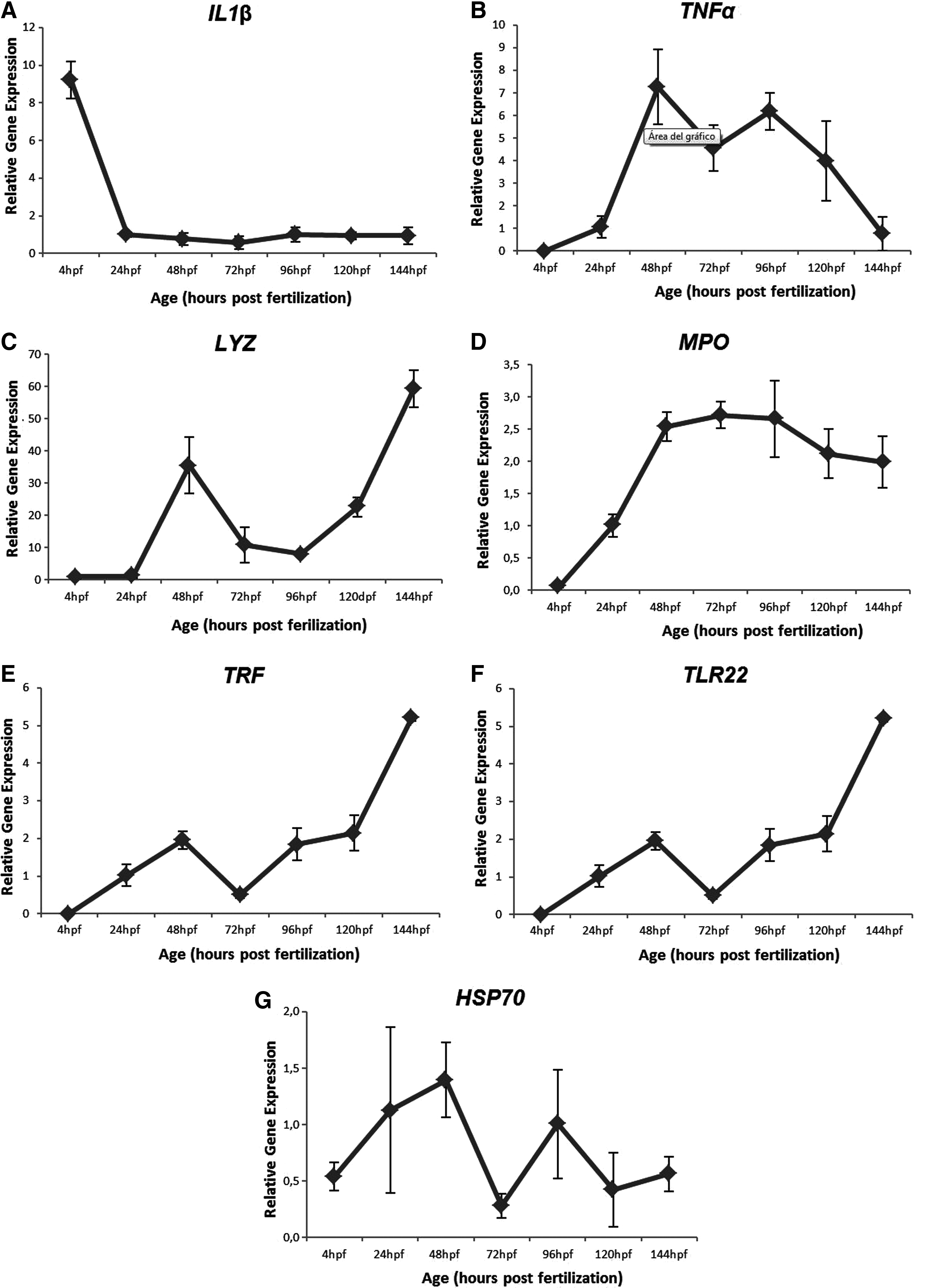
Expression profiles of the tested genes. Zebrafish β-actin gene was used as a control (not shown). Expression units were calculated by dividing the normalized expression values of each sample point by the normalized expression value at 24 hpf using the 2-ΔΔCt method. Each bar represents the mean of three replicates. Error bars represent standard deviation.
More specifically, IL1β showed a higher expression (9-fold) at the beginning of the development, whilst, after 24 hours, the level decreased and remained constant over time (Fig. 3A). Tumor necrosis factor α reached a maximum level of expression at 48 hpf and then it decreased gradually (Fig. 3B).
Lysozyme expression was detected from the earliest stages of development, but at very low concentrations; however, the expression of this gene increased at 48 hpf (35-fold). At 72 hpf, the expression decreased again until 120 hpf, when it started increasing again (Fig. 3C). Myeloperoxidase was first detected at 24 hpf, showing a constant expression level starting at 48 hpf (Fig. 3D). In the case of TRF, a biphasic expression profile was identified, whilst expression of this gene was detected from 24 hpf; however, its levels of transcripts decreased dramatically between 24–72 hpf. After 72 hours, the expression increased, reaching approximately a 5-fold increase at 144 hpf (Fig. 3E).
In this study, TLR22 was first detected at 24 hpf and had a tendency to increase until 144 hpf (Fig. 3F). The HSP70 expression profile was highly variable (Fig. 3G).
Evaluation of β-glucan uptake
β-Glucan was labeled with fluorescein and its presence was checked visually by microscopic analysis at the end of the experiment. These exposure experiments were carried out to verify that the polysaccharide entered the larvae and was indeed responsible for the effects detected.
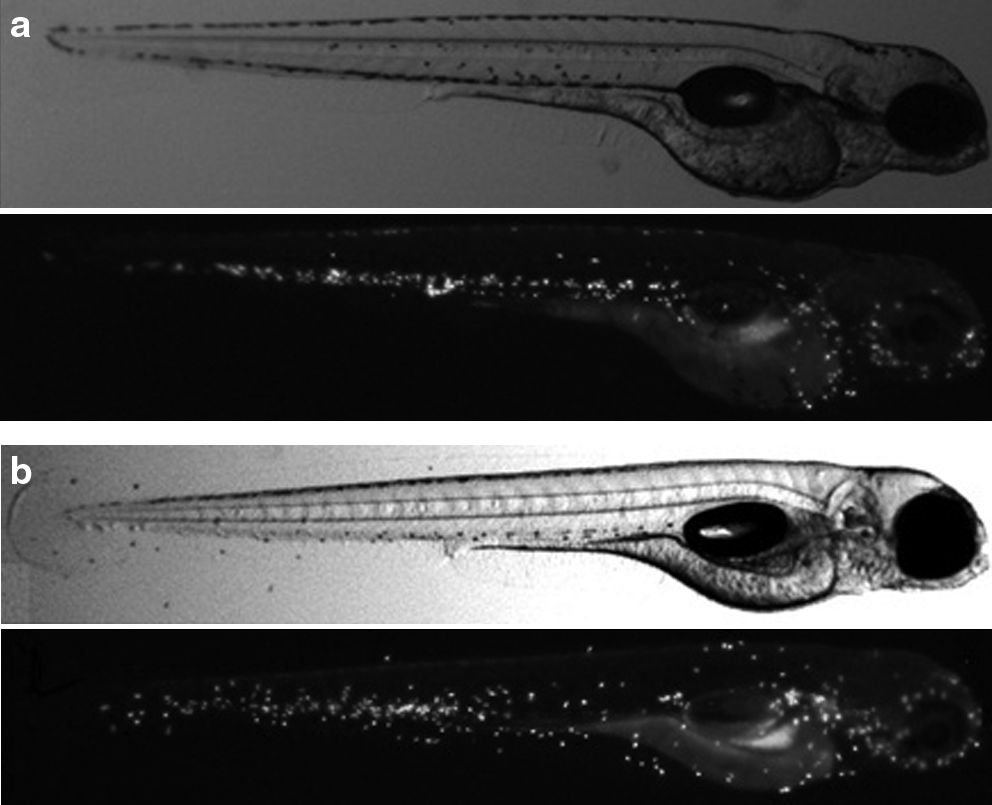
In all the exposure protocols tested, the presence of the molecule inside some larvae was evident. β-Glucan was observed throughout the digestive tract (Fig. 4). The presence of this molecule after 4 days of exposure was much lower (12.5%–30%) than after 5 days of exposure (83.2%–93.7%) (Table 2). These results showed an important increment of the entrance of the molecule from day 4 to day 5.

Localization of the polysaccharide inside the larvae. The β-glucan was labeled with DTAF, and it was detected in the digestive tract of the larvae.
Experiments were carried out with 20 larvae in duplicate.
Effect of β-glucan exposure on gene expression
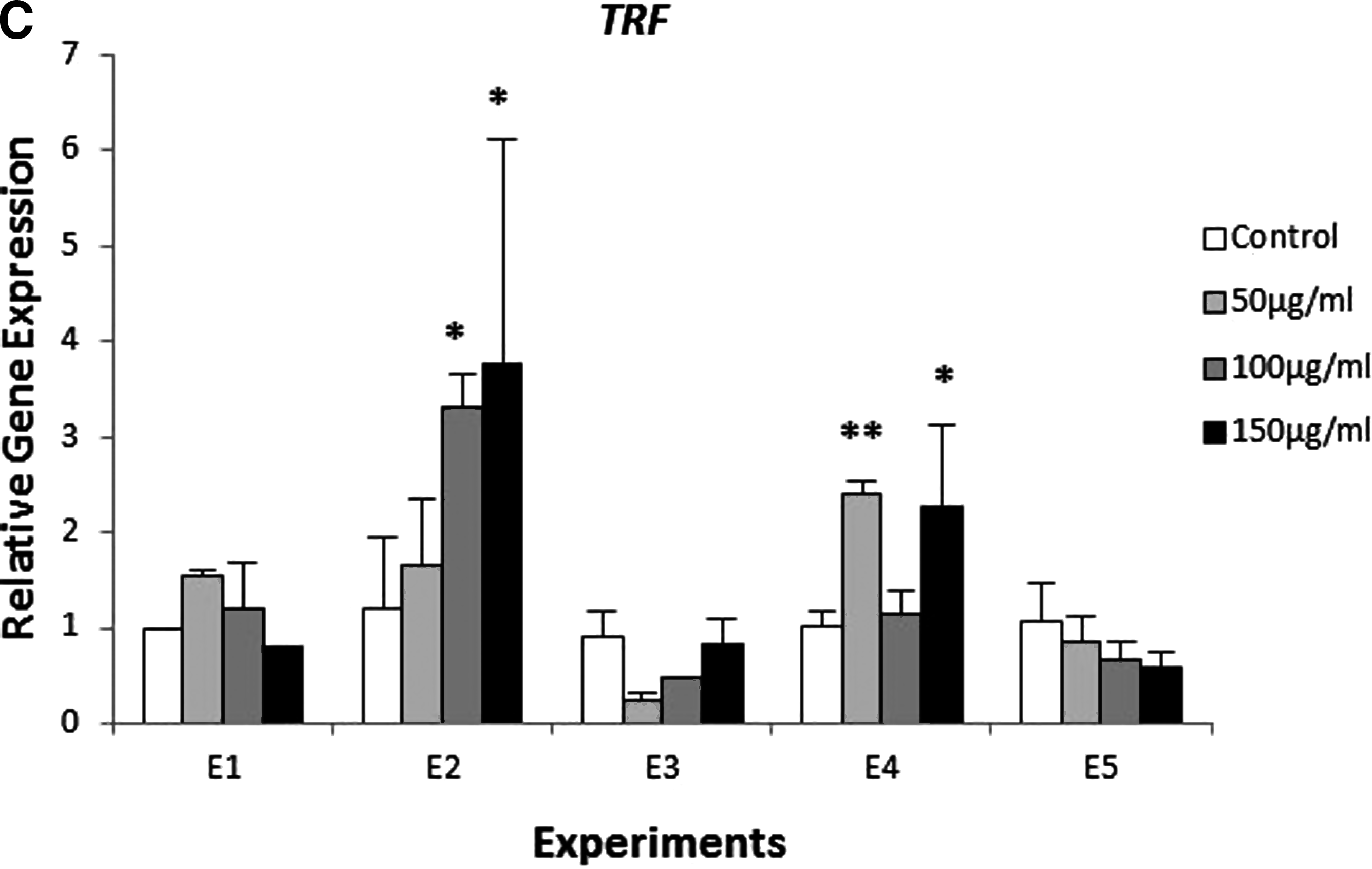
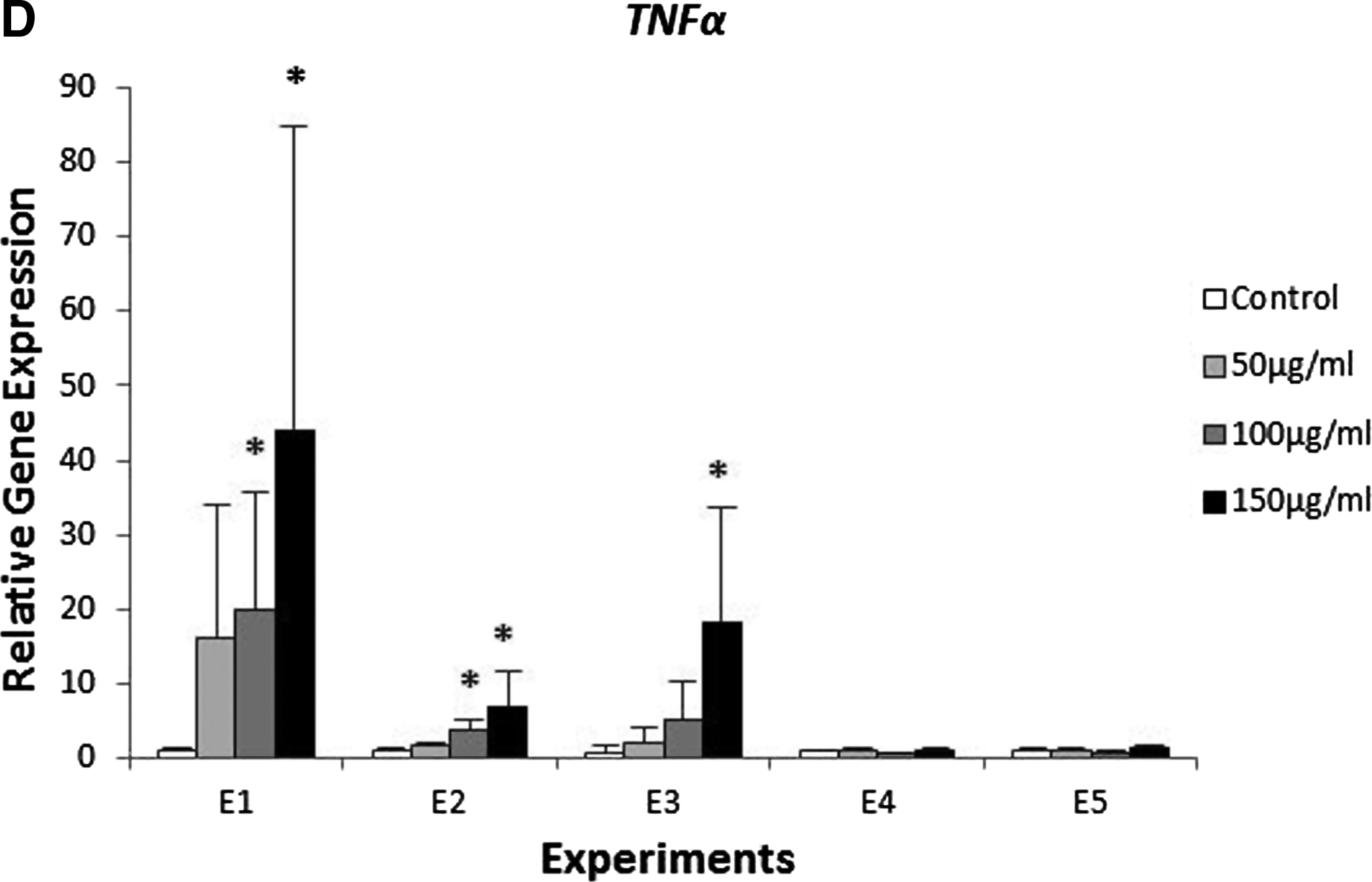
The exposure of zebrafish to β-glucan caused changes in gene expression. As a matter of fact, for longer times of exposure, higher changes in expression were detected. Specifically, in Experiments E1 and E2, we found a significant difference of the expression of MPO, LYZ, TRF and TNFα (Fig. 5).
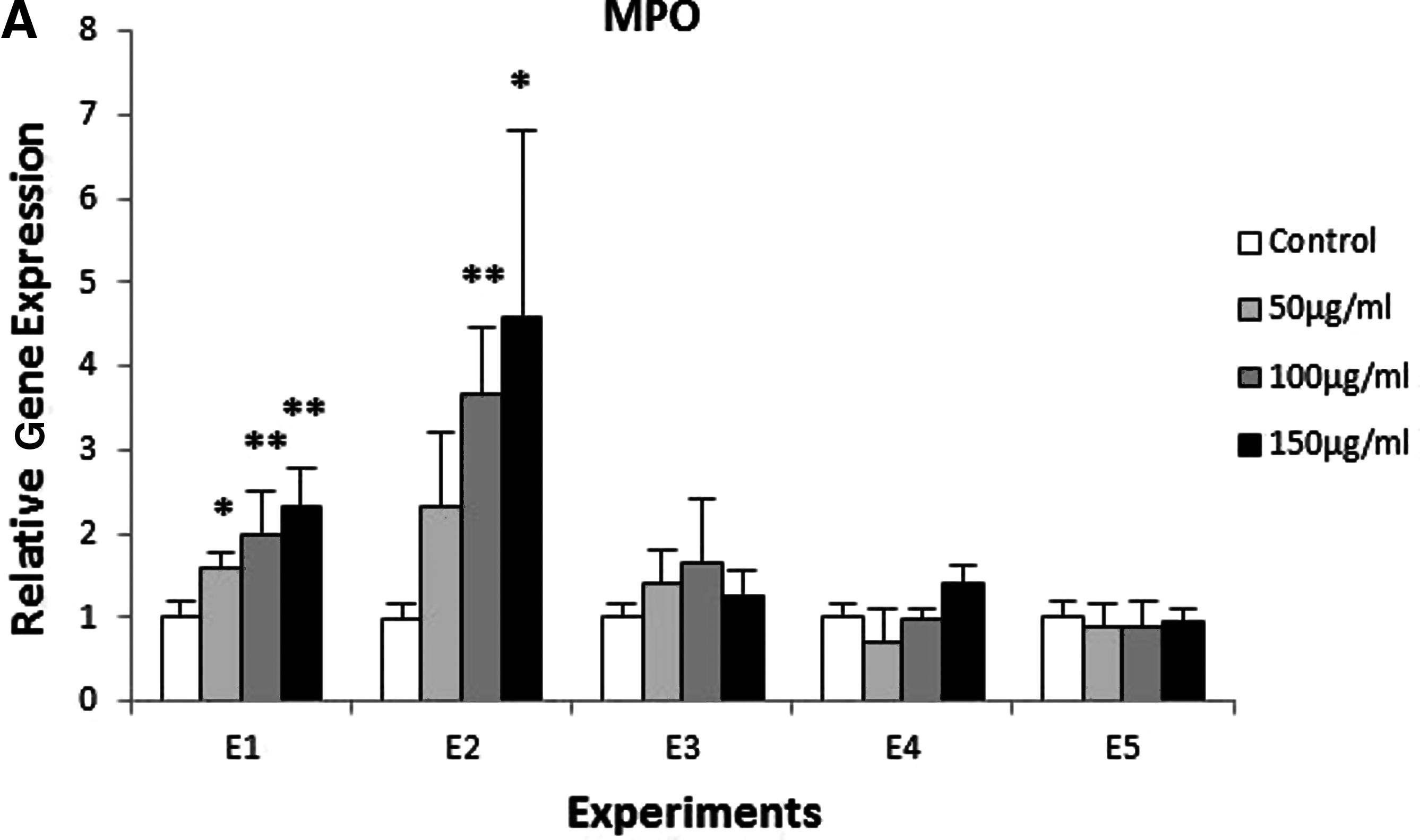
Expression of genes involved in the innate immune response in zebrafish exposed to different concentrations of β-glucan (50, 100, 150 μg/mL) and during different times of exposure E1, E2, E3, E4, and E5. Batches of 30 embryos. This test was carried out in triplicate and repeated twice. The expression was normalized with the housekeeping gene β-actin and compared with the controls (*p<0.05 and **p<0.01).

Myeloperoxidase showed a statistically significant increase at all the concentrations tested (higher than 1.5 fold), when exposure protocol E1 was used. Likewise, when exposure protocol E2 was used, it showed an overexpression of 3.6 (for exposure to 100 μg/mL of β-glucan) and of 4.5 fold (for exposure to 150 μg/mL of β-glucan). In the rest of the experiments performed, we did not find any change in the expression of this gene (Fig. 5A).
Lysozyme showed an overexpression in exposure protocols E1, E2, and E3. In E1 and E2, significant differences were observed at all the concentrations of β-glucan tested. However, in E3, an overexpression was found only at a concentration of 100 μg/mL. In E4 and E5, there were no variations in the level of LYZ expression between exposed samples and control groups (Fig. 5B).
Transferrin presented a significant increase in expression levels at 50 and 150 μg/mL of β-glucan in Experiment E4, greater than 2-fold (Fig. 5C). Likewise, TRF was also overexpressed in the highest concentrations tested in experiment E2.
Tumor necrosis factor α was overexpressed also at the highest concentration of β-glucan in exposure protocols E1, E2, and E3. In E1, we found high variability within replicates leading to high standard deviations. In experiments E4 and E5, no significant differences were found (Fig. 5D).
No changes were detected in expression of HSP70, TLR22, and IL1β, in any of the experiments performed (data not shown).
Exposure of the transgenic line (MPO::GFP) to β-glucan
The exposure of embryos originating from the transgenic line MPO::GFP caused a slight overexpression of MPO after β-glucan immersion. Figure 6 shows a comparison between the control group and the exposed larvae. It is clear from the photograph that the exposed larvae show a higher fluorescence with respect to the control and that the expression of MPO is widespread through the body of the larvae.
MPO expressing cells in MPO::GFP transgenic larvae of 120 hpf
Bacterial challenge
In a preliminary experiment the effect of salinity and survival rate of the bacteria in the EW5 was analyzed. No decrease in UFCs/mL was detected after 72 h of bacteria suspension in EW5.
The presence of the bacteria inside the larvae gut was observed. Bacteria were distributed in the first part of the digestive tract at 2 hpi and all through it after 24 hpi (Fig. 7).
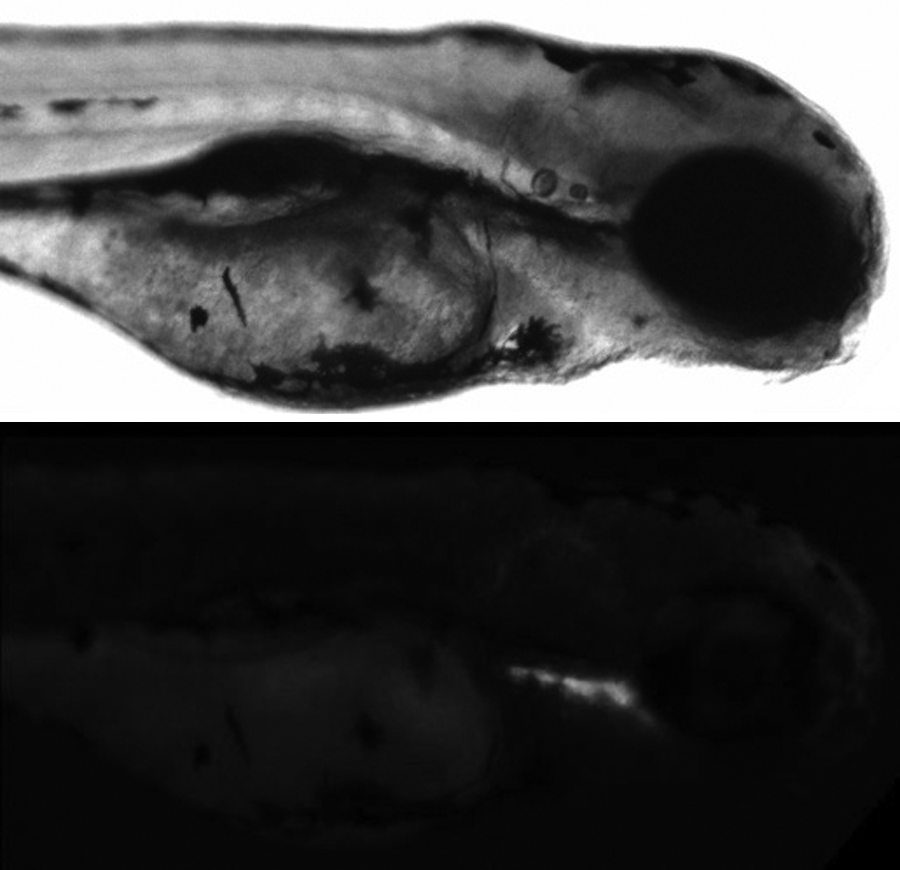
Zebrafish larvae infected with GFP-labeled V. anguillarum after 2 hpi. Note the presence of the bacteria in the first part of the digestive tract.
Three different challenge experiments were performed. The mortality rate of zebrafish larvae after addition of V. anguillarum is shown in Figure 8. In all experiments, the mortality rate of larvae previously exposed to β-glucan was significantly delayed compared with that of the control group, although the time of mortality varied within experiments.
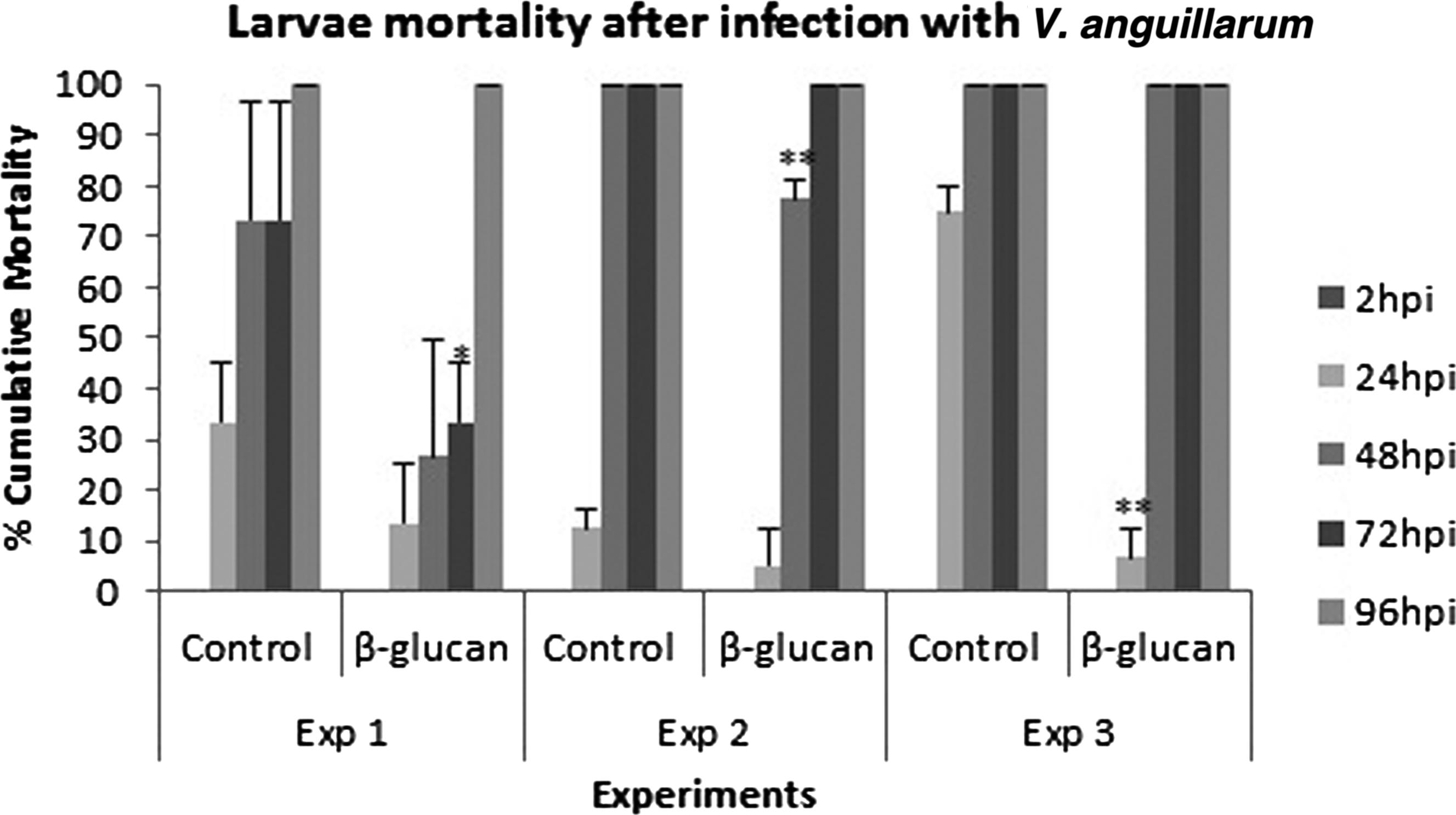
Average percent mortality in zebrafish larvae exposed to V. anguillarum. Treated zebrafish larvae were previously immersed in β-glucan for 5 days. This test was carried out in triplicate and repeated three times. (*p<0.05 and **p<0.01).
In Exp1, mortality rates were significantly higher in the control group at 48 and 72 hpi as compared to the test group. In Exp2, differences were observed at 48 hpi, and in Exp3 at 72 hpi. Mortality rates reached 100% in both groups of all experiments at 96 hpi.
Discussion
In nature, fish eggs are released and fertilized externally. As such, the resulting embryos and larvae are exposed to an aquatic environment full of potential pathogens, at a time when their immunological capacity is still critically limited. Although the embryos are protected by the chorion, as well as by several innate and adaptive immune substances that are transferred to the embryos during fish vitellogenesis, the most effective way of dealing with hostile situations is to discriminate rapidly self from non-self by recognizing PAMPs.10,22 The recognition of PAMPs by the PRRs induces the immune response. In other words, the invasion of a microorganism causes an immediate response of the fish. 2
In this study, the effect of an immune-modulating substance was evaluated by testing the differential expression of some selected genes in zebrafish larvae. The selection of such genes was made considering different routes of the innate immune system and to have biomarkers able to detect any change in gene expression after the exposure to a molecule that acts as a PAMP. 23 The following genes were used: a toll-like receptor TLR22, which is a protein that acts as a receptor and is stimulated by PAMPs; 24 two cytokines, TNFα and IL1β with proinflammatory and antibacterial activity;25–27 a growth inhibitor factor, TRF, which blocks or delays bacterial growth; two enzymes: one with bactericidal activity, LYZ,28,29 and one that contributes to the general inflammatory response, MPO; 30 and, finally, a heat-shock protein, HSP70, which plays essential roles during development by protecting against damage from stressors.10,31,32
Before testing the effects of the exposure to an immunostimulant, the expression level of some selected immune genes was analyzed during the early developmental stages of the zebrafish (4–144 hpf). This assay permitted gene expression analysis to be undertaken, at the optimum time on further experiments that were aimed at testing the effects of immunostimulants.
The expression profiles of the selected genes provided results similar to those reported by other studies. Ito et al. 27 found also high levels of IL1β at the first hours of the embryonic development, with decreasing levels towards the later stages. The early significant high level of IL1β suggests that the detected transcripts are not produced by the embryo itself but most probably are of maternal origin. The transfer of non-immune factors, such as complement and lysozymes, from the mother to the offspring has been previously reported in fish. These transferred maternal molecules seem to protect fish at early developmental stages from pathogens.22,33,34 Pressley et al. 35 detected also the expression of TNFα at 24 hpf in accordance with our findings.
Our results in terms of MPO are consistent with those reported previously by Bennett et al., (2001) who detected MPO expression at 18 hpf by in-situ hybridation. 36 Mudumana et al., (2004) studied the expression profile of TRF in early embryogenesis and larval stages, and they also observed a biphasic expression pattern. 37 In the case of LYZ, Liu and Wen (2002) described a similar expression pattern as that reported in our study. 28
According to the results obtained and from those reported in the literature, we can conclude that at the early stages of development (4–72 hpf) there is a higher variability in gene expression, in comparison to the following days. Thus, it could be assumed that the best moment to evaluate gene expression, for assessing the effect of immunomodulatory molecules such as β-glucan, would be starting from 72 hpf, independently of the exposure time. In fact, at this developmental stage, all the selected genes were expressed (Fig. 3). Additionally, since zebrafish embryos hatch between 48 and 72 hpf and the mouth starts gaping at 72 hpf, 38 larvae of this age are free from the chorion and in direct contact with the molecule to be tested; thus, facilitating its entry into the body by diffusion and/or through the mouth.
To verify the presence of the polysaccharide inside the gut of the larvae, we labeled the β-glucan with a fluorescent molecule and localized the fluorescence in the larvae, by microscope analysis. We observed an important increase in the number of larvae containing β-glucan inside the gut from day 4 (12.5%–30%) to day 5 (83.2%–93.7%) of exposure (Table 2, Fig. 4). These results reinforce the results of gene expression, that is, the longer the exposure (E1 and E2) the higher the effect of the polysaccharide. In both experiments, the exposure to the polysaccharide began at 4 hpf, and was terminated between 120 and 144 hpf. During the first 48–72 hpf, the embryo is surrounded by the chorion. β-Glucan may penetrate through the pore canals during the first 48–72 hpf because their diameter is sufficiently wide for the entrance of the molecule. 39 However, this penetration is random and might not be sufficient alone to produce a strong effect in the gene expression, as was observed in exposure protocol E3. In contrast, after hatching, the molecule enters into direct contact with the embryo. However, this entry alone does not appear to be sufficient to cause a strong effect on gene expression (E4). According to the results, only the exposure protocols that included embryos surrounded by the chorion, as well as larvae after hatching (E1 and E2), seem to produce a strong effect in gene expression. This cumulative effect could be due to the length of the exposure.
The present study demonstrates that the exposure of larvae to β-glucan, by simple immersion, can raise significantly the innate immunity levels. However, it indicates also that exposure time and larvae age are important factors to consider when designing an experiment for assessing immunostimulant activity. According to the results described above, we were able to detect an effect after immersion of zebrafish larvae in a suspension of β-glucan, through the increase in expression of MPO, LYZ, TRF, and TNFα. The influence of the polysaccharide was higher in longer exposures and older larvae. The highest level of overexpression was found in experiments E1 and E2, after an exposure to the polysaccharide of 5 and 6 days. This observation is in agreement with the finding of a higher level of β-glucan in older larvae.
Using embryos originating from a transgenic line for MPO, 21 we could detect a slight increase in expression after β-glucan immersion. Although the expression of MPO may vary depending on the health status of each larva, the fluorescence of the control group was lower in comparison to the treated group. These results corroborate those from gene expression analysis, were an overexpression of MPO was observed.
Moreover, we observed that the exposure to β-glucan is responsible for an increase of the resistance against infection. Immersion of zebrafish larvae with the molecule, in fact, delayed the mortality of larvae after addition of V. anguillarum. In spite of some variability within experiments, probably due to variability in hatchings, bacterial concentration in the inoculum, and stochastic colonization of bacteria in the gut of the larvae, in all cases immersion of β-glucan produced a temporal resistance of the larvae to the bacteria, delaying their death for a few hours.
These results indicate that the changes observed in the expression of immune genes are related to the increase of the resistance to a bacterial infection.
According to the literature, the time of exposure to an immunostimulant molecule necessary to trigger nonspecific defensive mechanisms in fish, may be highly variable.7,23 Watzke et al., (2007) 40 exposed dechorionated zebrafish embryos of 28 hpf during 4 hours at different concentrations of Escherichia coli LPS. These authors observed that just one of the concentrations tested (1 ng/μL) had an effect on the expression of TNFα and IL1β. Novoa et al., (2009) tested different immunostimulants at the early stages of zebrafish embryonic development; they observed an overexpression of IL1β in the case of LPS from E. coli. 41 Pressley et al. (2005) observed an overexpression of TNFα and IL1β after infection of embryos with Edwarsella tarda. 35 Other studies with adult zebrafish have shown an overexpression of genes involved in the innate immune response after a bacterial infection, as was the case of TLR22 after Mycobacterium marinum infection; 42 TNFα and IL1β in the case of Aeromonas hydrophila immersion; 43 and TNFα, MPO, and TRF after V. anguillarum infection. 12 Considering all these results, it can be seen that the major effect of immunostimulants is on cytokines and the genes related to the innate immune system.
The immunostimulant effect of β-glucans has been demonstrated also in other fish species. For example, a number of studies have shown that the oral administration of β-glucans to fish also causes an increase in the resistance to bacterial infections, as well as in the concentration of nonspecific immune factors such as serum lysozyme.44–48
It can be concluded also that the developmental stage and time of exposure are important factors when testing the effect of an immunostimulant. This study has shown that the most appropriate time of exposure is 5 days, commencing with recently fertilized embryo. A simple contact with the molecule by immersing the embryos/larvae in a suspension of the substance to test is sufficient for determining its effects on gene expression. Moreover, the contact with β-glucan during this time confers to larvae a temporal resistance against an infection.
Thus, zebrafish can be considered as a model system to perform the fast screening of immunomodulating molecules. The proposed assay is short and has the advantage of allowing the testing of different potential immunostimulants in a very simple way; this reduces time, cost, and the number of animals used.
The major components of the immune recognition systems and defense signaling pathways are highly conserved between fish, but significant divergence has been also observed among the innate immune recognition and components that are involved in direct interactions with pathogens.5,42,49 Further work will focus upon increasing the number of genes involved in other mechanisms of the immune system.
Footnotes
Acknowledgments
We thank sincerely: S. Renshaw (University of Sheffield) and R. ÓToole et al. (University of Umeå) for their generosity for providing the zebrafish transgenic line MPO::GFP and the GFP labeled V. anguillarum, respectively; J.C. Arboleya (AZTI-Tecnalia) and Professor Michael Collins (SOES Southampton, UK and AZTI-Tecnalia) for critical reading of the manuscript. This study was supported financially by the Science and Innovation Ministry of Spain (Project INMUGAL PID-580000-2009-1), the Agriculture and Fisheries Department of the Basque Government (Project IA2009Daretest). UO is the recipient of a PhD Fellowship from the Iñaki Goenaga Foundation.
Disclosure Statement
No competing financial interests exist.