
Abstract
Abstract
Zebrafish is a good model for studying regeneration because of the rapidity with which it occurs. Better understanding of this process may lead in the future to improvement of the regenerating capacity of humans. Signaling factors are the second largest category of genes, regulated during regeneration after the regulators of wound healing. Major developmental signaling pathways play a role in this multistep process, such as Bmp, Fgf, Notch, retinoic acid, Shh, and Wnt. In the present study, we focus on TGF-β-induced genes, bigh3 and bambia. Bigh3 encodes keratoepithelin, a protein first identified as an extracellular matrix protein reported to play a role in cell adhesion, as well as in cornea formation and osteogenesis. The expression of bigh3 in zebrafish fins has previously been reported. Here we demonstrate that tgf-b1 and tgf-b3 mRNA reacted with delay, first showing no regulation at 3 dpa, followed by upregulation at 4 and 5 dpa. Tgf-b1, tgf-2, and tgf-brII mRNA were back to normal levels at 10 dpa. Only tgf-b3 mRNA was still upregulated at that time. Bigh3 mRNA followed the upregulation of tgf-b1, while bambia mRNA behaved similarly to tgf-b2 mRNA. We show that upregulation of bigh3 and bambia mRNA correlated with the process of fin regeneration and regulation of TGF-b signaling, suggesting a new role for these proteins.
Introduction
The fin is comprised of both an endo- and an exoskeleton encompassing many segmented fin rays. Only fin rays have the ability to regenerate. The caudal fin contains between 16–18 fin rays. The process of regeneration can be divided into three phases: wound healing, blastema formation, and regenerative outgrowth. 5 Immediately after amputation the epidermis is induced to migrate to the wounded site, covering it within 1–3 hours to form a thick epidermal tissue called wound epidermis. The formation of wound epidermis does not require cell proliferation, 6 but this step is required for initiation of regeneration and it plays an important role as a source of growth factors. During this period, genes belonging to the Wnt-signaling pathway play a major role. In a second phase, the basal layer of wound epidermis expresses bmp2b, patched, and shh. Blastema formation occurs between 12 and 48 hpa and results in a proliferative mass of mesenchymal cells that sits atop each severed fin ray, which ultimately gives rise to the new structures of the fin. The mature wound epidermis is thought to be a source of growth factors that stimulate formation of the blastema and maintain its function. The blastema cells act like stem cells because their descendants either continue self-renewal or leave the niche and differentiate into new regenerate tissues. β-Catenin signaling plays an important role as β-catenin is presumed to function in maintaining cell–cell interactions that facilitate migration during this step. 7 Finally, between 2 and 14 dpa, the blastema cells proliferate and contribute to the regenerative outgrowth. The cell cycle in the blastema during regenerative outgrowth is 6 times faster than during the initial phase of blastema formation. The architecture of the regenerating fin results from the differentiation of cells of the blastema, forming nerves, bone, blood vessels, connective tissue, pigment cells, and skin. When regeneration is achieved, the blastema and wound epithelium are incorporated into the differentiated tissues.
Regeneration is the reactivation of the developmental processes in postembryonic life to restore missing tissues. Mesoderm is responsible for induction of a broad range of mesodermal markers and tissues. 8 Transforming growth factor β (TGFβ) superfamily signals have emerged as central components of the inductive process. 9 Members of this superfamily were shown to induce differentiation of cartilage and bone-forming cells in mammals and lepidotrichia-forming cells in zebrafish. 10 Bigh3 is induced by Tgf-β and is known to play a role in developmental processes, as well as in wound healing. In mammals, BIGH3 encodes keratoepithelin (KE), a 68-kDa protein expressed in the extracellular matrix (ECM) of many tissues, including nerve, bone, and skin.11,12 Despite many studies, little is known about its function. Four Fas1 homology domains, one EMI domain, and an RGD sequence characterize this protein, and studies have identified several integrin binding sites, including αvβ3 and αvβ5, within these domains.13–16 It is therefore assumed that KE may regulate cell–cell or/and cell–ECM interaction by binding ECM proteins such as fibronectin, laminin, and different types of collagen.17–20 Its role in wound healing has been highlighted during angiogenesis and vascular remodeling where it plays an important role in endothelial cell–matrix interaction. 21 During cartilage differentiation and osteogenesis, the protein functions as a negative regulator of mineralization.22,23
In human, BIGH3 has mainly been associated to corneal dystrophies, a group of inherited diseases characterized by amyloid or nonamyloid deposits in various layers of the cornea, and to various forms of cancers.12,24 These studies showed a strong genotype–phenotype correlation, especially for mutation of R124 and R555, the two most frequently mutated codons. Interestingly, despite their almost ubiquitous expression, in the event of mutations, pathological KE deposits are only seen in the cornea. 25 However, as these diseases originate from gain-of-function mutations, they do not help in understanding the physiological role of BIGH3 and through which pathways BIGH3 could act on regeneration and wound healing.
During fin regeneration, the level of apoptosis induced by fin amputation should be kept low in order for the proliferation and differentiation pathways to be effective.
In this study we focused on the role played by bigh3 and bambia in fin regeneration and its correlation with wound healing, osteogenesis, and development. We demonstrated that these genes were upregulated during fin regeneration.
Materials and Methods
Zebrafish husbandry and fin amputations
ABTL wild-type zebrafish were maintained as previously described.7,26 To determine the ratio of regeneration in the zebrafish fin, we first amputated the tail fin in 30 adults (males and females, 10–18 months of age). RNA was prepared from those fins and used for cDNA synthesis and for the determination of the baseline expression level of the genes studied. The fish were than divided into three groups and left to recover for 3, 4, 5, or 10 days post amputation (dpa). At these time points, the growing tissue of the fin was measured. For caudal fin amputations, fish were anesthetized in 0.1% tricaine, and fins were cut using razor blades. Six animals were used at each time point, and the growth was measured from the cut site to the end of new growing tissue. Directly after amputation, fish were shifted to 28.5°C water for the remainder of the experiment. Uncut control fish were kept at 28.5°C in the same system. The veterinary office of the state of Valais approved these experiments.
RNA extraction and cDNA synthesis
RNA from tail biopsies was obtained using the TRIZOL reagent following the manufacturer's instructions (Invitrogen, Carlsbad, CA). RNA concentration and purity were measured using NanoDrop ND-1000 (NanoDrop Technologies, Inc). Equal amounts of RNA were then transcribed into cDNA using Stratascript Reverse Transcriptase according to the manufacturer's instructions (Stratagene, La Jolla, CA).
Quantitative RT-PCR
PCR-specific primers were designed spanning two consecutive exons and in the 3′UTR using Clone Manager Professional Suite software (Sci Ed Central, Cary, NC). Amplifications were performed on SYBR Green mix containing 15 ng cDNA and 1 μM of each primer. Amplifications were performed with the MX3000P apparatus (Stratagene) using the following protocol. After a first step of 10 min at 95°C, samples were submitted to 40 cycles of amplification consisting of 30 sec incubation at 95°C, followed by 1 min incubation at 58°C. Data were analyzed using the MX3000P software; ef1α and β-actin were used as reference. The amount of mRNA in each sample was expressed in arbitrary units after normalization with ef1α and β-actin of the same condition. Primers for qPCR are listed in Table 1. Student's t-test was use for comparing the means of the different samples. For each time point, 6 fins were collected individually and analyzed in triplicate.
In situ hybridization
Standard methods for one-color whole-mount in situ hybridization were used. Samples were fixed overnight in 4% formaldehyde/phosphate buffered saline (PBS). The fins were then washed in PBS, dehydrated in methanol (MeOH) at room temperature and stored at −20°C. The tails were rehydrated in MeOH/PBST, then washed in PBS, and refixed in 4% formaldehyde/PBS at room temperature for 20 min. After the PBS wash, proteinase K (10 μg/μL in PBST) digestion was performed for 10 min and rapidly stopped with 3 changes of PBST. The samples were fixed in 4% formaldehyde for 20 min in hybridization buffer (500 μL) (50% deionized formamide/5x SSC/0.1% tween 20/5 mL/mL torula RNA) and incubated at 68°C for 2 h before replacement with 100 μL of probe diluted with hybridization buffer. Hybridization was performed at 68°C for 14 h. In situ probe were generated from pGEM-Easy. Specific primers are listed in Table 1.
Whole-mount in situ program
Samples were stained in 24-well plate with 500 μL SB buffer containing 3.5 μL/mL NBT and BCIP (Roche) and when the signal appeared, the reaction was stopped with 500 μL of stop solution (2 mg/mL glycine pH 2.5) for 15 min. The samples were subsequently washed in PBST and fixed again in 4% formaldehyde/PBS overnight at 4°C. Finally, they were washed with PBS solution and stored at 4°C in glycerol until further use.
Results
Zebrafish fin regeneration
At 3 dpa, a tiny amount of white tissue containing small vessels with circulating blood cells was observed at the tip of the amputated fin (Fig. 1B). At 4 dpa, this tissue was extended, and growing fin rays were observed under microscope (Fig. 1C). At 5 dpa, the pigmentation was already present (Fig. 1D). Over the following days, the tissue continued to grow and the fin was completely regenerated at about 2 weeks post amputation (Fig. 1E–G). Figure 2 shows the quantification of fin growth during the first 10 days.

Dynamics of zebrafish fin regeneration from 0 to 15 days post amputation (dpa).

bigh3 normalized mRNA expression (NGE) by qPCR with ef1α and β-actin as reference genes during zebrafish fin regeneration, together with fin growth on the right. The expression levels were normalized with the expression at 0 dpa.
Bigh3 mRNA expression in regenerating zebrafish fin
Various signaling pathways were proposed to play a role in zebrafish fin regeneration. We investigated a role of TGF in bigh3 activation in zebrafish fin regeneration. As previously described, tissue samples were collected at five different time points (0, 3, 4, 5, and 10 dpa) and the expression of bigh3 was analyzed by quantitative PCR (Fig. 2). At 3 dpa, a small increase in bigh3 expression was observed. At that stage, this increase was not statistically significant (p=0.059). However, at 5 dpa, the expression reached a 4.6x increase as compared to day 0 (p=0.002), which suggests a continuous demand of bigh3 during the process of regeneration. At 10 dpa, the expression of bigh3 declined towards levels comparable to those seen at 3 dpa and baseline (p=0.056). Similarly to qPCR, in situ hybridization showed increased bigh3 expression at the tip of the blastema. No expression of bigh3 was seen at 0 dpa; at 3 dpa, bigh3 was expressed in the whole blastema, later becoming concentrated at the periphery at 5 dpa. At 10 days, bigh3 expression was still present (Fig. 3).
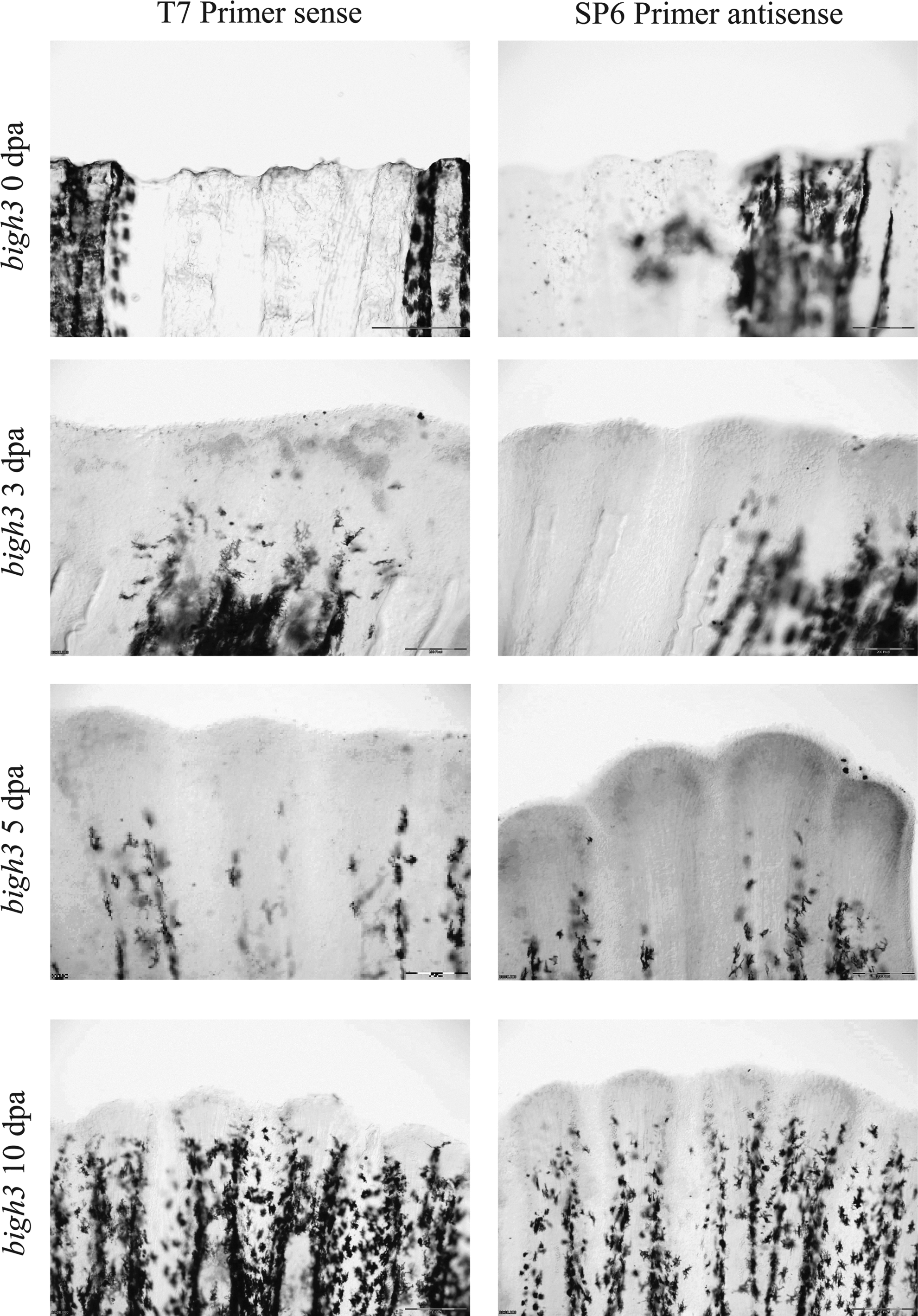
In situ hybridization with bigh3 during fin regeneration at 0, 3, 5, and 10 dpa. Comparison between sense and antisense probes. Scale bar represents 200 μm.
Tgf-beta signaling during regeneration
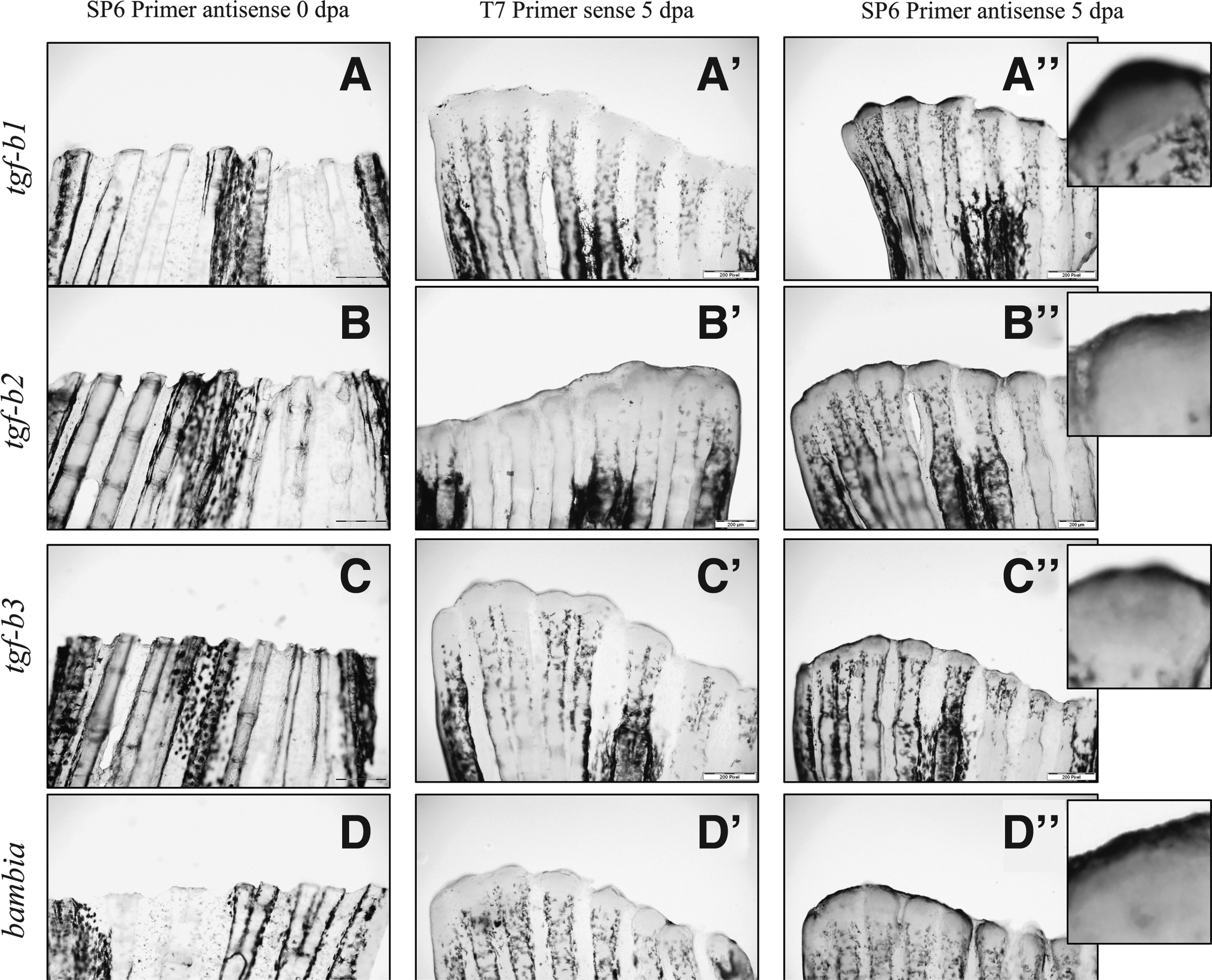
We analyzed expression of tgf-β1, -β2, and -β3 in regenerating fin according to time as described above. Tgf-β2 showed a 2.18-fold upregulation at 3 dpa (p=0.019) and remained significantly high until 5 dpa (2.79×, p=0.008). Tgf-b1 and tgf-b3 reacted with a delay and were upregulated at 4 dpa (2.11×, p=0.02, and 1.86×, p=0.034, respectively) reaching a maximum value at 5 dpa (2.97×, p=0.007, and 2.20×, p=0.018, respectively). At 10 dpa, tgf-b1 and tgf–b2 were back to normal values while tgf-b3 remained upregulated (1.8×, p=0.043) (Fig. 4). Similar to qPCR, in situ hybridization showed increased of tgf-β1, -β2, and -β3 expression at the tip of the regenerating fins at 5 dpa, together with expression in the blastema (Fig. 5).

tgf-β1, -β2, and -β3 normalized mRNA expression (NGE) by qPCR with ef1α and β-actin as reference genes during zebrafish fin regeneration. The expression levels were normalized with the expression at 0 dpa.
Bambia normalized mRNA expression by qPCR with ef1α and β-actin as reference genes during zebrafish fin regeneration. The expression levels were normalized with the expression at 0 dpa.
Other members of TGF-β signaling are also activated during regeneration
It was previously described that bambia, a TGF-pathway regulator and tgf-βrII were also modulated during the process of regeneration. 27 Bambia and tgf-βrII were upregulated at 3 dpa 4.2x (p=0.002) and 1.63x (p=0.044), respectively. At 5 dpa, they reached a maximum value of 5.82x (p=0.001) and 2.55x (p=0.011), respectively (Fig. 6). The expression of bambia and tgf-βrII were back to levels comparable to nonamputated fins at 10 dpa (Figs. 5 and 7).

In situ hybridization with tgf-β1, -β2, -β3, and bambia during fin regeneration at 0 and 5 dpa. Comparison between sense and antisense probes. Expression was present in the tips of the fins surrounding the blastema and in the blastema. Scale bar represents 200 μm.

tgf-βRII normalized mRNA expression (NGE) by qPCR with ef1α and β-actin as reference genes during zebrafish fin regeneration. The expression levels were normalized with the expression at 0 dpa.
Discussion
Regeneration is the restoration process of lost parts of the body. The term includes wound healing, tissue repair, and various kinds of restoring activity. The models studied involve epimorphosis, in which lost part is reproduced actively by cell proliferation and dedifferentiation. Regeneration is thought to reflect the mechanisms of development. The newt or axolotl can replace a lost limb within 1 to 2 months.28,29 Humans, however, can replace only the tips of their digits.30–32 Teleost fish can regenerate such complicated structures as fin5,7,33 or heart. 34 Their process of regeneration involves wound healing and regenerative outgrowth in which fin rays have to grow as well as epidermal/skin tissue, in addition to new vessel formation. Zebrafish regeneration has now been studied for more than 10 years. Factors implicated in early development such as Wnts, FGF, BMPs, and retinoic acid have been shown to play a role in the regeneration process. In this study, we focused on TGF-β signaling, which has been intensively studied in human wound healing process, and specifically its target gene, Bigh3 and bambia.
Bigh3, a target of TGF-β, could play a role in building up lepidotrichia, considering that it has already been associated with osteo- and chondrogenesis.22,23,35 BIGH3 has also been isolated from collagen-rich fractions of pig cartilage and from rabbit cornea. 36 In these studies, BIGH3 was demonstrated to play a role in wound healing in the rabbit cornea, especially during vessel remodeling. In angiogenesis, it probably increases the endothelial cell–extracellular matrix interactions. 21 BIGH3 also plays a role in skin regeneration (together with TGF-β3 and TGF-βRII). TGF-β1 is the most studied member of this family for its role in the wound healing process.37–41 It induces fibrosis and collagen synthesis in wound healing in addition to the inflammatory reaction, but only transiently participates in the repair and regeneration of tissue.42,43 In our study we did not observe any significant regulation of tgf-β1 expression at 3 dpa. At 5 dpa, however, its level was increased. Previous studies situated the process of wound healing within 12–24 hpa in zebrafish fin. Our study was performed after that period, which could explain the differences in the results. Tgf-β1 is also upregulated in the preparation phase of regeneration in salamander, whereas it is downregulated later on during the regeneration phase, which corresponds to the regenerative outgrowth phase in zebrafish. 44 Thus, the timing of activation of tgf-β1 seems to be conserved between these two species. Embryonic wounds that heal without a scar have low levels of tgf-β1 and tgf-β2 and high levels of tgf-β3. 43 The analysis of the expression of tgf-β1 and tgf-β2 in our model sustained that, apparently not only in rats, mice, and pigs, but also in regenerating zebrafish fin, these two growth factors need to be quickly returned to low levels in order to pursue the normal wound healing process. The upregulation of tgf-β3 is observed in embryonic wound healing. This skin morphogenic factor is predominantly synthesized by keratinocytes and fibroblasts. TGF-β3 and TGF-βIIR are involved in so-called “traffic control” occurring during wound healing. In unwounded skin, cells are fed by plasma; after the appearance of a wound, cells encounter transition of serum back to plasma. Human serum promotes epidermal cell migration but not dermal cell migration. In contrast, human plasma promotes dermal cell migration and not epidermal cell migration. This switch is regulated by TGF-β3, undetectable in serum but present in high amount in plasma, and by TGF-βRII, the levels of which are low in epidermal cells and high in dermal cells. 45 We observed an upregulation of tgf-β3 in the regenerating fin, confirming its role in this process. The expression of tgf-βrII regained normal values at 10 dpa, suggesting that similar mechanisms could play a role in zebrafish as well as other vertebrates.
In conclusion, our study showed an implication of bigh3 in fin regeneration. Further detailed analysis is required to elucidate the exact role of bigh3 in this process.
Disclosure statement
No competing financial interests exist.